
牦牛METTL14 CDS区克隆及表达谱分析
2022-07-23 07:50马兰花顾亚荣张永峰潘和平西北民族大学生命科学与工程学院甘肃兰州730030中国农业科学院兰州畜牧与兽药研究所甘肃省牦牛繁育工程重点实验室甘肃兰州730030
中国兽医学报 2022年5期
马兰花,顾亚荣,张永峰,潘和平,阎 萍 (1.西北民族大学 生命科学与工程学院,甘肃 兰州 730030;.中国农业科学院 兰州畜牧与兽药研究所 甘肃省牦牛繁育工程重点实验室,甘肃 兰州 730030)
牦牛生活在海拔3 000 m以上的高寒地区,作为高原上宝贵的生物遗传资源和财富,这是高原藏区的文化符号之一,具有极高的研究和开发价值[1]。牦牛在食物来源及种类匮乏、自然气候条件恶劣的高海拔高原生态环境中表现出极强的适应性,利用牦牛这种生物特性开展牦牛遗传资源保护与开发利用,对促进藏区经济价值和生态价值具有十分重要的发展意义[2]。青海省大通牦牛是通过传统的杂交手段培育的首个肉用型牦牛新品种[3],拥有肉质鲜美、高蛋白、低脂肪等特点[4-5],在长期自然选择过程中,其脂肪发育脂质沉积的调控在品种和组织中具有明显的特异性[6],但牦牛生长缓慢,脂肪沉积效率低、嫩度差、口感粗糙,影响牦牛的肉用价值[7]。随着生物技术的发展,从分子水平研究生物效应中的基础作用关系,有利于运用遗传学手段来改善生物表观遗传水平,使生物表型朝着生态/经济双重价值的方向发展[8]。
N6-腺苷酸甲基(N6-methyladenosine,m6A)是RNA分子上特定腺嘌呤N6位置上的CH3基团,是真核生物中最常见的一种动态可逆的转录后修饰。甲基化酶样蛋白14(methyltransferase-like protein 14,METTL14)作为m6A甲基化过程中底物结合平台,与甲基化酶样蛋白3(methyltransferase-like protein 3,METTL3)形成稳定的复合物[9],激活并增强METTL3的催化活性。复合物被肾母细胞瘤1相关蛋白(Wilms tumor 1 associated protein,WTAP)间接定位在核斑点上,S-腺苷甲硫氨酸(S-adenosyl methionine,SAM)作为甲基供体,在METTL3和METTL14甲基转移酶的催化下,将甲基转移到腺苷(A)碱基第6位的氮上,形成N6-甲基腺苷(m6A)。另外,METTL16、KIA也是甲基转移酶的组分之一,协同作用于甲基化过程[10-11]。m6A结合蛋白可以识别甲基化修饰,影响mRNA的命运。m6A结合蛋白主要包括YTH结构域蛋白家族YTHDF1-3、YTHDC1-2等,还有部分hnRNA蛋白家族,比如hnRNA2B1等[12]。与甲基化酶具有逆向作用的去甲基化酶,肥胖相关蛋白(fat mass and obesity associated protein,FTO)和ALKBH同源蛋白5(Alk B homologue 5,ALKBH5),能对已发生m6A修饰的碱基进行去甲基化修饰[13]。甲基化和去甲基化是由各自酶作用的可逆化学过程,用于动态平衡m6A的活性。
