
栓皮栎和侧柏水分利用率与叶片生理对海拔梯度的协同演化
2022-07-26 06:21温哲华侯沛轩余新晓
山东农业大学学报(自然科学版) 2022年3期
温哲华,侯沛轩,余新晓
栓皮栎和侧柏水分利用率与叶片生理对海拔梯度的协同演化
温哲华1,侯沛轩2,3*,余新晓3
1. 河北省林业和草原局宣传中心, 河北 石家庄 050081 2. 河北省林业和草原工程项目中心, 河北 石家庄 050081 3. 北京林业大学, 北京 100083
本研究选取了华北土石山区四个海拔梯度(50 m、350 m、650 m和950 m)的栓皮栎和侧柏针阔混交林为研究对象,利用稳定碳同位素技术和叶片生理生态观测,分析针阔混交林水分利用效率和叶片生理生态变化适应海拔梯度变化的协同或权衡关系。研究结果表明:海拔每增加100 m,林地土壤含水量和叶片含水量均增加约1.5%,叶比面积和碳同位素比值分别降低约0.06~0.07 cm2/g和0.76~0.81‰,表明针阔林的水分利用效率随海拔梯度逐渐降低。栓皮栎和侧柏的光合速率随海拔呈先上升后降低趋势,并在650 m海拔处达到最大值,分别为4.57 μmol·m-2·s-1,和7.73 μmol·m-2·s-1。栓皮栎和侧柏均能通过调节水分利用效率和改变叶片生理生态协同适应海拔梯度变化,但在海拔梯度下,侧柏的叶片含水量、比叶面积低于栓皮栎,但侧柏的碳同位素比值和光合速率高于栓皮栎,说明栓皮栎对海拔梯度变化的适应更强性。研究结果为我国北方山区森林管护和生态可持续发展和提供理论依据。
栓皮栎; 侧柏;海拔; 协同演化
许多环境参数随海拔变化,包括温度、湿度、降水和太阳辐射[1]。海拔可以通过改变当地的小气候条件(例如地形地貌)以影响区域的降水[2]。此外,不同海拔梯度下水文过程和林分结构变化可能会影响物种间的生理生态过程和物种对环境变化的适应性[3-5]。不少学者担心全球气候变化的海拔效应可能限制植物物种的叶片生理对环境变化的适应性。而这一担心已在其它地区成为现实[6]。樊莹等[7]研究发现随着海拔的上升,蒙古栎幼树的叶片质量和叶片生物量明显减少,说明海拔升高不利于植物光合收益的最大化,这也表明植物会通过改变生物量和生物量的分配使其所受到的资源限制最小化,以此来适应不同的环境。廖兴利等[8]和吴炜等[9]发现植物体内游离脯氨酸含量随着海拔的升高而显著升高,而叶绿素含量等与海拔呈负相关。
水分利用效率(Water Use Efficiency,WUE)是评价植物叶片对环境适应性的一个重要参数[10]。这也是分析叶片水和碳循环关系的重要切入点之一[11]。它已被广泛应用于生态和水文学的研究[12]。随着稳定同位素技术的发展,从植物叶片中提取木质素可以测定稳定碳同位素(δ13C),进而量化植物叶片的短期水分利用效率。当环境发生变化时,植物的正常生理活动将受到干扰,导致水分利用效率异常高或低[13-15]。Henn JJ等[16]发现森林结构的潜在变化会导致植物叶片形态和植物-大气界面碳水关系的变化。有前人研究表明,高海拔地区树木的水分利用效率比低海拔地区对气候变化更敏感[17,18]。低海拔乔木的水分利用效率主要受有效水分的影响,高海拔地区的乔木水分利用效率主要受低温的限制[19]。金高明[20]发现土壤有效水分的增加会降低植物的水分利用效率。然而,由于不同地区的气候和微环境差异,这种模式并不适用于所有地区[21]。此外,一些植物能够调整其叶片形态以适应由海拔变化引起的生态条件的变化。例如,Cui H等[25]在研究中发现,海拔2750 m和3050 m的比叶面积显著低于其他海拔高度。
尽管植物已经表现出了对水分可利用性变化的一些适应,比如从叶片形态、调整光合生理参数和改变水分利用效率适应环境,但在全球气候变化的大背景下,气候变化和海拔梯度的混淆效应较少研究。例如,植物在碳同化和叶片形态间是否存在协同或权衡作用适应环境变化。因此,有必要探究植物叶片生理生态对海拔的适应过程。本文以北京山区分布范围较广的栓皮栎()和侧柏()为研究对象,通过设置不同设置四个海拔梯度(50 m、350 m、650 m和950 m),利用稳定碳同位素技术和叶片生理生态结合,探究植物水分利用效率和植物叶片生理生态过程如何适应海拔变化,辨析以上两者是否存在协同或权衡关系适应海拔变化,研究结果将对我国北方山区生态系统管理和物种恢复提供理论参考。
