
珠江口红树林土壤甲烷和二氧化碳通量特征及其影响因素研究
2022-07-22 06:40张涵唐常源禤映雪江涛黄品怡杨秋曹英杰
生态环境学报 2022年5期
张涵,唐常源,*,禤映雪,江涛,黄品怡,杨秋,曹英杰
1. 中山大学地理科学与规划学院,广东 广州 510275;2. 中山大学环境科学与工程学院,广东 广州 510275;3. 中山大学/广东省环境污染控制与修复技术重点实验室,广东 广州 510006;4. 南方海洋科学与工程广东省实验室(珠海),广东 珠海 519080
甲烷(CH4)和二氧化碳(CO2)是大气中最重要的温室气体,两个多世纪以来,CO2在大气中的摩尔浓度由工业革命前的 280×10−6增长到409.5×10−6(Lüthi et al.,2008),国际普遍认为 CO2是造成全球气候变暖的主要原因,虽然CH4在空气中含量较低,但是其在大气中寿命较长,远红外吸收容量大,单个分子的CH4增温潜势是CO2的21—33倍(Church et al.,2013),对全球变暖的贡献也不可忽视。在陆地生态系统碳循环成为全球变化研究热点的背景下,红树林以其碳循环周期短和初级生产力极高的特点,受到国内外学者的广泛关注(Kristensen et al.,2008)。一方面,红树林虽然仅占陆地面积的0.1%,但其土壤有机碳含量超过陆地生态系统总有机碳含量的10%(Alongi,2014;Duarte et al.,2005;McLeod et al.,2011),是所有森林中最富集的碳汇(Bhomia et al.,2016;Donato et al.,2011;Murdiyarso et al.,2015)。另一方面,红树林土壤中的有机碳经过矿化作用产生 CO2和 CH4(Allen et al.,2007;Chen et al.,2016),是不可忽视的碳源。开展红树林土壤CH4和CO2排放通量及其影响因素研究对认知典型陆地森林生态系统碳源汇效应具有重要意义。
红树林湿地土壤被潮水周期性淹没,湿地坡度不同位置的淹没时间长度存在差异(Marchand et al.,2012),土壤淹没和排水(不饱和)条件下,气体的扩散不同,直接影响了温室气体向大气的排放,同时也间接导致了土壤一系列性质的差异,包括土壤含水量、有机质含量、盐分等等(Jacotot et al.,2018;李森等,2020)。已有的研究表明,土壤有机质是温室气体产生所需要的底物和能量的主要来源(徐华等,2008),土壤CH4与CO2排放量往往与有机质含量呈正相关(宋长春等,2004)。土壤含水量通过改变土壤的氧化还原电位、透气性以及温室气体的扩散速率来影响气体的产生与排放,在氧化还原电位较低的条件下,有机质分解不彻底,有机物的厌氧分解占主导(Maier et al.,2011)。由于硫酸盐还原菌比产甲烷菌具有更高的竞争率,盐度往往与CH4排放呈反比关系,当土壤盐度大于18时,土壤的CH4排放将受到限制(Biswas et al.,2007;Poffenbarger et al.,2011)。土壤的水分状况对CH4的产生和氧化有一定影响(张晓艳等,2016),当土壤浸没时,沉积物底部的CH4大部分溶解于孔隙水中,潮水退去土壤不饱和时,CH4会从沉积物扩散至大气中,剩余的CH4则通过气泡的形式排出(Neue et al.,1994)。