METTL14是m6A 的核心酶之一,与METTL3形成异质二聚体[14]。有研究发现,METTL14可能在多种细胞分化发育过程中起到决定细胞命运的作用。沉默处理METTL3和 METTL14可抑制小鼠胚胎干细胞的自我更新[15]。在晚期精原细胞分裂过程中,METTL3和METTL14 2个基因联合缺失会破坏精子发生[16]。然而,点突变METTL14 和METTL3活性中心的关键氨基酸,发现METTL3突变后会导致异质二聚体的活性降低,而METTL14活性并没有明显的改变,说明METTL14是异质二聚体的非催化亚基,其在甲基化过程中具有特异性功能。在小鼠睾丸间质细胞分化过程中,METTL14表达量逐渐降低,表明m6A修饰与睾丸激素合成过程中睾丸间质细胞的自噬之间存在负相关,在睾丸间质细胞启始阶段METTL14表达量最高[17]。缺少METTL14的小鼠Treg细胞(功能调节性T细胞)的转录因子RORγt表达降低,表明METTL14缺乏会导致Treg细胞的诱导分化受损[18]。敲除小鼠胚胎大脑皮质神经干细胞中的METTL14,皮质神经发生会迟缓到出生后阶段,胶质细胞的细胞周期也会延长[19]。m6A修饰在哺乳动物前体脂肪细胞分化中发挥着重要的调控作用,WTAP/METTL3/METTL14复合物通过促进细胞有丝分裂周期转变积极调控脂肪发生,敲除这3个基因会使细胞周期蛋白A2上调,导致细胞周期停止,影响脂肪细胞分化[20]。对猪脂肪细胞早期增殖分化与脂质代谢能力的研究发现,过量的支链氨基酸(branched-chain amino acids,BCAA)能使猪前体脂肪细胞中METTL14下调,从而通过m6A RNA甲基化修饰的方式抑制脂质代谢的过程[21]。热应激时脂肪细胞大量从血液中摄取甘油三酯并沉积在脂肪组织中,此时m6A甲基化水平提高,甲基化的作用大于去甲基化的作用,METTL14在脂肪组织中的表达量显著提高[22-23]。所有这些结果均表明,METTL14在细胞分化、脂肪沉积等过程中有着重要作用。目前,m6A修饰在脂肪沉积中的相关研究主要集中在去甲基化酶FTO的研究上[24],而METTL14与反刍动物脂肪沉积之间的研究尚不清楚。本试验通过对大通牦牛METTL14进行克隆及序列分析,依据分子生物信息学技术,在分子和细胞2个层面,分析牦牛脂肪组织中METTL14的生物信息学特性及表达规律,以期为阐释METTL14调控高原家畜脂肪沉积的特异性分子机制奠定理论基础。
1 材料与方法
1.1 组织样品选取体质量相近的18月龄和30月龄的大通牦牛各3头,屠宰后采集其心脏、肝脏、脾脏、肺脏、肾脏、背最长肌、皮下脂肪组织,用PBS洗涤,液氮过夜,-80℃保存备用。
1.2 细胞体外培养牦牛前体脂肪细胞由实验室前期培养并提供[25],细胞贴壁生长至培养孔底面积的70%左右时,用胰岛素、3-异丁基-1-甲基黄嘌呤和地塞米松处理脂肪细胞2 d,诱导其分化,此后,每隔1 d用含10%胎牛血清和10 mg/L胰岛素的培养液替换正常培养液,持续培养12 d[26]。
1.3 主要试剂PrimeSTAR HS DNA Polymerase、PrimeScriptTMRT reagent Kit、pMD19-T Vector Cloning Kit、TRIzol、Transcriptor First Strand cDNA Synthesis Kit购自宝生物工程(大连)有限公司;SYBR Premix Ex TaqTMⅡ、E.ColiDH5α感受态细胞、TIANgel Midi Purification Kit购自北京全式金生物技术公司;氨苄西林、三氯甲烷购自Sigma公司。
1.4 主要仪器低温离心机购自苏州培英实验设备有限公司;梯度PCR仪购自美国伯乐公司;压力蒸汽灭菌器购自上海申安医疗器械厂;Bio-Rad CFX 96 qRT-PCR购自时代联想生物科技有限公司;生物安全柜购自上海力申科学仪器有限公司。