1 材料与方法
1.1 研究区概况
本研究实验点位于鹫峰国家森林公园的首都圈生态系统定位研究站(40°03´N, 116°05´E)站。研究区年平均气温为11.6 ℃。年平均降水量为660 mm,其中约70%的降水发生在6-9月,年潜在蒸散量为1200 mm~1600 mm。研究区海拔范围为43 m~1153 m。研究区树种组成沿海拔梯度变化,各海拔梯度均为针叶树种和阔叶树种的混合为主,混交树种主要包括栓皮栎、侧柏等,林下有孩儿拳头、荆条等。研究区土壤以粘土为主,表层含有丰富有机质。
1.2 实验设计
在研究区设置了4个海拔梯度,分别为50 m、350 m、650 m和950 m。在每个海拔梯度选择研究区分布范围最广,占比最多的侧柏和栓皮栎针阔混交林设置3个20 m×20 m样地。在每个样地采集土壤和叶片样品,分别测定土壤含水量、叶片含水量、叶片比叶面积、光合生理参数和叶片碳同位素组成等。
1.3 土壤含水量测定
在所选的海拔梯度每一样地中,在所选植株的附近,随机选择地势相对均一地点,用土钻按0-10、10-20、20-40、40-60、60-80、80-100 cm的土壤深度采集样品,每个层次采集3个,立即放入铝盒中,带回室内用烘干称重法测定土壤含水量。
1.4 叶片样品采集与同位素样品分析分析
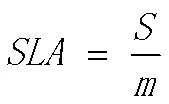
其中,和分别为叶面积(cm2)和叶片干重(g)。
1.5 叶片光合参数测定
在采集叶片样品时,随机选择侧柏和栓皮栎的冠层上方的成熟叶片,利用便携式Li-6400光合仪器测定叶片光合参数,具体操作参考陈根云等。测定时间为06:00-18:00,按照每2小时的间隔测定1次,每次选择3片叶子,重复3次。
1.6 数据处理
利用SPSS 18.0软件统计和分析论文数据。采用双因素(two-way ANOVA)对不同海拔处理的含水量、比叶面积、叶片含水量和水势等进行方差分析,同时利用Duncan法对不同海拔梯度处理多重比较(=0.05)。
2 结果与分析
2.1 不同海拔梯度林地土壤含水量变化规律
50 m、350 m、650 m和950 m4个海拔梯度的林地土壤含水量差异显著,其林地平均土壤含水量分别为17.4%、19.2%、21.8%和31.3%(图1)。950 m的林地土壤含水量比50 m的高79.4%,这表明海拔每升高100 m,林地土壤含水量增加1.5%。4个海拔梯度的林地土壤含水量均随土壤深度呈先升高趋势,并在40 cm~60 cm处达到最大值,100 m、400 m、700、m和1000 m的最大土壤含水量分别为20.4%、22.9%、30.5%和39.5%,随后随着土壤深度的增加而逐渐减小。4个海拔梯度在深层80 cm~100 cm的林地土壤含水量与表层0 cm~10 cm的林地土壤含水量无线组差异。

图 1 林地土壤含水量随海拔的变化规律
2.2 不同海拔梯度植物的叶片含水量和比叶面积变化规律
栓皮栎和侧柏的比叶面积随海拔升高而逐渐降低(2=0.77,<0.05)(图2)。在同一海拔梯度,栓皮栎的比叶面积要显著高于侧柏。栓皮栎在50 m、350 m、650 m和950 m四个海拔梯度的比叶面积差异显著,其平均比叶面积分别为65.6、62.4、62.8和57.9 cm2/g,栓皮栎在50处的比叶面积比其在950处的比叶面积高13.3%。侧柏在50 m、350 m、650 m和950 m四个海拔梯度的比叶面积差异显著,其平均比叶面积分别为63.2、62.7、59.6和56.6 cm2/g,侧柏在50 m处的比叶面积比其在950 m处的比叶面积高11.6%。海拔每升高100 m,栓皮栎和侧柏的比叶面积分别减低0.07 cm2/g和0.06 cm2/g,这表明栓皮栎比叶面积对海拔的响应可能比侧柏敏感。

图 2 栓皮栎和侧柏的比叶面积随海拔的变化规律
2A和2B分别表示栓皮栎和侧柏的比叶面积随海拔的变化规律(不同字母表示同一树种不同海拔间差异显著,<0.05)
The 2a and 2b were the variation of specific leaf area of Q. variabilis and P. orientalis changed with altitude, respectively.
