
大麦NRAMP全基因组鉴定及重金属胁迫下基因表达 分析
2022-07-22 03:19李一涵于浪柳李春燕张蒙蒙张晓勤方云霞薛大伟
生物技术通报 2022年6期
李一涵 于浪柳 李春燕 张蒙蒙 张晓勤 方云霞 薛大伟
(杭州师范大学生命与环境科学学院,杭州 311121)
多种重金属在维持植物生理和生化功能稳定等方面起着关键作用,如缺少铜(Cu)、锰(Mn)等金属离子将导致植物生长缓慢、植株矮小等现象;而重金属含量过高则又会对植物产生毒害,抑制细胞生长、光合作用等生理活动,进而影响植物的生长发育[1-2]。近年来,因重金属大量排放导致的土壤重金属污染问题愈发严峻,致使农作物产量和质量不断下降。大麦(Hordeum vulgare L.)是种植面积仅次于水稻、玉米、小麦的禾谷类粮食作物,具有食用、饲用、酿酒等多种用途[3]。重金属污染会严重影响大麦正常的生理活动,使其产量下降,因此,研究重金属胁迫下大麦的抗逆机理具有重要 意义。
NRAMP家族是一类重要的膜转运蛋白家族,此类蛋白广泛存在于植物中[4],在调控和维持植物体内金属离子的稳态中发挥着重要作用,可有效缓解重金属离子胁迫对植物生长发育的危害[5]。在锰、铁缺乏条件下,将水稻膜转运家族基因OsNramp6分别导入Fe2+和Mn2+转运缺陷酵母突变体中,能够回复突变菌株的异常表型,表明OsNramp6参与植物体内铁和锰的转运[6]。在拟南芥中,AtNramp1的功能缺失突变体在低锰条件下生长减缓且锰积累量降低,可见AtNramp1是低锰条件下锰吸收的关键转运蛋白[7]。深入研究表明,NRAMP蛋白不仅对锰、铁等微量元素的吸收分配发挥作用,还参与了植物对镉(Cd)等重金属的吸收和转运途径中[8]。
目前已在拟南芥[9]、水稻[5]、大豆[10]、甘草[11]中分别鉴定到6、8、8和11个NRAMP基因,其调控吸收、转运重金属功能在模式植物拟南芥和水稻中研究最为深入,但大麦NRAMP基因的研究却鲜有报道。
本研究采用生物信息学方法鉴定大麦NRAMP基因家族,对该家族成员的序列特征、基因结构、系统发育以及差异表达等方面进行了分析,并研究Mn2+、Cd2+、Cu2+三种重金属离子胁迫对大麦幼苗的生理生化指标及NRAMP家族基因表达的影响,以期了解上述3种重金属离子与NRAMP家族基因表达调控之间的作用机制,为进一步明确NRAMP家族基因对大麦抗重金属胁迫的作用提供依据。
1 材料与方法
1.1 材料
大麦品种Golden Promise(GP)。
1.2 方法
1.2.1 材料处理 将大麦种子用0.1% H2O2消毒10 min,蒸馏水冲洗3-4次,置于暗处萌发24 h。种子萌发后将其转移至培养盒,26℃恒温培养室中水培(光周期14 h/10 h,光合有效辐射18 000 lx)。待幼苗长至一叶一心,进行重金属离子胁迫处理。试验设4个处理组,对照组用1/2 Hoagland培养液[12]培养,试验组分别用含50 μmol/L MnCl2、50 μmol/L CdCl2、50 μmol/L CuCl2的1/2 Hoagland培养液进行胁迫处理,每个处理设置3个重复。胁迫1、3和5 d后,选取幼苗叶片液氮冷冻,并于-80℃保存,用于总RNA提取;同时,胁迫处理3 d时取样,用于生理指标 测定。