稳定同位素分馏贯穿于生态系统的生物、物理、化学过程中,能够在时间和空间尺度上反映生态过程对环境条件变化的响应,广泛用于量化与碳循环相关的生态过程(Diao et al.,2022;姜懿珊等,2020)。土壤呼吸以根系呼吸和土壤微生物的呼吸为主(Hanson et al.,2000),土壤呼吸产生的CO2来源于有机质的分解和植物根的呼吸,由于土壤有机质主要来源于群落内植物的凋落物,因此土壤呼吸产生的 CO2的δ13C值受所在区域植物群落的影响。由于不同类型植物的δ13C值差异明显,导致不同植被的土壤呼吸产生的 CO2的δ13C值也存在显著差异(Bender,1971;Farquhar et al.,1982)。CH4的δ13C值与CH4的生产过程有关,乙酸发酵产生的CH4的δ13C 值通常介于−65‰— −50‰,而氢气还原 CO2产生的 CH4的δ13C 值介于−110‰— −60‰(Vaughn et al.,2016)。将碳稳定同位素同静态箱技术结合,可以研究土壤呼吸产生的CO2和CH4来源和途径问题。
因此,本研究选择广州南沙区的红树林潮汐湿地,开展红树林土壤CO2和CH4的排放通量时空变化特征及影响因素研究,并结合稳定同位素示踪法探究CH4和CO2的来源,为深入开展红树林温室气体研究提供数据支持,并为保护和恢复红树林生态系统提供参考。
1 采样和分析
1.1 研究区概况
研究区(图1)位于广州市南沙区虎门轮渡码头附近(113°34′33″—113°34′47″E,22°48′44″—22°49′02″N),属南亚热带海洋季风气候,雨热同期(李海生等,2020)。图2所示为采样期间的气温和降水量变化,研究区年均降水量1933 mm,4—9月降雨量占全年降雨量的81%,年均气温23 ℃,年总辐射量为 4400—5000 MJ·m−2·a−1。研究区的潮汐湿地为珠江的狮子洋和伶仃洋潮汐长期冲刷而成,坡面总长度约为40 m,平均坡度0.1°,潮汐性质为不正规半日潮,基准面在平均海平面下140 cm,最高潮位为252.88 cm,最低潮位为0 cm,平均潮差约1.37 m。研究区现存红树林45.5 hm2,主要植物有桐花树(Aegiceras corniculatum)、老鼠簕(Acanthus ilicifolius)、无瓣海桑(Sonneratia apetala)、水黄皮(Pongamia pinnata)等(康文星等,2008;李海生等,2020)。采样区以桐花树群落为主,平均树高为3.09 m,平均干径为7.36 cm。

图1 南沙红树林研究区分布位置Figure 1 Locations of sampling sites in mangrove in Nansha

图2 采样期间气象记录Figure 2 Meteorological records during sampling period
1.2 研究方法
1.2.1 样品采集
观测时间为2020年8月—2021年7月,观测频率为每月一次。在潮汐湿地垂直岸线方向选取外滩、中滩、内滩3个采样点。外滩为没有植被覆盖的淤泥裸滩,中滩植物较多土壤紧实,内滩为有机质含量较少的沙土。每个采样点设置 0.5 m×0.5 m样方,保证地表无明显植物根系、幼苗和凋落物。研究区各点通量观测采用静态箱法(Chen et al.,2010)。静态箱由有机玻璃制成,内置微型风扇,箱侧设气体收集孔。采样前将静态箱插入土壤约 3 cm,确保采样箱处于密闭状态。从0时刻开始,每隔10分钟采一个样,共计30 min,共收集4个气体样品,用于土壤呼吸通量的计算,每次采样后,记录静态箱覆盖土壤的温度(℃)。采样时用微型真空泵将箱内气体导入300 mL铝箔采气袋,每次抽气前2 min打开风扇,将箱内空气搅匀。
1.2.2 实验分析