栓皮栎和侧柏的叶片含水量随海拔升高而逐渐增加(2=0.68,<0.05)(图3)。在同一海拔梯度,栓皮栎的叶片含水量要显著高于侧柏,约比侧柏的叶片含水量高9.5%。栓皮栎在50 m、350 m、650 m和950 m四个海拔梯度的叶片含水量差异显著,其平均叶片含水量分别为45.2%、49.6%、51.0%和58.9%,栓皮栎在950 m处的叶片含水量比其在50 m处的叶片含水量高30.3%。侧柏在50 m、350 m、650 m和950 m四个海拔梯度的叶片含水量差异显著,其平均叶片含水量分别为41.2%、45.7%、48.6%和55.8%,侧柏在950 m处的叶片含水量比其在50 m处的叶片含水量高35.5%。海拔每升高100 m,栓皮栎和侧柏的叶片含水量分别升高1.5%和1.6%。

图 3 栓皮栎和侧柏的叶片含水量随海拔的变化规律(不同字母表示同一树种不同海拔间差异显著,P<0.05)
2.3 不同海拔梯度植物光合速率变化规律
栓皮栎和侧柏的日均光合速率随海拔呈先升高,在650 m的海拔处达到最大值,随后降低的趋势。栓皮栎和侧柏从早晨6:00~10:00,光合速率呈上升趋势,在10:00达到峰值,随后呈逐渐降低趋势(图4a)。而栓皮栎到了正午12:00,光合速率出现极小值,而后在下午14:00出现第二次峰值,随后在14:00~18:00,呈下降趋势。在低海拔50 m处,栓皮栎和侧柏光合速率的平均值分别为2.81 μmol·m-2·s-13.72 μmol·m-2·s-1,在10:00的峰值分别为3.94 μmol·m-2·s-1和5.18 μmol·m-2·s-1,随着海拔升高,栓皮栎和侧柏光合速率呈增加趋势,在650 m处的平均光合速率为3.20 μmol·m-2·s-1和4.28 μmol·m-2·s-1,分别增加了27.5%和17.0%;栓皮栎在650 m处的光合速率峰值为4.57 μmol·m-2·s-1,比50 m处的高25.8%。侧柏在650 m光合速率在10:00的峰值为7.73 μmol·m-2·s-1,在14:00的峰值为5.74 μmol·m-2·s-1,14:00峰值比在10:00的峰值小25.7%。随着海拔的继续升高,两者光合速率又呈下降趋势,其950 m处的日均光合速率为3.09 μmol·m-2·s-1和3.40 μmol·m-2·s-1,比650 m处日均光合速率分别低16.4%和12.7%。
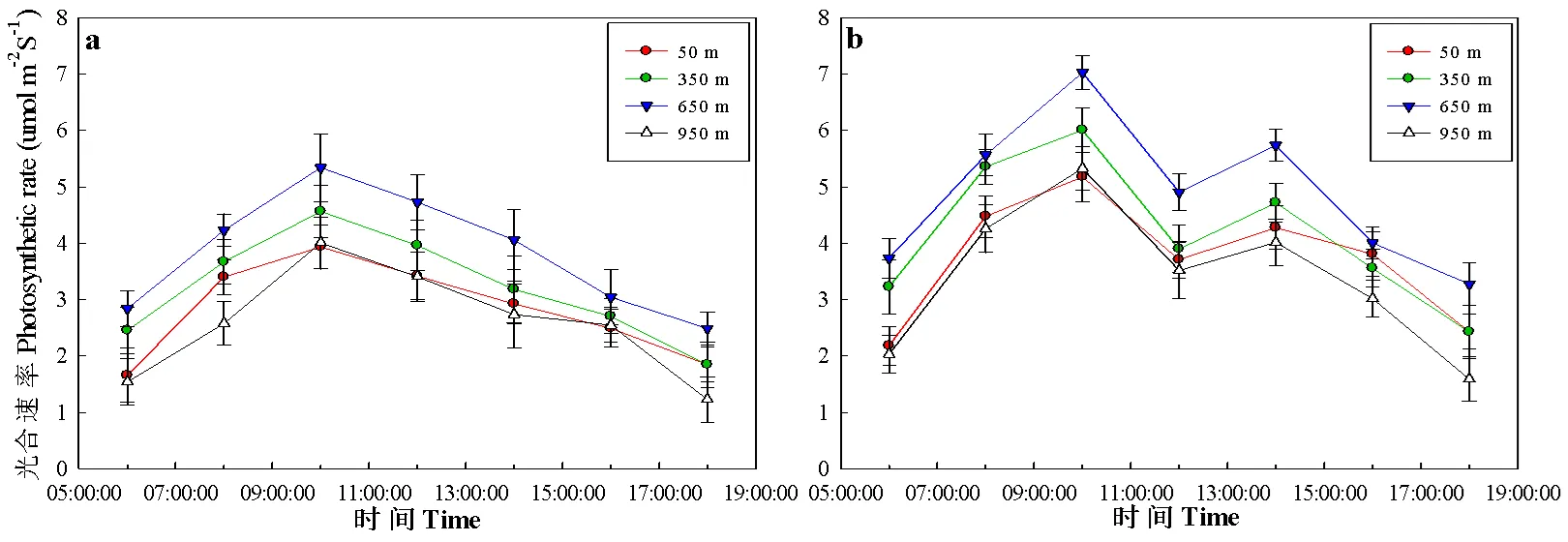
图 4 栓皮栎和侧柏光合速率随海拔的变化规律
4a和4b分别表示栓皮栎和侧柏光合速率随海拔的变化规律
The 4a and 4b were the variation of photosynthetic rate ofandchanged with altitude, respectively
2.4 不同海拔梯度的叶片碳同位素组成
在同一海拔梯度,栓皮栎的碳同位素比值要低于侧柏,约比侧柏的碳同位素比值低9.5%,表明同一海拔梯度范围内侧柏的水分利用效率要高于栓皮栎。栓皮栎在50 m、350 m、650 m和950 m四个海拔梯度的碳同位素比值差异显著,其平均碳同位素比值分别为-22.58‰、-24.81‰、-25.51‰和-29.44‰。栓皮栎在50 m处的碳同位素比值比其在950 m处的碳同位素比值高29.3%。侧柏在50 m、350 m、650 m和950 m4个海拔梯度的碳同位素比值差异显著,其平均碳同位素比值分别为-20.61‰、-22.88‰、-24.29‰和-27.93‰,侧柏在50 m处的碳同位素比值比其在950 m处的碳同位素比值高33.3%。海拔每升高100 m,栓皮栎和侧柏的碳同位素比值分别降低了0.76‰和0.81‰。

图 5 栓皮栎和侧柏叶片碳同位素组成随海拔的变化规律(不同字母表示同一树种不同海拔间差异显著,P<0.05)
3 讨 论
土壤水分主要受降水补给和蒸散作用处于不断的波动状态。本研究的土壤含水量随海拔梯度的升高而增大,可能是由于海拔越高,林冠截留量越少,林内降雨越多,导致土壤水分增加。较为充足的土壤水分被植物吸收后运输到叶片,导致叶片的含水量与土壤水分呈现同步的变化规律。此外,林地微气象随着海拔梯度发生变化,如温度降低、辐射和土壤含水量增加,这可能会影响植物叶片气孔开闭,降低植物的光合能力,进而降低植物的水分利用效率[26-28]。本研究也发现,栓皮栎和侧柏的光合速率与水分利用效率均随着海拔升高而逐渐降低,这可能栓皮栎和侧柏对海拔梯度的一种适应策略。郭文文等[29]也表明植物一般可以可能通过减少叶宽、叶长和调节水分利用效率从形态和生理上对海拔的适应。随着海拔的升高,栓皮栎和侧柏的叶比面积逐渐减少,这可能是栓皮栎和侧柏对高海拔强辐射和低温的一种适应策略。