1.2.2 生理生化指标测定 丙二醛(malondialdehyde,MDA)含量、过氧化物酶(peroxidase,POD)活性与脯氨酸(proline,Pro)含量均采用试剂盒(苏州科铭生物技术有限公司)测定,具体测定步骤见说明书。
1.2.3 总RNA提取和cDNA的合成 利用RNA植物提取试剂盒(TIANGEN)提取叶片总RNA,再按照反转录试剂盒(ReverTra Ace qPCR RT Kit,TOYOBO)中的方法将样品总RNA反转录合成cDNA第一条链。产物用于实时荧光定量PCR(quantitative real-time polymerase chain reaction,qRTPCR)或-20℃保存备用。
1.2.4 大麦NRAMP基因家族的生物信息学分析
1.2.4.1 NRAMP家族基因的鉴定和结构分析 利用HMMER在线获取大麦的NRAMP蛋白候选序列,再借助NCBI Conserved Domains(https://www.ncbi.nlm.nih.gov/cdd/)和SMART(http://smart.embl-heidelberg.de/)对候选序列进行筛选和去冗余,以获得非冗余的大麦NRAMP基因及蛋白序列。借助Ex-PASy(https://web.expasy.org/protparam/)在线软件预测理论氨基酸长度、理论等电点、相对分子质量等[13]。从EnsemblPlants(http://plants.ensembl.org/ index.html)数据库中获取基因在染色体上的位点信息,并分别利用MG2C(http://mg2c.iask.in/mg2c_v2.0/)、MEME(http://meme-suite.org/)及Gene Str ucture Display Server 2.0(http://gsds.gao-lab.org/)在线绘制染色体定位图、大麦NRAMP蛋白的保守基序和NRAMP基因家族成员的外显子和内含子结构模式图。利用Cell-PLoc 2.0(http://www.csbio.sjtu.edu.cn/bioinf/Cell-PLoc-2/)对大麦NRAMP蛋白进行亚细胞定位预测。
1.2.4.2 NRAMP基因家族进化树的构建 利用MEGA7软件中的邻接法(neighbour-joining)对10个NRAMP基因、6个来自NCBI已确定功能的拟南芥AtNramp基因及7个水稻OsNramp基因的蛋白序列进行系统进化树构建。模型选择为p-distance,校验参数(Bootstrap)设置为1 000,其余参数保持 默认[14]。
1.2.5 大麦NRAMP基因表达谱绘制及候选基因的 筛选 在IPK(https://galaxy-web.ipk-gatersleben.de/) 网站下载相应RNA-seq数据,包括EMB、ROO1、LEA、INF1、INF2、NOD、CAR5、CAR15等多个阶段的表达量信息。将FPKM值作为NRAMP基因的表达谱数据源,核对整理后采用软件MeV绘制表达量热图,依据表达量筛选候选基因[15]。