每个季度采集一次表层土壤样品,去除土壤表层的植物和根系,带回实验室使用烘干法测量含水量,去除风干土壤中的有机质和碳酸钙后,用Mastersizer激光粒度仪测量土壤的粒径。每次采样过程中,抽取采样点处的土壤水,用哈希探头测定溶解氧(DO)、酸碱度(pH)、电导率(EC)、氧化还原电位(ORP),溶解性无机碳(DIC)、硫酸根离子(SO42−)和硝酸根离子(NO3−)由离子色谱测得,各个指标测3次,取平均值。野外观测发现中滩和内滩有藻类出现,取藻类样品开展藻种分析。
1.2.3 CO2/CH4含量和同位素测量
所有气样在采集后24 h内,利用碳稳定同位素分析仪Picarro G2201-i,测定样品中CO2和CH4的摩尔浓度以及碳稳定同位素组成(δ13C-CH4和δ13CCO2),其中 CO2和 CH4的摩尔浓度的精度分别是210×10−9+0.05%和 60×10−9+0.05%,δ13C-CO2的精度小于±0.16‰,δ13C-CH4的精度小于±1.15‰。每次测样前,使用标准气体检查仪器的稳定性。每个气体样品测试10 min,剔除前2 min因换样导致的不稳定数据,计算后8 min CO2和CH4的摩尔浓度以及δ13C-CH4和δ13C-CO2的平均值。13C测定结果以相对于VPDB(Vienna Pee Dee Belemnite)的千分率(‰)给出,用δ13C表示,公式为:

式中:
Rsample和Rstandard——样品和标准物(VPDB)的13C和12C的丰度比。
土壤中产甲烷方式主要分为氢气还原产甲烷过程和乙酸发酵产甲烷过程(Whiticar et al.,1986),利用同位素分馏系数αc可以衡量土壤中主要的产甲烷方式,分馏系数αc的计算公式如下(张尹等,2020):
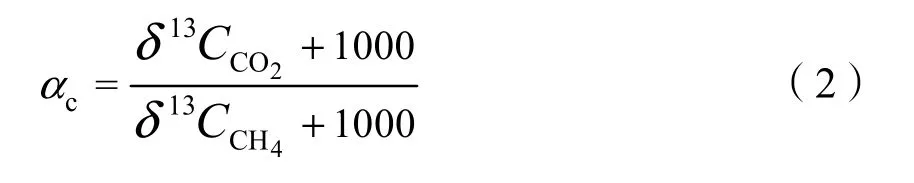
式中:
δ13C——CO2和CH4的碳稳定同位素比值,当αc<1.055时,产甲烷方式以乙酸产甲烷为主,当αc>1.055时,产甲烷方式以氢气还原二氧化碳为主。
1.2.4 温室气体通量及增温效应的计算
CO2和 CH4通量通过静态箱内摩尔浓度随时间的线性回归计算(Jacotot et al.,2018),使用以下方程:

其中:
F——CO2和 CH4的通量(mol·m−2·h−1);
d[c(CO2,CH4)]/dt——CO2或CH4的物质的量浓度随时间的变化率(10−6mol·mol−1·min−1);
V——静态箱的总体积(m3);
S——静态箱罩在土壤上的面积(m2);
R——摩尔气体常数 8.205746×10−5(atm·m3·K−1·mol−1);
T——静态箱内的平均温度(K);
60——换算系数。
将测得的温室气体通量转换为CO2当量通量以表示它们的增温效应,在100 a时间尺度内,CH4和CO2的全球增温潜力系数(GWP)分别是 28和 1(IPCC,2014)。CO2当量通量的计算公式为:

其中:
Fe——CH4的 CO2当量通量(kg·hm−2·d−1);
F——甲烷的通量(μmol·m−2·h−1);
M——CH4的摩尔分子质量 16 g·mol−1;
KGWP——甲烷的增温潜力系数28;
2.4e−4——转换系数。
1.2.5 Keeling Plot
Keeling在美国Mauna Loa等地对CO2浓度季节变化的连续监测结果表明,当进入空气中的CO2的碳同位素丰度与初始空气中的二氧化碳的丰度不同时,大气中13C-CO2碳同位素比值的变化与CO2摩尔浓度的倒数成正比(Keeling,1958),其拟合直线与垂直轴的截距δ13Cs表示生态系统中CO2排放源的δ13C值。基本原理是质量守恒:

式中:
cz、cx和cs——所测大气中的浓度、本底大气的浓度和土壤排放/吸收的气体的浓度(×10−6)。结合13C同位素,构建质量守恒方程,变形后得到以下公式:

式中:
w(δ13C)——各部分的碳同位素值(‰),下标含义与浓度cz、cx和cs相同,气体浓度倒数与同位素值之间的线性回归截距可以作为来源向原始空气中增加的气体的碳同位素的整合。
由于静态箱隔绝了箱内气体与大气的交换,且箱内没有植物,箱内增加的CO2浓度全部由土壤呼吸所产生,因此可将静态箱与Keeling曲线法结合起来,求得土壤呼吸产生的CO2的δ13C值。
1.3 数据处理和分析
应用IBM SPSS 22和Origin 2018c进行数据分析和绘图,采用单因素ANOVA检验对数据的空间差异进行显著性分析。采用独立样本t检验比较通量和各环境因子之间的差异性。计算理化因子与通量的相关性系数(Spearman),判断影响温室气体通量的因素。差异性和相关性分析的显著性水平均为P<0.05。
2 结果
2.1 气象及土壤理化因子
外滩、中滩、内滩采样点气象及土壤理化因子指标统计结果见表1。外滩、中滩和内滩的 EC、ORP、NO3−、Cl−和土温没有显著差异,pH、DIC、SO42−、土壤含水量、OC、容重在外滩、中滩和内滩有显著差异。研究区土壤pH范围为6.5—8.3,呈中性偏碱,外滩的pH值显著高于中滩和内滩,研究区土壤水的 EC 均值为为 10.79×103μS·cm−1,ORP均值为137.73 mV,内滩土壤孔隙水的DIC质量浓度均值为 2346.21 g·L−1,显著高于中滩(579.52 mg·L−1),外滩的土壤含水量最高,达到饱和状态,内滩最小,中滩的有机碳含量和容重最高,有机碳质量分数达到了 46.97 g·kg−1,中滩容重为 1.38 g·cm−3。研究区土壤水中 NO3−的平均质量浓度为13.47 mg·L−1,Cl−平均质量浓度为 3845.61 mg·L−1,中滩土壤水中 SO42−浓度显著高于外滩和内滩,达到了306.70 mg·L−1,内滩土壤水中DO浓度显著低于外滩和中滩,仅有 3.47 mg·L−1。
粒径分析表明,按照≤2 μm(黏粒)、2—50 μm(粉粒)、≥50 μm(砂粒)将土壤粒径划分为3个等级。3个采样点的土壤以砂粒为主,其中,内滩土壤砂粒含量达到了 100%,中滩和外滩含有少量黏粒和粉粒,中滩土壤粉粒和黏粒和含量分别为7.12%和1.14%,外滩土壤含有的粉粒和黏粒较少,为0.05%和3.85%。
相关性分析表明(表2),pH、EC、ORP、土壤含水量、有机碳含量和溶解氧浓度与CO2通量呈显著负相关,pH、DIC、土壤含水量和有机碳含量与CH4通量呈正相关。本研究证实了土壤排放CO2和CH4通量的季节性差异,土温与CH4和CO2通量没有显著相关性,季节更替除了影响温度外,还会导致湿度和含水量等产生对应的变化,而理化因子分析也证明了外滩、中滩和内滩的理化性质有着较显著的区别。其他指标与CH4和CO2没有显著相关性。
2.2 土壤CO2和CH4通量的时空变化规律
2.2.1 时间变化规律
将外滩、中滩、内滩温室气体的通量求均值,作为各月的平均CH4和CO2通量(图3)。采样期间 CO2通量均值为 4.19 mmol·m−2·h−1,除 10、6、7月以外,平均CO2通量表现为正通量。方差分析表明CO2通量有明显的月变化(P<0.05),6—10月的月均CO2通量显著低于其他季节,其中6月的月均CO2通量最低,为−1.53 mmol·m−2·h−1,3 月的月均CO2通量最高,为 18.91 mmol·m−2·h−1。CH4通量均值为 18.97 μmol·m−2·h−1,各月的 CH4通量均值均表现为正通量,方差分析表明CH4通量在不同采样月份也呈现出明显的差异(P<0.05),3月的月均CH4通量最低,为 0.29 μmol·m−2·h−1,9 月的月均 CH4通量最高,为 120.42 μmol·m−2·h−1。