高海拔地区的光和辐射高于低海拔地区,植物通过减少叶面积减少蒸腾损失[30],与此同时,有些在低海拔地区植物增加叶量和叶面积以应对较低的光强和辐射[31]。Midolo G等[32]表明当光照不足时,植物可以通过增加叶比面积截获更多的光能,而当光照强度增加时,植物会降低比叶面积。Poorter H等[33]也表明,在土壤水分含量较低的环境中,降低比叶面积可减少植物蒸腾作用。在我们的研究区,高海拔950 m处的林地土壤质量相对较差,而高海拔区林地土壤含水量高,比叶面积较低。这与前人研究结果稍有不同,李芳兰等[34]发现有较低比叶面积的植物比高比叶面积的植物更能适应干旱和贫瘠的土壤环境。这可能是由于土壤养分和土壤水分的双重作用,而土壤养分对栓皮栎和侧柏的影响要大于土壤水分,导致高海拔地区的比叶面积较低。本研究表明比叶面积与植物的光合速率呈现的正相关,这与白婷等[35]研究结果相似。施建敏等[36]发现,为了缓解强光对植物叶片组织的损伤,高海拔植物会减少对光合系统的氮分配,而增加对保护性结构成分的分配,导致光合固碳量和水分利用效率下降。这也证实了本文提出的观点。本研究中栓皮栎和侧柏的水分利用效率与光合速率随海拔呈先上升后降低的趋势,在海拔650 m处达到峰值。这有可能是因为在海拔650 m处恰好能让栓皮栎和侧柏达到大的光合固碳能力,而950 m处的辐射太大,栓皮栎和侧柏为了减少对叶片组织伤害,启动了对植物的保护性机制。尽管栓皮栎和侧柏处于同一土壤水分环境,但两者对环境的适应呈现差异。在同一海拔梯度中,侧柏的水分利用效率和光合速率均高于栓皮栎,但侧柏的叶片含水量低于栓皮栎,这表明栓皮栎叶片中的水分多数通过气孔返回大气,而侧柏保留的水分较多,这表明侧柏对海拔的适应性优于栓皮栎。在季节性干旱的华北土石山区,植物可能长期处于水分匮缺的状态,侧柏这种对环境的适应性可能有利于其渡过干旱的胁迫环境。Liu Z等[37]研究也发现侧柏对水分的利用深度和范围要大于栓皮栎。在研究区,我们发现侧柏分布的范围和占比栓皮栎大,这也可能是侧柏对环境适应性要强与栓皮栎的原因。
4 结 论
林地土壤含水量和叶片含水量与海拔梯度呈正相关关系,而混交林的叶比面积和碳同位素比值与海拔呈负相关关系,表明针阔林的水分利用效率随海拔梯度逐渐降低。海拔每增加100 m,林地土壤含水量和叶片含水量均增加约1.5%,叶比面积和碳同位素比值分别降低约0.06~0.07 cm2/g和0.76~0.81‰。栓皮栎和侧柏的光合速率随海拔呈上升趋势,并在650 m海拔处达到最大值,分别为4.57 μmol·m-2·s-1,和7.73 μmol·m-2·s-1,随后呈降低趋势。栓皮栎和侧柏均能通过调节水分利用效率和改变叶片生理生态协同适应海拔梯度变化,但在海拔梯度下,侧柏的叶片含水量、比叶面积低于栓皮栎,但侧柏的碳同位素比值和光合速率高于栓皮栎,表现出比栓皮栎对海拔梯度更强的适应性。研究结果为我国北方山区生态系统管理和物种恢复提供理论参考。
[1] Midolo G, De Frenne P, Hölzel N,Global patterns of intraspecific leaf trait responses to elevation [J]. Global Change Biology, 2019,25(7):2485-2498
[2] Ryel RJ, Caldwell MM, Leffler AJ. Rapid soil moisture recharge to depth by roots in a stand of Artemisia tridentate [J]. Ecology, 2003,84(3):757-764
[3[ Szymczak S, Barth J, Bendix J,. First indications of seasonal and spatial variations of water sources in pine trees along an elevation gradient in a Mediterranean ecosystem derived from δ18O [J]. Chemical Geology, 2020,549:119695
[4] 马元屾,王中生,余华,等.不同海拔梯度下黄山松与马尾松针叶形态·光合生理特性的研究[J].安徽农业科学,2012,40(29):14155-14158,14173
[5] 柴勇,朱华,孟广涛,等.云南哀牢山古茶树群落优势树种的种群结构与分布格局[J].林业科学研究,2011,24(3):277-284