1.2.6 NRAMP基因的表达分析 遵循荧光定量PCR的引物设计原则设计10对引物(表1)。利用引物定量分析10个NRAMP基因的表达量,相对表达量用2-ΔΔCt(Livak法)计算,每个样品重复4次。根据qRT-PCR的试验结果,用SigmaPlot v10.0作图,并利用IBM SPSS Statistics 20将每组数据分别与对照组进行2个独立样本t检验 (*表示P<0.05,**表示P<0.01)。

表1 荧光定量PCR扩增引物Table 1 Primers for qRT- PCR analysis
2 结果
2.1 重金属离子胁迫对大麦幼苗生理指标的影响
POD的活力变化反映植株受到的胁迫状况。3种重金属分别处理大麦幼苗3 d后,Mn2+、Cd2+和Cu2+处理组的POD活性比对照组分别提高了9.83、9.17和2.36倍,其中,前2组的POD活性差异达到了极显著水平(图1-A),表明不同重金属离子胁迫处理对大麦幼苗POD活性均有影响,但影响幅度不同,其中,Mn2+和Cu2+胁迫处理对大麦幼苗POD活性的影响最显著。MDA是反应细胞膜脂过氧化及膜结构受损程度的重要指标。通过测量处理3 d的大麦幼苗中的MDA含量,Mn2+、Cd2+和Cu2+处理组的MDA含量均明显提高(图1-B),表明3种重金属均对幼苗的膜结构造成了损伤。Pro含量测定结果(图1-C)显示,胁迫处理3 d后,3个处理组的Pro含量均有所提高,其中,Cu2+处理组Pro含量显著升高,Mn2+处理组Pro含量差异极显著增加。由此可见,3种重金属离子胁迫处理均导致大麦幼苗通过大量积累脯氨酸来降低重金属毒害,提高自身的抗逆能力。

图1 重金属离子胁迫处理对大麦幼苗叶片生理指标的影响Fig. 1 Effects of heavy metal ion stress on the physiological indexes of barley seedling leaves
2.2 NRAMP基因家族的生物信息学分析
2.2.1 NRAMP基因家族的分子特征 通过冗余序列的剔除以及二次结构的校验,共得到10条大麦NRAMP蛋白序列,确定其为NRAMP基因家族成员,将其命名为HvNramp1-HvNramp10。对HvNramp1-HvNramp10进行理化性质分析发现,除HvNramp5、HvNramp7、HvNramp9外,其余HvNramp氨基酸长度与分子量均接近,分别介于515-606 aa和55.88-66.31 kD。水稻NRAMP基因家族中的7个基因编码分子量为55.8-59.7 kD,具有500-588 aa[16]。表明大麦HvNramp与水稻OsNramp成员在理化性质上较为相似(表2)。HvNramp蛋白所有成员均有5-12个跨膜结构域,9个HvNramp蛋白定位于细胞质膜,1个HvNramp蛋白(HvNramp8)定位于细胞核 (表2),表明大麦的HvNramp成员可能普遍参与重金属的跨膜转运。

表2 大麦NRAMP蛋白理化性质及染色体定位Table 2 Physicochemical properties and chromosome location of NRAMP protein in barley
2.2.2 NRAMP基因家族成员保守域序列分析 通过对HvNramp蛋白序列保守性分析(图2-A),得到了10个保守基序。HvNramp蛋白分别含1-9个基序,基序数目为9的成员有6个,表明多数基因保守域完整且相似,可能存在相似的功能。此外,除HvNramp8外,其余所有成员都含有基序1和基序2。