图3 红树林土壤各月CH4和CO2通量Figure 3 CO2 and CH4 fluxes from the mangroves soils for the different months

表1 外滩、中滩、内滩土壤理化指标Table 1 Physicochemical properties of soil and soil water in seaward, middle and landward
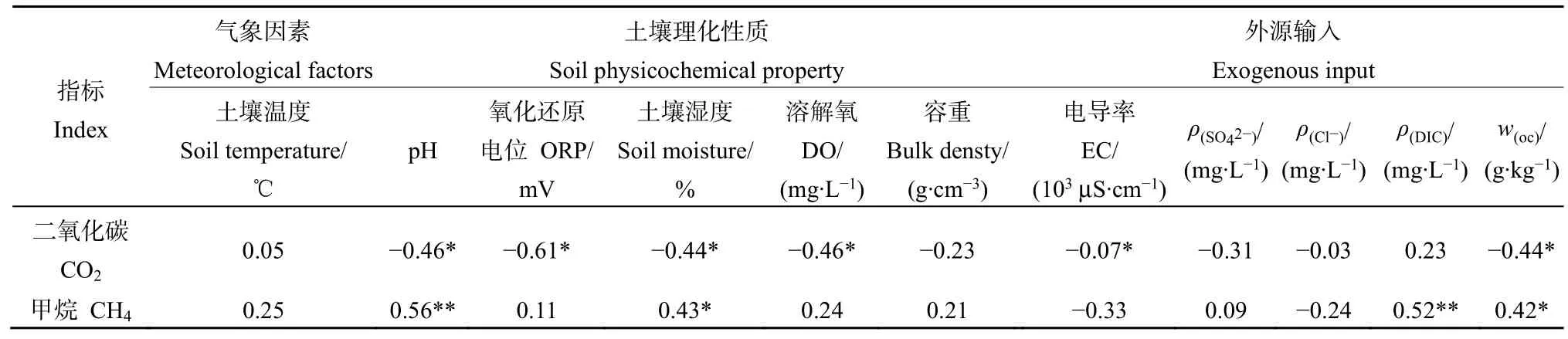
表2 研究区CO2和CH4排放通量与理化指标之间的Spearman相关性Table 2 Spearman correlation between CO2 and CH4 fluxes and physicochemical factors
2.2.2 空间变化规律
外滩、中滩和内滩的CO2通量均值分别为1.10、1.76、14.29 mmol·m−2·h−1(图4a)。方差分析表明,内滩的CO2排放通量显著大于中滩和外滩(P<0.05),中滩和外滩之间没有显著差异(P=0.814)。
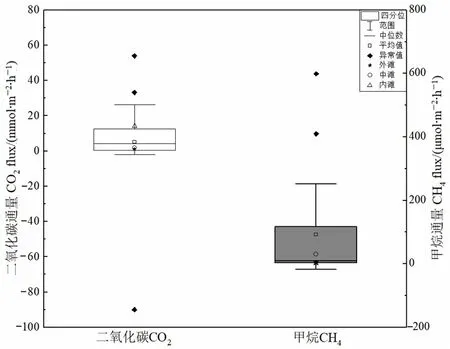
图4 红树林3个采样点的CO2和CH4通量Figure 4 CO2 and CH4 fluxes from the mangrove soils in seaward, middle and landward
外滩、中滩和内滩的CH4通量均值分别为4.62、30.03、1.37 μmol·m−2·h−1(图4b),总体而言,研究区的 CH4通量变化范围为−3.54—148.28 μmol·m−2·h−1。方差分析表明,中滩 CH4通量显著高于外滩和内滩(P<0.05),外滩和内滩的CH4的通量没有显著差异(P>0.1)。
本研究中,内滩、外滩、中滩的CO2通量大部分为正值,CH4通量均为正值,判定红树林土壤是该区域的温室气体来源。本研究区CO2平均通量为4.19 mmol·m−2·h−1,CH4平均通量为 18.97 μmol·m−2·h−1。用增温潜力系数将外滩、中滩、内滩土壤的 CH4和 CO2的平均通量转化为二氧化碳当量通量(表3)。

表3 土壤呼吸温室气体通量的CO2当量通量Table 3 CO2-equivalent fluxes from the mangrove soils in seaward, middle and landward
整合先前对于湿地土壤温室气体通量的研究,并将收集的数据绘制成箱线图(图5)。先前的研究中,CO2的平均通量为 5.17 mmol·m−2·h−1,范围为−1.96—26.21 mmol·m−2·h−1。CH4的平均通量为 92.47 μmol·m−2·h−1,范围为−18.00—252.00 μmol·m−2·h−1。将外滩、中滩、内滩的平均CH4和CO2通量绘制在箱线图中,比较本研究与先前研究的差异。

图5 先前研究所测通量值与本研究对比Figure 5 Comparison of the mangrove soil CO2 and CH4 fluxes of this study with previous studies
2.3 CO2、CH4产生过程的同位素示踪