[6] Mills SL, Eugenia V, Bragina AV,. Winter color polymorphisms identify global hot spots for evolutionary rescue from climate change [J]. Science, 2018,359(6379):1033-1036
[7] 樊莹,乔雪涛,赵秀海.长白山自然保护区蒙古栎幼树生理生长特性随海拔梯度的变化[J].北京林业大学学报,2019,41(11):1-10
[8] 廖兴利,黎云祥,权秋梅,等.连香树生理生化物质随海拔的变化及其适应性研究[J].广西植物,2011,31(5):641-645
[9] 吴炜,高小明,胡娟娟,等.北缘油茶对不同海拔生境的生理响应[J].安徽农业大学学报,2019,46(4):655-659
[10] Philips DL, Newsome SD, Gregg JW. Combining sources in stable isotope mixing models: alternative methods [J]. Oecologia, 2005,144:520-527
[11] Keenan TF, Hollinger DY, Bohrer G. Increase in forest water–use efficiency as atmospheric carbon dioxide concentrations rise [J]. Nature, 2013499(7458):324-327
[12] Tarin T, Nolan RH, Medlyn BE,. Water–use efficiency in a semi–arid woodland with high rainfall variability [J]. Global Change Biology, 2019,11:1354-1013
[13] 朱军涛,李向义,张希明,等.昆仑山北坡前山带塔里木沙拐枣对不同海拔生境的生理生态响应[J].生态学报,2008,30(3):602-609
[14] 周永斌,吴栋栋,于大炮,等.长白山不同海拔岳桦非结构碳水化合物含量的变化[J].植物生态学报,2009,33(1):118-124
[15] 潘红丽,李迈和,蔡小虎,等.海拔梯度上的植物生长与生理生态特性[J].生态环境学报,2009,18(2):722-730
[16] Henn JJ, Buzzard V, Enquist BJ,. Intraspecific trait variation and phenotypic plasticity mediate alpine plant species response to climate change [J]. Frontiers in Plant Science, 2018,9:1548
[17] Körner C, Farquhar GD, Roksandic Z. A global survey of carbon isotope discrimination in plants from high altitude [J]. Oecologia,198874(4):623-632
[18] 杜流姗,陆琦,梁紫嫣,等.贡嘎山海螺沟冰川退缩区原生演替不同阶段优势植物光合生理特征[J].生态环境学报,2019,28(12):2356-2363
[19] Wright IJ, Dong N, Maire V,. Global climatic drivers of leaf size [J]. Science, 2017,357(6354):917-921
[20] 金高明.黄土高原不同海拔高度的富士苹果光合特性及生理效应的比较研究[D].兰州:甘肃农业大学,2015
[21] Benítez LA, Alkemade R, Schipper AM,. The impact of hunting on tropical mammal and bird populations [J]. Science, 2017,356(6334):180-183
[22] 黄晓霞,冯程程,姜永雷,等.云南玉龙雪山川滇高山栎生理生化特性对海拔梯度的响应[J].西部林业科学,2015,44(4):1-6,24
[23] Sun XY, Wang GX, Huang M,. The asynchronous response of carbon gain and water loss generate spatio-temporal pattern of WUE along elevation gradient in southwest China [J]. Journal of Hydrology, 2019,581(2):124389