2.2.3 NRAMP基因家族成员鉴定及系统发育分析 利用10条HvNramp蛋白序列、6条拟南芥蛋白序列及7条水稻蛋白序列共同构建进化树(图2-B)。根据拟南芥NRAMP蛋白分组[17]及蛋白保守基序的相似性将进化树分为5个亚家族,依次命名为亚家族I-V。其中,HvNramp8的保守基序较为特殊,且其在进化树中单独处于一个分枝,故将其单独列为一个亚家族;亚家族 II中包括大麦HvNramp10、拟 南 芥AtNramp1、AtNramp6及 水 稻OsNramp3;HvNramp5、HvNramp7的保守基序较为相似,属于同一亚家族;亚家族 IV中包括大麦HvNramp1、HvNramp4、HvNramp9及 水 稻 中 的OsNramp1、OsNramp4、OsNramp5、OsNramp6;亚家族Ⅴ中包括大麦中的HvNramp2、HvNramp3、HvNramp6,拟南芥中的AtNramp2、AtNramp3、AtNramp4、AtNramp5及水稻中的OsNramp2、OsNramp7。
2.2.4 NRAMP基因家族成员的外显子和内含子结构分析 内含子-外显子的结构可能会影响到潜在的特定蛋白质的结构[18]。大麦NRAMP家族基因内含子较长,外显子较短,为展现基因结构,手动去除了过长的内含子冗余序列后绘制基因结构图(图2-C)。内含子的长度与相应基因的表达水平有着较强的负相关性,低水平表达基因的内含子长度大于高水平表达基因的内含子。基因结构与亚家族分类结果有一定的相关性,除亚家族Ⅲ的HvNramp5、HvNramp7分别含有5和1个外显子差异较大外,其余亚家族基因外显子数量均相近,如亚家族 IV的HvNramp1、HvNramp4、HvNramp9分 别 含 有8、12和9个外显子;亚家族Ⅴ的HvNramp2、HvNramp3、HvNramp6分别含有4、3和4个外显子。
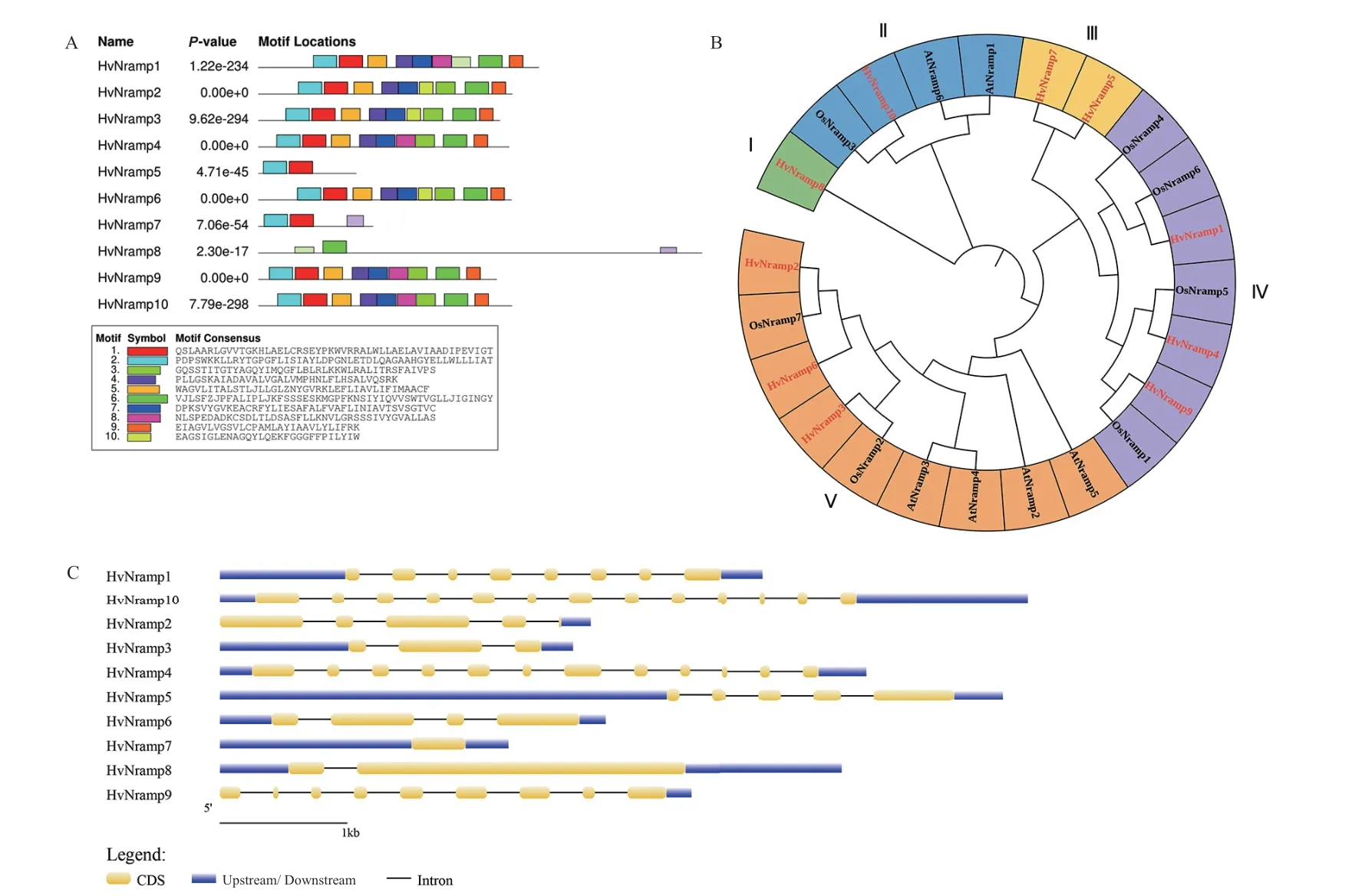
图2 大麦NRAMP基因家族蛋白保守基序、基因结构及系统进化分析Fig. 2 Conserved sequences,gene structure and phylogenetic analysis of barley NRAMP gene family
2.2.5 NRAMP基因家族成员的表达量预测分析 利用10个大麦NRAMP基因在不同组织中的表达数据,绘制热图,探究NRAMP家族成员的表达特征。结 果(图3)表 明,HvNramp1、HvNramp3、HvNramp6、HvNramp7、HvNramp8在各个组织和果实的不同发育阶段中均有较高程度的表达,被聚类为 高 表 达;而HvNramp2、HvNramp4、HvNramp5、HvNramp9、HvNramp10表达量相对较低,被聚类为低表达。HvNramp表达具有组织差异,在胚胎、根及叶中多数高表达,而在花序组织中表达较低。

图3 大麦NRAMP基因的表达谱分析Fig. 3 Expression profile analysis of NRAMP gene family in barley