1.5 总RNA的提取及反转录取-80℃保存的组织,为提高提取的RNA质量,脂肪组织提取前去除油脂[27],加入液氮研磨后用TRIzol试剂、三氯甲烷提取RNA;牦牛前体脂肪细胞用六孔板培养至12 d,分别收集培养时间为0,4,8,12 d的细胞并用TRIzol试剂、三氯甲烷提取细胞总RNA。用紫外分光光度计测定RNA含量及纯度(D260 nm/D280 nm>1.8),通过1.2%琼脂糖凝胶电泳鉴定RNA完整性。用PrimeScrip RT reagent Kit将总RNA 反转录合成长片度cDNA,于-20℃保存备用。
1.6 牦牛METTL14引物设计与合成根据NCBI GenBank数据库中公布的野牦牛METTL14(XM_005898072.2)序列,确定METTL14的CDS区,采用Primer-BLAST设计METTL14的PCR扩增引物、qRT-PCR引物以及qRT-PCR内参基因β-actin引物,引物相关信息见表1。

表1 METTL14扩增及qRT-PCR所用引物信息
1.7 牦牛METTL14的克隆与测序根据TaKaRa公司PrimeScriptTMRT reagent Kit的说明书操作,将牦牛脂肪组织RNA反转录成长片段cDNA,以此为模板,用METTL14上游引物F、下游引物R扩增牦牛METTL14的CDS区全长序列。反应体系25 μL:2×Taq PCR Master Mix 12.5 μL,METTL14-F/R各1 μL,cDNA 2 μL,ddH2O 8.5 μL。反应条件:95℃预变性3 min;95℃变性30 s,55.1℃退火30 s,72℃延伸2 min,37个循环;72℃延伸10 min。PCR扩增产物经1%琼脂糖凝胶电泳检测后,切胶回收目的片段并连接至pMD19-T载体,转化至Trans5α感受态细胞中,涂布到含氨苄西林的LB固体培养基上,37℃倒置培养12 h,挑取单菌落继续振荡培养12 h后,进行菌液PCR鉴定,将阳性菌液送西安擎科生物技术有限公司进行测序。
1.8 牦牛METTL14的生物信息学分析将METTL14测序结果用SeqMan拼接后,用表2中生物学软件进行基因序列和蛋白质结构分析。

表2 核苷酸和蛋白质序列数据分析工具
1.9 牦牛METTL14 qRT-PCR检测选用β-actin(NM_001009784)作为内参基因,Primer-BLAST设计qRT-PCR引物(表1)。使用Bio-Rad CFX 96 qRT-PCR仪检测目的基因及内参基因的表达量,每组设生物学重复和技术重复(3×3)。反应体系10 μL:SYBR Premix Ex TaqTMⅡ5 μL,METTL14-F/R(10 μmol/L)各0.4 μL,cDNA模板0.8 μL,ddH2O 3.4 μL。反应条件:95℃预变性1 min;95℃变性30 s,61.4℃退火30 s,95℃延伸30 s,45个循环;55℃延伸2 s。观察荧光信号的变化,信号强度随着温度的升高先上升后降低,且形成单一峰,说明产物特异性良好。
2.0 统计分析qRT-PCR结果采用2-△△Ct方法进行分析,数据均用平均值±标准误表示,用SPSS 23.0软件进行比较平均值中的独立样本t检验。
2 结果
2.1 牦牛METTL14编码基因扩增及克隆根据野牦牛METTL14 mRNA设计特异性引物,以大通牦牛脂肪组织cDNA 为模版进行PCR扩增,扩增产物经凝胶电泳后,通过紫外凝胶成像仪观察到在1 666 bp 附近出现清晰条带(图1),特异性良好,与预期结果相符。
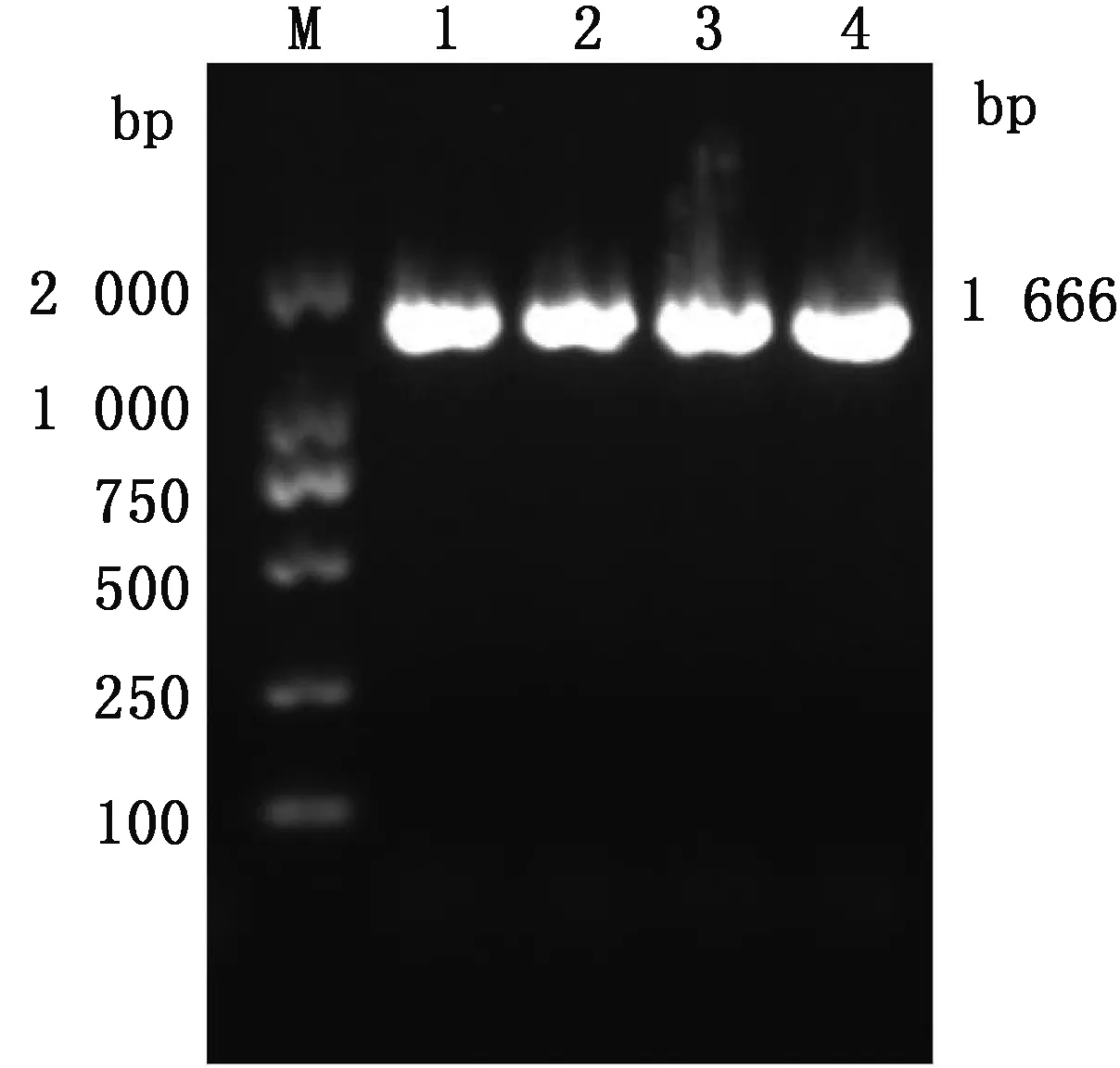