[24] 刘贤赵,宿庆,李嘉竹,等.控温条件下C3、C4草本植物碳同位素组成对温度的响应[J].生态学报,2015,35(10):3278-3287
[25] Cui H, Töpper JP, Yang Y,. Plastic population effects and conservative leaf traits in a reciprocal transplant experiment simulating climate warming in the Himalayas [J]. Frontiers in Plant Science, 2018,9:1069
[26] 付作琴,吕茂奎,李晓杰,等.武夷山不同海拔黄山松新叶和老叶氮磷化学计量特征[J].生态学杂志,2019,38(3):648-654
[27] 魏捷,余辉,匡廷云,等.青海高原不同海拔珠芽蓼叶绿体超微结构的比较[J].植物生态学报,2000,24(3):304-307
[28] 王玉萍,高会会,刘悦善,等.高山植物光合机构耐受胁迫的适应机制[J].应用生态学报,2013,24(7):2049-2055
[29] 郭文文,卓么草,何竹,等.藏东南色季拉山不同海拔川滇高山栎叶解剖结构及环境适应性[J].西南林业大学学报 (自然科学),2022,42(6):1-6
[30] Xu HY, Wang H, Prentice IC. Predictability of leaf traits with climate and elevation: a case study in Gongga Mountain, China [J]. Tree Physiology, 2021,41(8):tpab003
[31] García MP, Cásares GM, Gazol A. Run to the hills: Forest growth responsiveness to drought increased at higher elevation during the late 20th century [J]. Science of the Total Environment, 2021,772:145286
[32] Midolo G, Alkemade R, Schipper A,Impacts of nitrogen addition on plant species richness and abundance: A global meta–analysis [J]. Global Ecology and Biogeography, 2019,28:398-413
[33] Poorter H, Niinemets U, Poorter L,. Causes and consequences of variation in leaf mass per area (LMA) [J]. The New Phytologist, 2009,182(3):565-588
[34] 李芳兰,包维楷,刘俊华,等.岷江上游干旱河谷海拔梯度上白刺花叶片生态解剖特征研究[J].应用生态学报,2006,15(1):5-10
[35] 白婷,靳玉龙,朱明霞,等.海拔差异对青稞品质影响的研究[J].中国粮油学报,2019,34(2):34-39
[36] 施建敏,叶学华,陈伏生,等.竹类植物对异质生境的适应——表型可塑性[J].生态学报,2014,34(20):5687-5695
[37] Liu Z, Yu X, Jia G. Water uptake by coniferous and broad leaved forest in a rocky mountainous area of northern China [J]. Agricultural and Forest Meteorology, 2019,265(1):381-389
Synergistic Evolution of Water Availability and Leaf Physiology with Altitude Gradients inand
WEN Zhe-hua1, HOU Pei-xuan1,2*, YU Xin-xiao2
1.050081,2.100083,