2.2.6 NRAMP基因家族成员的染色体定位分析 根据EnsemblPlants上获得的NRAMP基因位置信息,绘制了基因的染色体定位图(图4),10个HvNramp分布于大麦的3H、4H、5H、7H四条染色体上且多集中在染色体近末端。分布在第4染色体上的基因最多,为4个;分布在第3染色体上的基因最少,仅有1个。着丝粒和近着丝粒区域易发生序列变化和结构重塑,是植物新基因发生和起始的热点区,并且着丝粒区域内的基因有着向外逃离的进化趋势[19]。本研究中所有NRAMP基因都位于远着丝粒区。由此推测大麦NRAMP基因进化历程较长,功能较为稳定。

图4 大麦NRAMP家族基因在染色体上的分布Fig. 4 Distribution of barley NRAMP family genes on chromosomes
2.3 NRAMP基因家族成员在重金属离子胁迫条件下的表达分析
NRAMP编码的蛋白具有吸收和转运重金属的功能,为对筛选基因进行重金属逆境胁迫验证,利用qRT-PCR对Mn2+、Cd2+、Cu2+胁迫下大麦幼苗中10个HvNramp的表达进行检测,因HvNramp6、HvNramp7、HvNramp10在大麦幼苗叶片中的表达过低,最终仅对其余7个基因的相对表达量进行了分析。
qRT-PCR分析结果显示,Mn2+胁迫下,HvNramp1、HvNramp2的表达量显著升高,分别提高了1.33和1.10倍;Cd2+胁迫下,HvNramp8的表达量下降了16%,与对照间存在极显著差异,其他基因表达量虽然略有提高,但与对照间的差异不显著;而Cu2+胁迫时7个基因表达量均上调,其中HvNramp1、HvNramp2、HvNramp5、HvNramp9在Cu2+胁迫下表达量显著升高,分别提高了2.93、4.48、1.03和3.51倍(图5)。结果表明,除了HvNramp3 外,其他6个HvNramp均是大麦苗期响应重金属胁迫的基因。

图5 3种重金属离子胁迫下7个大麦NRAMP家族基因相对表达分析Fig. 5 Relative expression analysis of seven barley NRAMP family genes under three heavy metal ion stresses
3 讨论
重金属离子刺激植物产生应激反应,使其生成较多的过氧化物酶、过氧化氢酶与超氧化物歧化酶等抗氧化酶,植物的抗氧化酶可清除过多的氧自由基,通常作为植物抵抗重金属胁迫的重要指标[20]。本研究中,3种重金属离子处理下的大麦幼苗叶片中POD活性较对照组均显著提高,尤其以Mn2+、Cd2+胁迫组升高最显著。表明3种重金属离子处理激发了大麦的抗氧化反应,刺激抗氧化酶的产生以去除重金属胁迫产生的氧基自由基,其中POD活性对锰、镉胁迫较敏感。MDA是植物受到逆境胁迫后膜脂过氧化的产物,MDA含量的高低能够反映细胞膜脂过氧化的程度和植物受逆境伤害程度的强弱[20]。本研究中,与对照组相比,各处理组的MDA含量均显著高于对照组,表明重金属离子胁迫处理对大麦幼苗具有毒害作用,其中Mn2+、Cd2+胁迫的毒害作用更强,Cu2+胁迫的毒害作用相对较弱。植株积累的脯氨酸可以通过渗透调节增强自身的抗逆性[21],其在一定程度上能够衡量植物抗逆性,含量越高抗性越强。本研究中,处理组大麦幼苗叶片的Pro含量均高于对照组,其中Mn2+处理时上升最显著,Cu2+次之,Cd2+最低。3种重金属离子处理组中大麦幼苗Pro含量均有显著上升,这表明在重金属离子胁迫条件下,大麦幼苗通过大量积累脯氨酸来降低重金属毒害,从而提高自身的抗逆能力。