M.DL2000 DNA Marker;1~4.METTL14扩增产物图1 牦牛METTL14的PCR扩增产物琼脂糖凝胶电泳结果
2.2 牦牛METTL14 mRNA CDS区序列分析目的基因测序结果经SeqMan软件拼接,获得METTL14 cDNA序列的长度为1 666 bp,与预期一致。使用NCBI的ORF Finder进行分析,获得1个1 371 bp 的最长ORF,与GenBank中公布的野牦牛(XM_005898072.2)METTL14 mRNA CDS的同源性为99.81%,因此,确定大通牦牛METTL14 CDS区长度为1 371 bp,编码456个氨基酸。
2.3 牦牛METTL14基因及其编码蛋白同源性比对分析和系统进化树构建
2.3.1METTL14基因及其编码蛋白序列同源性比对 将牦牛METTL14的核苷酸序列与GenBank中与野牦牛(Bosmutus,XM_005898072.2)、黄牛(Bostaurus,NM_001083714.1)、绵羊(Ovisaries,XM_004009592.4)、猪(Susscrofa,XM_003129231.6)、双峰骆驼(Camelusbactrianus,XM_010952608.1)、人(Homosapiens,NM_020961.4)、野马(Equusprzewalskii,XM_008534527.1)、猕猴(Macacanemestrina,XM_011748914.2)、兔(Musmusculus,NM_201638.2)、小鼠(Musmusculus,NM_201638.2)、鸡(Gallusgallus,NM_001031148.1)等11个物种该基因CDS区序列进行同源性比对。结果发现,大通牦牛METTL14核苷酸序列与野牦牛、黄牛、绵羊、猪、双峰骆驼、人、野马、猕猴、兔、小鼠和鸡苷酸序列的同源性分别为99.8%,99.6%,99.0%,95.5%,95.1%,95.1%,94.8%,94.5%,92.0%,89.9%和81.5%(图2);大通牦牛METTL14氨基酸序列与GenBank中的野牦牛(Bosmutus,XP_005898134.1)、绵羊(Ovisaries,XP_004009641.1)、野马(Equusprzewalskii,XP_008532749.1)、黄牛(Bostaurus,NP_001077183.1)、猪(Susscrofa,XP_003129279.3)、猕猴(Macacanemestrina,XP_011747216.1)、人(Homosapiens,NP_066012.1)、双峰骆驼(Camelusbactrianu,XP_010950910.1)、兔(Oryctolaguscuniculus,P_002717295.2)、小鼠(Musmusculus,XP_004009641.1)、鸡(Gallusgallus,NP_001026319.1)等11个物种的氨基酸序列同源性分别为99.6%,99.6%,99.6%,99.3%,99.1%,99.1%,99.1%,98.9%,98.9%,98.3%,93.5%(图3)。
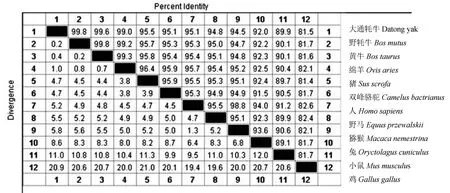
图2 大通牦牛与其他物种METTL14核苷酸序列同源性对比结果
图3 大通牦牛与其他物种 METTL14蛋白氨基酸序列同源性比对结果
2.3.2METTL14核苷酸序列系统进化树分析 通过MEGA 7.0软件构建核苷酸同源性系统进化树,分析了包括大通牦牛METTL14 CDS区在内的11个物种该基因的CDS区核苷酸序列。建树结果(图4)表明,在11个物种中,大通牦牛METTL14和野牦牛的亲缘关系最近,与鸡和小鼠的亲缘关系最远。