采样期间,静态箱中气体的 CO2同位素特征(δ13C-CO2)均值为−10.14‰,变化范围为−20.89‰— −1.71‰,12月均值最低,为−16.62‰,9月均值最高,为−3.78‰,方差分析结果表明,δ13C-CO2在不同月份呈现出显著差异。外滩δ13CCO2均值为−11.00‰,中滩δ13C-CO2均值为−8.36‰,内滩δ13C-CO2均值为−12.06‰。方差分析表明,δ13C-CO2在不同采样点呈现出显著差异,中滩δ13C-CO2最高,内滩δ13C-CO2最低。
采样期间,静态箱中气体的 CH4同位素特征(δ13C-CH4)均值为−49.38‰,变化范围为−70.50‰—−25.32‰,9月均值最低,为−62.82‰,12月均值最高,为−41.00‰,方差分析表明,δ13CCH4在不同月份呈现出明显的差异。外滩δ13C-CH4均值为−47.20‰,中滩δ13C-CH4均值为−51.70‰,内滩δ13C-CH4均值为−45.10‰。方差分析表明,δ13C-CH4在不同采样点呈现出显著差异,表现为内滩δ13C-CH4最高,中滩δ13C-CH4最低。
将 CO2浓度和δ13C-CO2拟合成 Keeling曲线(Keeling,1958),结果表明土壤呼吸作用产生的CO2的δ13Cs范围为−34.68‰— −12.18‰,其中,内滩的δ13Cs均值为−16.05‰,中滩的δ13Cs均值为−17.73‰,外滩的δ13Cs均值为−29.77‰(表4)。

表4 Keeling Plot拟合土壤呼吸的δ13C-CO2Table 4 Regression equation and δ13C-CO2 obtained by keeling curve
3 讨论
3.1 研究区土壤CO2、CH4排放强度
温室气体当量通量换算结果表明(表3),CO2占总排放的 97.9%,CH4占总排放量的 2.10%,表明红树林土壤作为温室气体源,其增温潜力以CO2为主导。本研究主要在退潮后进行,土壤处于不淹水状态,有氧呼吸强度大于厌氧呼吸(郝庆菊等,2004),CO2通量高于 CH4。
对比其他红树林湿地温室气体通量的研究发现(n=22,图5),本研究区的CO2、CH4通量在中国的红树林研究中处于较低的水平。由于坦头村红树林位于珠江入海口,循环速度快,且红树林规模较小,因此红树林生物量较小,土壤CO2和CH4的排放通量不及英罗港、东寨港、鳌江镇的红树林。本研究只针对退潮后的土壤,对于同一片区域退潮前后的通量,有研究证明红树林退潮期间土壤CO2通量高于被淹没时期(Jacotot et al.,2019),因为退潮期间土壤中的氧气扩散增强了有机物分解,以及气体在大气中比在水中更容易扩散。
3.2 CO2和CH4通量的变化及影响因素
3.2.1 影响通量变化的气候因子
多项研究表明,土壤温度是影响土壤温室气体排放的重要因素(Mazzola et al.,2022)。一方面,土壤温度升高会提升土壤微生物的活性,促进植物的生长和代谢,进而加速土壤有机质的分解和土壤的呼吸速率(Rustad et al.,2001),另一方面,温度较高的夏季是植物生长的高峰期,根系可能会向土壤中输送更多的有机物,为微生物提供更多的物质基础(Kelley et al.,1995)。本研究区属于亚热带季风气候,总体上温度变化范围相对较小,不足以影响到温室气体的排放。土壤含水量反映了土壤的水分状况,本研究中土壤含水量与CO2呈负相关关系,与CH4呈正相关关系,土壤水分含量较少时,有利于氧气进入土壤,促进土壤有机质的好氧分解,从而加速CO2的释放。当土壤中含水量较高时,土壤内部的厌氧条件有利于产CH4作用的发生(贾庆宇等,2015;杜明智等,2022)。有研究表明,降水量增加会在减少CO2通量的同时增加CH4通量(高春等,2017),在本研究中,9月采样期间出现降雨,降雨增加了土壤含水量,更好的厌氧条件使得9月CH4通量高出其他季节。