The coniferous and broad-leaved mixed forests ofandat four altitudes (50 m, 350 m, 650 m and 950 m) in the rocky mountain area of North China were selected as the research objects. The stable carbon isotope technique and leaf physiological and ecological observation were used to analyze the synergistic or trade-off relationship between water use efficiency and leaf physiological and ecological changes of coniferous and broad-leaved mixed forests. The results showed that soil water content and leaf water content were positively correlated with altitude gradient, while leaf specific area and carbon isotope ratio of mixed forest were negatively correlated with altitude, indicating that water use efficiency of coniferous and broad-leaved forest decreased with altitude gradient. When the altitude increased by 100 m, the soil water content and leaf water content increased by 1.5%, and the leaf specific area and carbon isotope ratio decreased by 0.06-0.07 cm2/g and 0.76-0.81‰ respectively. The photosynthetic rate ofandwere 4.57 μmol·m-2·S-1and 7.73 μmol·m-2·s-1, respectively at 650 m. Bothandcould adapt to the change of altitude gradient by adjusting water use efficiency and changing leaf physiology and ecology. However, under the altitude gradient, the leaf water content and specific leaf area ofwere lower than that of, but the carbon isotope ratio and photosynthetic rate were higher than that of, showing stronger adaptability to the altitude gradient than that of. The results provide a theoretical reference for ecosystem management and species restoration in northern mountain areas of China.
;altitude; synergistic evolution
Q941
A
1000-2324(2022)03-0433-07
10.3969/j.issn.1000-2324.2022.03.015
2021-08-25
2021-10-10
国家自然科学基金项目(42007182);国家重点研发计划项目(2020YFF0305905-03)
温哲华(1976-),男,大学本科,工程师,研究方向:生态学. E-mail:zhehuawen0608@163.com
Author for correspondence. E-mail:459240446@qq.com
猜你喜欢
农学学报(2022年8期)2022-11-16
新农民(2022年22期)2022-11-07
农业与技术(2022年16期)2022-09-01
热带农业工程(2021年5期)2021-12-06
农村实用技术(2021年11期)2021-12-03
星星·诗歌原创(2021年3期)2021-06-01
扬子江(2018年1期)2018-01-26
分析化学(2017年5期)2017-06-21
同位素(2014年3期)2014-06-13
同位素(2014年2期)2014-04-16