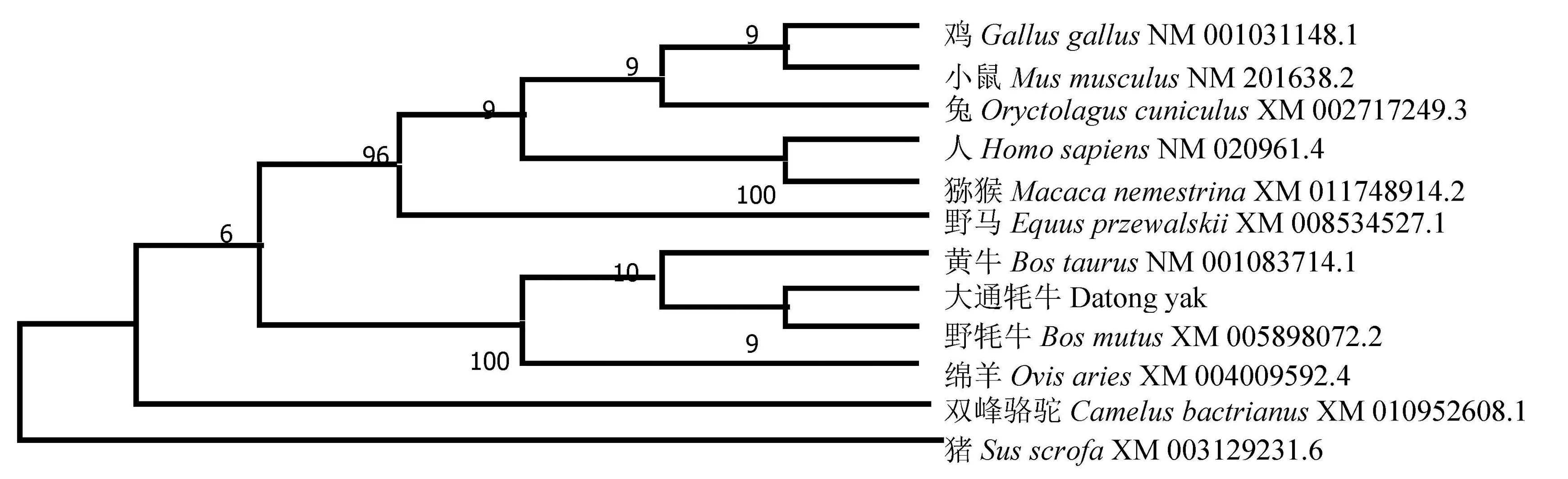
图4 牦牛METTL14系统进化树
2.4 牦牛METTL14蛋白质分析
2.4.1蛋白及核酸理化性质分析 经BioEdit软件分析发现,METTL14碱基分布情况为:A、G、C和T平均含量分别为32.60%,18.31%,24.95%和24.14%,其中G+C的平均含量(43.25%)低于A+T平均含量(56.75%),表明该基因碱基使用存在偏向性,偏好使用AT碱基。经ProtParam软件分析发现,大通牦牛METTL14分子式为C2277H3575N669O709S16,其相对分子质量为52 193.33 kDa,等电点为5.89,氨基酸组分中谷氨酸含量(10.50%)最高,色氨酸含量(1.30%)和蛋氨酸含量(1.30%)最少(表3)。METTL14分子中正电荷氨基酸残基 (Arg+Lys) 70个,负电荷氨基酸残基(Asp+Glu) 77个。METTL14的脂肪系数(AI)为66.14,不稳定系数(Ⅱ)为52.76,总平均亲水指数(GRAVY)为-0.878。
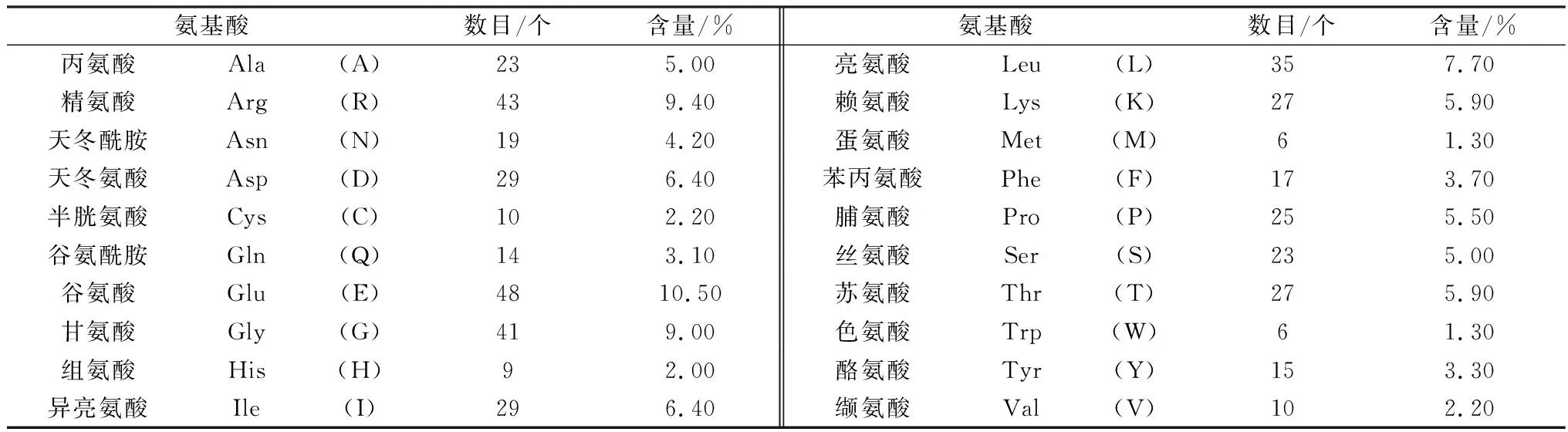
表3 牦牛METTL14蛋白的氨基酸组分
2.4.2METTL14亲水性/疏水性预测 经ExPASyProtScale软件预测发现,METTL14亲水性氨基酸的总分值高于疏水性氨基酸(图5),牦牛METTL14总体表现出强的亲水性,与GRAVY值预测相符。多肽链第429位精氨酸(R)亲水性最强(-3.300);第234位色氨酸(W)疏水性最强(1.544)。
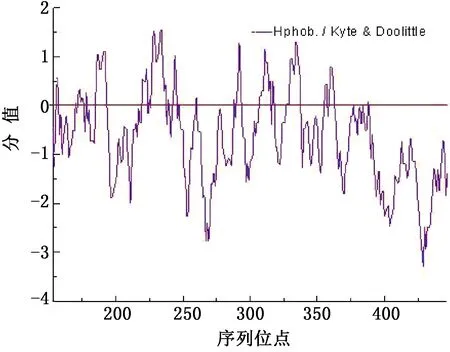
图5 牦牛METTL14蛋白的亲疏水性分析
2.4.3METTL14结构域预测 通过CDD软件对METTL14的保守结构域预测显示,在METTL14氨基酸序列的186~363位有1个MT-A70结构域(图6),E-value=8.22e-80。MT-A70是mRNA上SAM的结合亚基,可特异地甲基化前体pre-mRNA中的腺嘌呤。
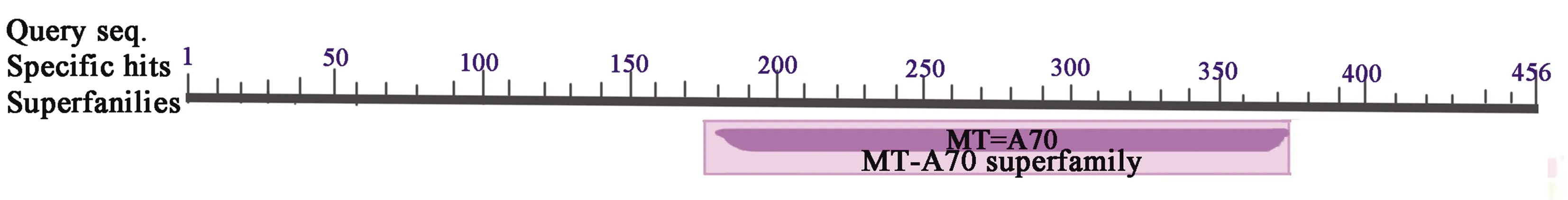
图6 牦牛METTL14蛋白保守结构域
2.4.4METTL14信号肽和跨膜结构域预测运用 SignalP 4.1分析METTL14氨基酸序列,预测有信号肽的概率是0.092%,C值为0.111,Y值为0.119,S值为0.161,S-mean及D-mean值分别为0.127、0.132,均小于阈值0.450,说明METTL14 无信号肽(图7)。通过TMHMM METTL14跨膜结构域进行预测(图8),显示METTL14均处于膜内,即METTL14无跨膜结构域,不是膜蛋白。
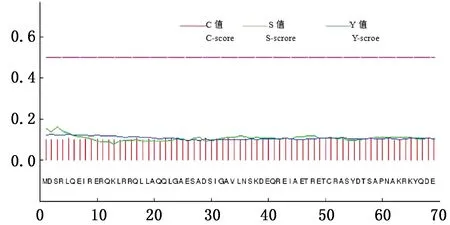
图7 牦牛METTL14蛋白信号肽预测
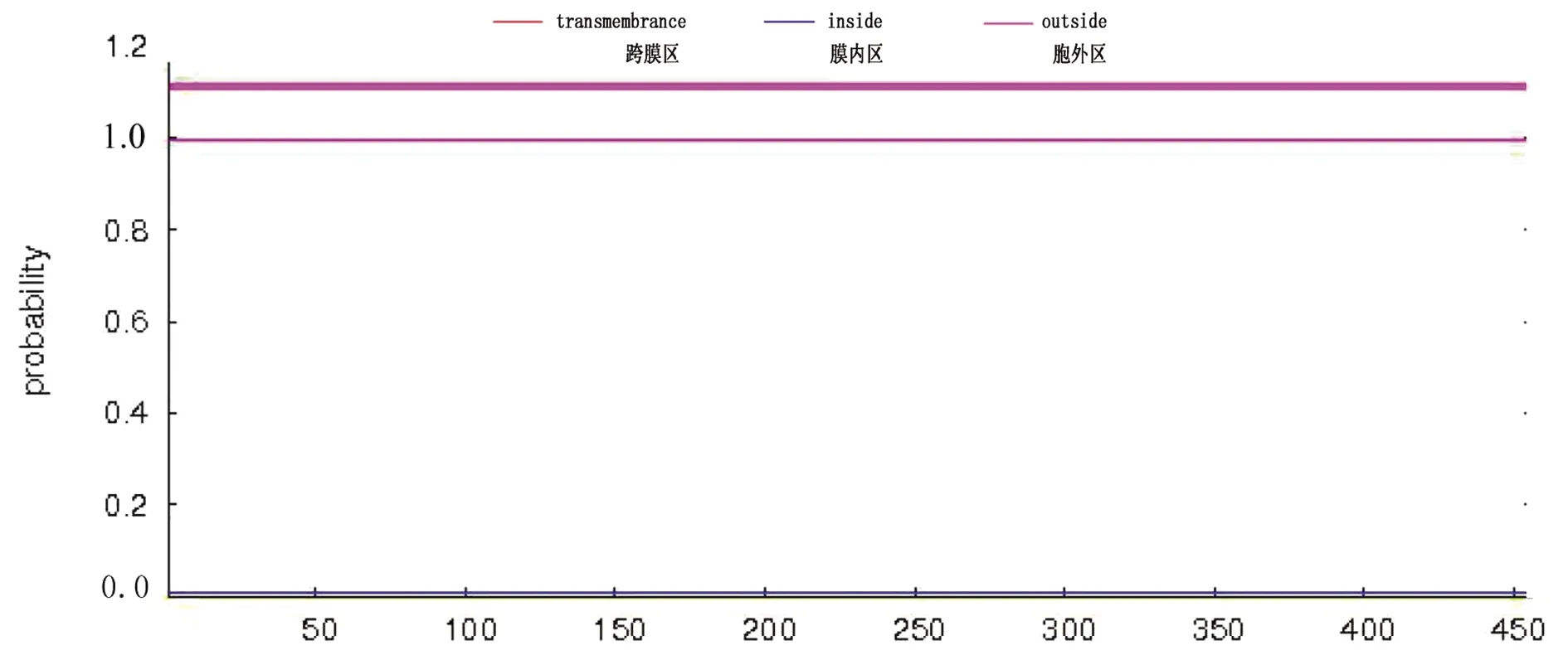
图8 牦牛METTL14蛋白跨膜结构域预测
2.4.5METTL14亚细胞定位与核定位预测 用PSORT预测METTL14亚细胞定位,k-NN预测结果(表4)显示METTL14定位在细胞核上(69.60%)。用cNLS-mapper对METTL14进行分析,发现其存在一段核定位序列,序列为PNAKRKYQDE (score=8),位置在59~70位之间。