3.2.2 影响通量变化的土壤理化因子
土壤pH因对化学反应和酶多样性的调控而对土壤呼吸有明显的影响(Bottner et al.,1998),适宜微生物生存的pH区间为6—8,超过这个范围,微生物的活性会显著下降(谢军飞等,2002)。土壤pH与CO2通量呈负相关,由于研究区pH中性偏碱,位于最适区间的后半段,因此从内滩到外滩随着土壤pH的增加,CO2通量逐渐减少。据报道,沿海湿地甲烷排放的最适pH为7.7(Garcia et al.,2000;Chang et al.,2003)。研究区的土壤pH值与CH4通量呈正相关,其中,内滩的pH为6.9达到最低,对应了内滩最低的CH4通量。外滩土壤pH值最接近7.7,但是外滩的CH4通量低于中滩,证明有其他因素导致中滩更高的 CH4通量。本研究 CO2通量与ORP有显著的负相关关系,氧化还原电位越高表示有机物被分解得较为完全。内滩的氧化还原电位相对较低,意味着有机物未被完全分解,因此内滩的CO2通量显著高于中滩和外滩。相关研究表明,随着水中溶解氧的消耗,水中溶解的CO2会释放更多(Soumis et al.,2004),本研究中溶解氧的浓度与CO2呈负相关,较高的溶解氧浓度反映出溶解在孔隙水中的CO2未被释放。此外,由于内滩地势较高,潮水淹没时间较短,土壤含水量较低,较粗的土壤质地和较低的含水量增加了土壤的通气性,促进了有氧呼吸,从而增加了内滩CO2的释放通量。中滩容重最大,土壤紧实透气性差,良好的厌氧环境从一定程度上促进了CH4的产生。海藻在中滩有分布,在外滩没有海藻分布,但是中滩CO2通量与外滩并无显著差异,因此海藻不是影响土壤CO2排放的主控因素,但是其决定了CO2的来源。此外,藻类在氧化条件下可以利用碳酸氢、二甲基硫醚、二甲基亚砜或蛋氨酸亚砜作为碳前体可以会产生CH4(Liu et al.,2022),对于同样有藻类覆盖的中滩的内滩,中滩 CH4通量显著高于内滩,说明藻类对于土壤CH4通量的影响不显著。
3.2.3 影响通量变化的外源输入
由于产甲烷菌与 SO42−还原菌之间对乙酸和氢的竞争关系,SO42−的质量浓度对 CH4往往有抑制作用(Biswas et al.,2007)。但是Purvaja et al.(2001)发现当水和沉积物中的硫酸盐浓度很高时,红树林河口中CH4的排放量仍然很高,这表明产CH4和硫酸盐还原剂是可以共存的。土壤有机碳含量是CH4产生的重要影响因素,作为CH4产生所需要的底物和能量的主要来源之一,有机质含量与CH4有显著的正相关关系(沙晨燕,2011)。由于中滩植被密集,有机碳含量显著高于外滩和内滩,为根系和土壤微生物的呼吸提供底物和能量,为产甲烷菌提供了充足分解底物的同时促进了产甲烷作用的发生。
3.3 CO2、CH4来源解析
有机质分解过程伴随着同位素分馏的发生,分解过程中12C会优先反应,富集在产物 CO2、CH4中,而13C则富集在分解后残余的有机质中,因此产物中δ13C偏负值时代表有机质分解程度高,产物中12C相对富集(Zhang et al.,2015)。本研究结果显示,中滩δ13C-CO2高于外滩和内滩,内滩δ13CCO2最低,表明在土壤呼吸释放CO2这一过程中,内滩的有机质分解程度最高,中滩有机质分解程度最低。对于研究区产甲烷过程而言,中滩δ13C-CH4最低,内滩δ13C-CH4最高,中滩有机质分解程度最高,内滩的有机质分解程度最低。同位素分馏系数αc可以衡量湿地土壤中发生的产甲烷方式(张尹等,2020),当αc≤1.055时,产CH4方式以乙酸产甲烷为主,当αc>1.055时,产CH4方式以氢气还原CO2为主(Whiticar et al.,1986)。外滩的αc为 1.040,中滩为1.045,内滩为1.033,3个采样点的产CH4作用均以乙酸产甲烷为主。