大麦对重金属离子胁迫的应答过程中发生的一系列生理生化变化,是由复杂的内在分子机制来调控的。本研究在大麦全基因组范围内共鉴定到10个NRAMP基因,对该家族成员进行理化特性分析发 现,蛋白的保守基序有1-9个不等,且不同的HvNramp蛋白序列、氨基酸长度均有较大的差异。已有研究表明,NRAMP家族蛋白的亚细胞定位位置基本可以分为两类,其一主要定位于细胞质膜,负责元素的吸收;其二主要定位于液泡膜,负责元素的再利用[19]。亚细胞定位预测9个基因定位于细胞质膜,推测HvNramp主要与重金属离子的跨膜转运相关。先前有研究表明NRAMP家族基因编码膜整合蛋白分子,一般具有10-12个跨膜区TM,且TM1-8具有高度同源性[22],但在最新研究中发现甘蔗NRAMP基因家族蛋白跨膜结构域数位于6-12的情况[23],这与本研究中大麦HvNramp蛋白跨膜结构域数较为相近。已有研究发现,拟南芥AtNramp1、 AtNramp2、AtNramp3、AtNramp4[7,24-26]均 对Mn2+的吸收和转运起到关键作用,水稻OsNramp1、OsNramp2、OsNramp5、OsNramp6[7,27-29]均 与Cd2+的吸收转运相关。本研究中,在Mn2+胁迫下,HvNramp1、HvNramp2表达水平显著上调,暗示它们可能参与Mn2+的吸收和转运;在Cd2+胁迫下,除HvNramp8表达量显著下调外,其余基因均上调,这与先前的结果相似[30],表明大麦HvNramp可能参与Cd2+的吸收与转运;在Cu2+胁迫下,发现6个基因表达量均显著上调,其中,HvNramp1、HvNramp2、HvNramp5的响应最强烈,暗示NRAMP基因家族可能还参与Cu2+的吸收与转运,在水稻中也有类似的报道[31]。因此,2个Mn2+胁迫下的显著上调基因、6个Cd2+胁迫下的上调基因、6个Cu2+胁迫下的显著上调基因可初步认定为大麦HvNramp抵御重金属胁迫的相关基因。近年来,土壤重金属污染形势愈发严峻,重金属胁迫会对植物体造成毒害作用,NRAMP基因家族相关的转基因植物特别是农作物可能得到广泛研究,有较好的应用前景。
4 结论
在50 μmol/L的Mn2+、Cd2+、Cu2+胁迫处理下,大麦幼苗体细胞内可能出现了积极的保护作用来抵御重金属离子的毒害,大麦NRAMP基因家族通过吸收和转运重金属离子来调控和维持植物体内金属离子的稳态,对大麦抗重金属胁迫具有重要作用。
猜你喜欢
作文小学高年级(2022年5期)2022-06-16
南方医科大学学报(2022年3期)2022-04-13
今日农业(2021年10期)2021-11-27
环境卫生工程(2021年4期)2021-10-13
浙江大学学报(农业与生命科学版)(2021年3期)2021-07-10
疯狂英语·新读写(2021年2期)2021-02-25
中国预防兽医学报(2020年8期)2020-11-05
三农资讯半月报(2020年15期)2020-08-25
华人时刊(2016年19期)2016-04-05
中国火炬(2015年12期)2015-07-31