表4 牦牛METTL14亚细胞定位预测
2.4.6METTL14潜在磷酸化位点预测 使用NetPhos 3.1 Server分析METTL14的氨基酸序列潜在磷酸化位点,结果显示,该序列具有40个潜在的磷酸化位点,丝氨酸(serine)、苏氨酸(threonine)及酪氨酸(tyrosine)占比分别为35%,42.5%和22.5%(图9)。

图9 牦牛METTL14基因氨基酸序列潜在磷酸化位点分析
2.4.7METTL14二级结构预测 利用Sopma软件预测牦牛METTL14二级结构(图10),发现牦牛METTL14中的无规则卷曲占51.10%、α-螺旋占33.11%,氨基酸残基连接成的延伸链占11.84%,β-转角占3.95%。其中,柔性无规则卷曲占比较高。
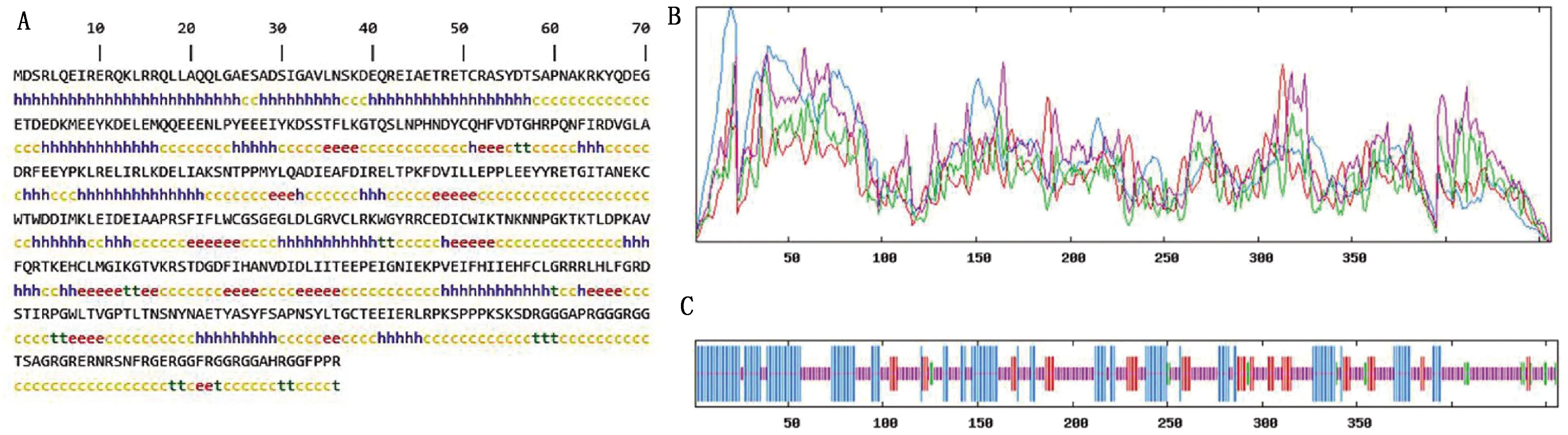
A.蛋白质二级结构位点图(h.α-螺旋;c.无规则卷曲;t.β-转角;e.延伸);B.蛋白质二级结构峰图;C.蛋白质二级结构竖条纹图。紫色.示无规则卷曲;蓝色.示α-螺旋;红色.示延伸链;绿色.示β-转角图10 牦牛METTL14蛋白二级结构预测
2.4.8METTL14三级结构预测 使用SWISS-MODEL软件对牦牛METTL14氨基酸序列进行同源建模(图11),全局模型质量估计(global model quality estimation,GMQE)值为0.68,发现牦牛METTL14三级结构主要由α-螺旋、无规则卷曲、β-转角和延伸链构成。
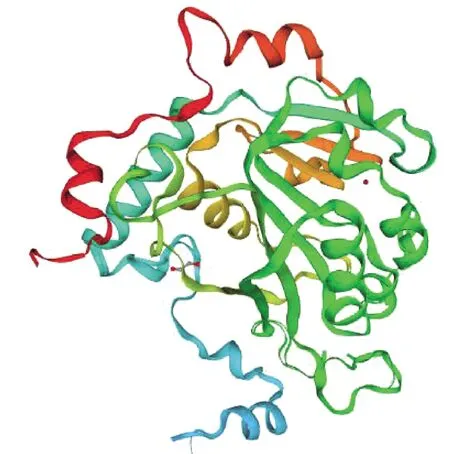
图11 牦牛METTL14蛋白三级结构预测
2.4.8METTL14蛋白互作网络预测 通过STRING软件构建了METTL14互作蛋白网络,结果如图12所示,牦牛METTL14相互作用的蛋白有10个,包括作用于DNA(或RNA)甲基化过程的METTL3、METTL4等MT-A70家族蛋白,参与修复被甲基化损伤的DNA(或RNA)的ALKBH1、ALKBH5等AlkB家族蛋白,以及WTAP、ENSBTAG00000002641、KIAA1429、YTHDF1、CBLL1等其他参与m6A甲基化修饰的相关蛋白。METTL14在这个互作网络中有RNA甲基转移酶活性、RNA结合和催化等分子功能,与各互作蛋白共同参与RNA的甲基化修饰、转运加工、剪接等生物过程。
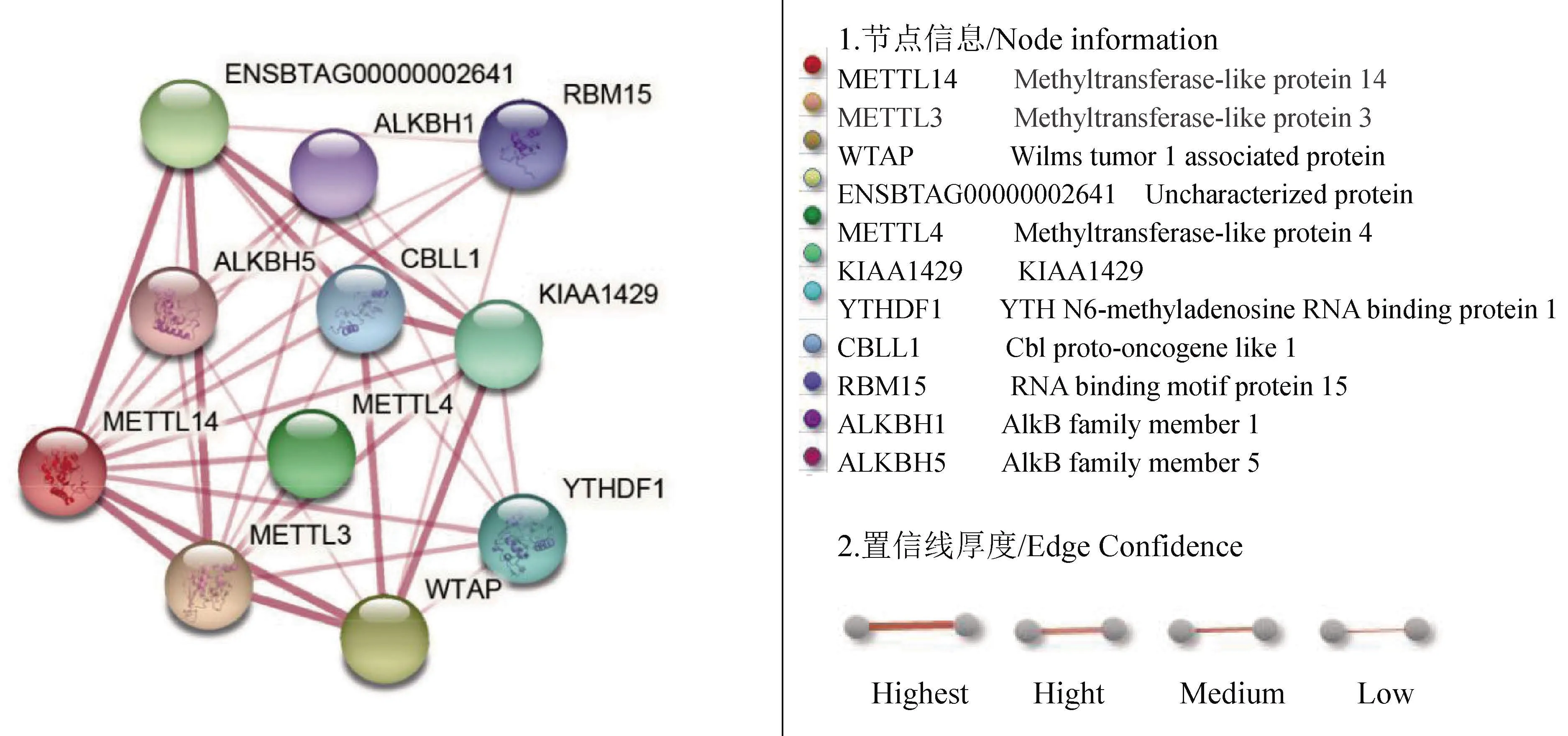
图12 牦牛METTL14蛋白互作网络预测
2.5 METTL14表达谱分析qRT-PCR结果显示,METTL14 mRNA在牦牛心脏、肝脏、脾脏、肺脏、肾脏、背最长肌和皮下脂肪组织中都有所表达,但存在一定的差异,METTL14在肝脏高表达,在脾脏则低表达;与脂肪组织中的表达量相比,METTL14在脾脏的表达量最低,差异极显著(P<0.001);在心脏、背最长肌的表达量极显著高于脂肪组织(P<0.01) (图13A)。牦牛生长发育不同阶段METTL14的表达量也有差异,30月龄牦牛脂肪组织中METTL14表达量显著高于18月龄的(P<0.05) (图13B),表明牦牛METTL14随着个体生长发育的进行,表达也表现出差异。为了进一步探究METTL14在牦牛前体脂肪细胞分化过程中的动态变化规律,诱导牦牛前体脂肪细胞分化并培养12 d,发现随着前体脂肪细胞向成熟脂肪细胞的分化,METTL14在0,4,8,12 d的表达量呈现持续上升的趋势,4,8 d的表达量显著高于0 d (P<0.05),并且12 d的表达量极显著高于0 d (P<0.01) (图13C),表明在脂肪细胞分化后期METTL14发挥着重要的作用。