在红树林生态系统中,土壤呼吸以活根呼吸和土壤微生物呼吸为主,由于呼吸过程中的低碳分馏(Lin et al.,1997),产生的二氧化碳的δ13C接近其来源的δ13C,根呼吸作用产生的CO2与植物根组织的δ13C具有相同特征,土壤微生物呼吸产生的CO2与土壤有机质的δ13C具有相同特征(Singh et al.,1977;Hanson et al.,2000)。既往研究表明,C3植物的δ13C 值为−35‰— −20‰,平均为−27‰;C4植物的δ13C 值为−15‰— −7‰,平均为−13‰(Andres et al.,1996;Widory et al.,2003)。本研究测定了研究区内主要优势植物叶片和根部的δ13C值为−31.17‰,确定研究区以C3植物为主。此外,分析表明 3个采样点土壤有机碳碳同位素特征相差不大,外滩土壤为−26.44‰,中滩为−26.50‰,内滩为−25.59‰,具备C3植物特征。根据Keeling Plot的拟合结果,外滩的δ13Cs为−29.77‰,接近C3植物碳同位素特征,外滩的土壤CO2主要来源于土壤中C3植物有机碳分解;内滩和中滩的δ13Cs偏大,分别为−16.05‰和−17.73‰,接近C4植物的碳同位素范围。藻种分析表明中滩和内滩土壤表层分布的藻类大部分是绿藻,包括浒苔(Enteromorpha prolifera)和肠浒苔(Enteromorpha intestinalis),少量分布鹧鸪菜(Caloglossa leprieurii),属于红藻。根据先前的研究结果,浒苔属藻类的δ13C为−13.93‰(Irandoost et al.,2021),鹧鸪菜属藻类的δ13C 为−29.85‰(Medina et al.,2022),藻类产生的有机碳的δ13C与藻类组织具有相同的特征,由于藻类向土壤表层增加的有机碳和呼吸作用,导致中滩和内滩的CO2偏大。
4 结论
(1)本研究区红树林土壤的CO2和CH4主要表现为排放特征,CO2和CH4通量存在较明显的时空变化。6—10月的CO2通量显著低于其他月份,7—10月的CH4通量显著高于其他月份,内滩的二氧化碳通量最大,中滩的甲烷排放通量最大。
(2)研究区的CO2通量主要受ORP和土壤含水量的影响,ORP越低,对应了有机物未被完全分解,有足够的底物进行土壤呼吸,土壤含水量降低,促进了有氧呼吸,从而增加了CO2的通量。CH4通量主要受有机碳含量和土壤容重的影响,有机碳含量越高,为根系和土壤微生物的呼吸提供底物和能量,容重越高代表土壤结构越紧实,透气性差,为CH4的产生提供良好的厌氧条件。
(3)在土壤呼吸产生 CO2过程中,内滩δ13CCO2低于外滩和内滩,有机质分解程度最高。而在产甲烷过程中,中滩δ13C-CH4低于外滩和内滩,有机质分解程度最高,这验证了内滩的CO2通量最高和中滩的CH4通量最高。本研究区群落以C3植物为主,研究区内CH4的产生以乙酸产甲烷为主。
猜你喜欢
音乐天地(音乐创作版)(2022年3期)2022-11-05
海洋通报(2022年2期)2022-06-30
农业灾害研究(2022年1期)2022-05-07
儿童故事画报·自然探秘(2022年3期)2022-04-27
幼儿园(2021年4期)2021-07-28
暴雨灾害(2021年2期)2021-04-02
环境卫生工程(2021年1期)2021-03-19
现代装饰(2020年12期)2021-01-18
现代装饰(2019年10期)2019-10-17
歌海(2018年4期)2018-05-14