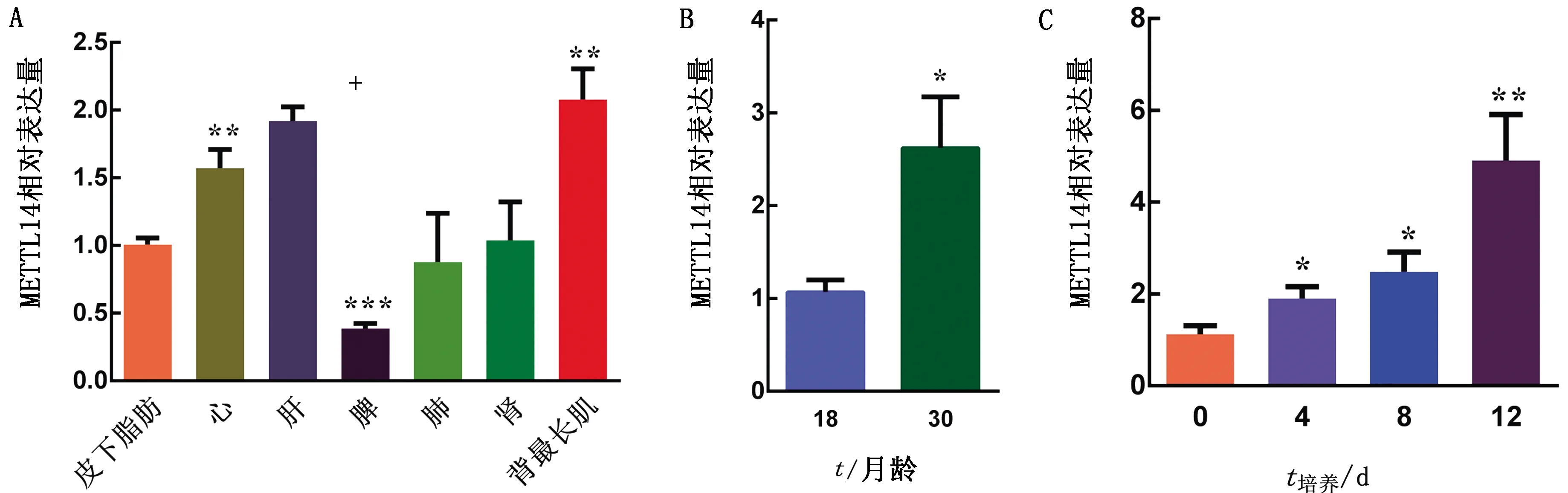
A.牦牛METTL14基因在不同组织中的表达;B.牦牛METTL14基因在2个不同时期的表达;C.牦牛METTL14基因在脂肪细胞分化过程中的表达(0,4,8,12 d)。*.示P<0.05;**.示0.05 本试验经克隆获得牦牛脂肪组织METTL14基因的完整CDS区,由起始密码子ATG开始到终止密码子TAG结束共1 371 bp。大通牦牛METTL14核苷酸与氨基酸序列与其他物种同源性比较结果显示,11个物种之间都有差异但相似性很高,说明METTL14基因结构和功能存在物种及品种特异性,但也保持了酶自身在不同物种中的一致性,进一步说明依赖METTL14功能的m6A在真核生物中广泛存在。大通牦牛METTL14基因和野牦牛亲源关系最近,与鸡和小鼠的亲缘关系最远。这3组进化关系数据说明METTL14具有高度保守性,但存在一定的种属特异性,不同物种间保守性不高。 大通牦牛METTL14基因CDS区共编码456个氨基酸,相对分子质量为52 193.33 kDa,负电荷氨基酸残基总数比正电荷氨基酸残基总数多20个,其-COOH解离程度大于-NH3解离程度,等电点为5.89,表明METTL14为酸性氨基酸;亲疏水性预测结合GRAVY值,可以判断METTL14具有较强的亲水性,有利于其在细胞内发挥活性; METTL14在氨基酸序列186~363位预测到1个MT-A70的保守结构域,此结构域比较接近DNA6mA的甲基转移酶。有研究表明,METTL14是DNA6mA同源物的一个独立亚分支,MT-A70酶结构域和具有替代机制偏好的结构域相连,催化DNA6mA的形成[28]。由此推断,牦牛METTL14中MT-A70的保守结构域维持了甲基转移酶的保守机制;有试验用免疫共沉淀法定位WTAPMETTL3METTL14酶复合物,发现其定位在细胞核上且富集在核斑点上,并一一对复合物的单个蛋白进行核定位信号(nuclear localization signals,NLS)预测,发现METTL14不包含功能性NLS,而METTL3和METTL14构成的异质二聚体在细胞质中形成,细胞质中METTL3的功能性NLS携带METTL14一起转运到核中[29],这与预测的METTL14的信号肽概率为0.092%,几乎不存在引导蛋白质定位到亚细胞内的短肽序列的结果一致,而且METTL14不属于分泌蛋白没有跨膜结构域,所以新合成的METTL14没有分泌通路,无法分泌到细胞外而存在于细胞质中,但本试验METTL14亚细胞定位主要定位在细胞核上,细胞质中有4.3%,核定位预测结果同样有69.60%的该蛋白定位在细胞核上,其可能原因与它的功能相关,METTL14虽然不具有单独的m6A催化活性,但METTL14在细胞核中决定RNA上靶基因的结合,促进METTL3的催化,参与了RNA甲基化修饰。牦牛METTL14潜在磷酸化位点总共有40个,其中苏氨酸上潜在的磷酸化位点数占42.5%,高于丝氨酸(35%)及酪氨酸(22.5%)上的潜在磷酸化位点,磷酸化作用与甲基化作用一样是蛋白质性质改变的方法之一,同样需要通过激活来影响蛋白理化性质;METTL14和METTL3的甲基转移酶结构域紧密结合在一起,形成异质二聚体,METTL3和 METTL14中各有56个和67个氨基酸参与了相互作用,互作界面的氨基酸非常保守,研究中作者根据结构设计了一系列的点突变,但还是很难破坏其稳定结构[30]。但METTL14的不稳定系数为52.02,是不稳定水溶性酸性蛋白,其高级结构以无规则卷曲为主,卷曲的柔性构象会使肽链改变走向,METTL14自身不具备稳定的结构基础,由此推测METTL14单独表达时,疏水互作界面暴露出来,导致蛋白不稳定而与METTL3互作共表达时2个蛋白相互起到稳定异质二聚体构象的作用。在METTL3和METTL14之间有1条富含正电荷的沟槽对RNA 的结合有着重要的功能[30],参与RNA甲基化修饰、RNA转运加工、剪接等生物过程。 METTL14在18龄大通牦牛各组织中均有表达,心脏、背最长肌中的表达高于脂肪组织中的表达,在脾脏表达量最低,表明牦牛METTL14的表达具有组织特异性,与动物代谢活动具有一定的相关性;随着牦牛生长发育,30月龄的牦牛脂肪组织中METTL14表达量高于18月龄的,说明该基因的表达量受时空的影响;METTL14在牦牛前体脂肪细胞中的表达随着前体脂肪细胞向成熟脂肪细胞的发育呈现不断上升的趋势,表明在脂肪细胞分化后期METTL14发挥着重要的作用。本研究从分子水平和细胞水平都表明METTL14基因的表达与脂肪沉积之间存在一定的相关性。 脂肪组织的发育是由一个复杂的转录因子网络调控的过程,除了转录调控外,在以往的研究中发现可通过表观遗传分子机制调控脂肪的发育和脂质的沉积,近年来除了DNA甲基化修饰、组蛋白修饰和染色质空间重塑等DNA和蛋白质上的化学修饰外,RNA修饰日趋成为表观遗传学领域的研究热点[31-32]。新的证据表明,m6A修饰在动物生长发育过程中,对脂肪发育和脂质沉积起着至关重要的作用[33-35]。 m6A修饰位点具有典型的一致序列DRACH(D=G,A或U;R=G或A;H=A、C或U),前体mRNAs(pre-mRNAs)上的m6A可以招募剪接因子两种核糖核蛋白A2和B1(HnRNPA2B1)或加强侧翼RNA序列对剪接因子A2和B1的可变性[36]。据报道,METTL14可以对组蛋白H3赖氨酸36三甲基化(H3K36me3)修饰进行识别,并选择性介导m6A富集在mRNA上[37]。构建Cre/loxP基因敲除小鼠,敲除METTL14会使小鼠胰岛素β细胞中胰岛素分泌量减少,促进肝脏脂肪分解,使脂肪细胞中的脂质沉积减少[38]。m6A依赖METTL14修饰非编码RNA,影响微小RNA microRNAs(miRNAs)的表达和环状RNA(circRNA)的生物合成[39-40],由此可见,理论上机体内脂肪沉积的含量、脂质代谢的过程可以通过METTL14基因进行微调。本试验结果表明,METTL14在大通牦牛脂肪沉积过程中发挥了一定作用,但其作用于脂肪沉积的机制尚未明确。对此,后期可将METTL14作为调控牦牛m6A脂肪沉积的候选基因进一步研究。本试验进一步为METTL14基因在牦牛脂肪沉积调控机制的研究奠定了理论基础。3 讨论
猜你喜欢
上海师范大学学报·自然科学版(2022年3期)2022-07-11
青海湖(2022年3期)2022-06-09
滇池(2022年5期)2022-04-30
汉语世界(The World of Chinese)(2022年1期)2022-02-25
散文诗(2021年22期)2022-01-12
健康之友(2020年1期)2020-03-24
分析化学(2019年3期)2019-03-30
中文信息(2017年2期)2017-04-13
江苏农业科学(2016年11期)2017-03-21
智能计算机与应用(2016年4期)2016-09-26
