
干旱胁迫对红树莓幼苗生长及根系酶活性变化的影响
2022-07-20 09:40:36弓萌萌张瑞禹张培雁张雪梅郭素萍齐国辉
经济林研究 2022年2期
弓萌萌,张瑞禹,刘 洋,2,张培雁,李 寒,张雪梅,2,郭素萍,2,齐国辉,2
(1. 河北农业大学 林学院,河北 保定 071000;2. 河北省核桃工程技术研究中心,河北 邢台 054000)
红树莓Rubus idaeus为蔷薇科悬钩子属多年生落叶半灌木,地下部分为多年生,当年结果后地上部分枯死,俗称托盘、山莓果、悬钩子[1]。其果实酸甜,芳香味浓,营养丰富,被世界粮农组织称为“第3 代水果之王”[2]。除鲜食外,果实还可加工成果酒、果汁、果酱等,深受消费者青睐。红树莓果实除了富含蛋白质、维生素、矿物质等一般果蔬具备的营养成分外,还含有一些特殊的功能性成分,如黄酮类、鞣花酸、超氧化物歧化酶(SOD)、水杨酸等,在抗癌、防衰老和治疗心脑血管栓塞等方面具有一定的医疗保健作用[3-6]。
干旱是影响植物正常生长的主要非生物胁迫之一[7]。不同植物在干旱环境下会有不同的生理特征,以抵抗恶劣环境,维持正常的生命活动[8]。受到干旱胁迫后,植株出现生长速度减慢、叶片失绿畸形萎蔫乃至植株死亡等问题。林姝婧等[9]的研究结果表明,随着干旱胁迫程度的增加,长柄扁桃盆栽苗叶片SOD 活性、过氧化物酶(POD)活性、叶绿素含量、丙二醛(MDA)含量均呈现先升高后降低的趋势。王鑫等[10]的研究结果表明,干旱胁迫下,8 个平欧杂种榛品种的1 年生盆栽苗的高和叶片叶绿素含量均显著下降,MDA含量显著升高。王荔等[11]的研究结果表明,干旱胁迫限制火龙果品种‘紫红龙’盆栽苗茎的伸长和增粗生长,促进根系的萌发和伸长生长。马斌等[12]对武当木兰、红花木莲、乐昌含笑和乐东拟单性木兰4 种木兰科植物进行了干旱胁迫,研究结果表明,随着干旱胁迫程度的增加,4 种木兰科树种的根冠比均呈上升趋势。在生长过程中红树莓幼苗对水分相当敏感,滞水或土壤通气不良,会使植株衰弱,引起病害,破坏根细胞,尤其是水分亏缺时,幼苗叶片发黄、卷曲,植株生长严重受限。在我国北方地区,特别是在西北、华北及春季干旱少雨的地区,春季建园时合理的水分管理及选择抗旱性较强的品种是提高红树莓植株成活率的主要措施。
本试验中研究了持续干旱胁迫条件下3 个红树莓品种幼苗根系酶活性的变化以及地上部分的生长特性,采用隶属函数法对不同红树莓幼苗的抗旱性进行综合评价,以期为红树莓苗期适宜的水分管理及抗旱品种选择提供参考。
1 材料与方法
1.1 试验材料
2018 年6—8 月,在河北农业大学标本园避雨棚中,选择3 个红树莓品种‘海尔特兹’‘秋福’‘托拉蜜’的组培苗进行干旱胁迫试验。苗高约15 cm,栽植在花盆中,每盆4 株。花盆上口径44 cm、下口径25 cm、高30 cm,盆内填充20 kg 基质(园土和沙子质量比为2∶1)。基质pH 为8.72,基质中有机质、全氮、全磷、全钾的质量分数分别为11.62、0.21、0.53、5.35 g/kg,碱解氮、速效磷、速效钾的质量分数分别为56.93、8.34、57.25 mg/kg。
1.2 试验设计
缓苗30 d 后进行持续干旱胁迫处理。干旱胁迫组,于胁迫前倒数第1 天灌透水(以花盆底部有水渗出为标准),之后持续胁迫,分别在胁迫进行了5、10、15、20 和25 d 时取样;复水组,分别在胁迫进行了5、10、15、20 和25 d 时灌透水(以花盆底部有水渗出为标准),复水5 d 后取样;正常管理组(对照),每天18:00 采用称量法进行补水,使土壤含水量控制在田间持水量的60%~80%。每处理20 株,3 次重复。
采集红树莓苗的中部功能叶片置于冰盒,带回实验室清洗,待测;将整株幼苗从盆中取出分离根系,洗净晾干,用液氮快速冷冻并置于-80 ℃冰箱,待测。
1.3 指标测定和分析
使用钢卷尺测量株高生长量;采用比叶重法[13]测定叶面积;采用烘干称量法测定并计算根冠比(地下部分和地上部分干质量的比值);采用丙酮乙醇混合液法[14]测定叶绿素含量;采用硫代巴比妥酸法[15]测定MDA 含量;采用氮蓝四唑(NBT)光还原法[16]测定SOD 活性;采用愈创木酚法[17]测定POD 活性;采用过氧化氢法[16]测定过氧化氢酶(CAT)活性。
1.4 数据处理
采用Duncan 新复极差法进行数据统计与分析。采用隶属函数法对3 个红树莓品种的抗旱性进行综合评价,隶属函数值越大,表明抗旱性越强。

式中:Xi为i指标的测定值;Xmax与Xmin分别为i指标对应的最大值与最小值;U(Xi)和U'(Xi)分别为i指标与抗旱性呈正相关和负相关时的隶属函数值。
2 结果与分析
2.1 干旱胁迫及复水对红树莓地上部分生长的影响
2.1.1 干旱胁迫及复水对株高生长量的影响
干旱胁迫及复水处理下红树莓的株高生长量如图1 所示。由图1 可知,处理期间,正常管理组3 个红树莓品种的株高生长量为3.17 ~4.00 cm。随着干旱程度的加剧,3 个红树莓品种株高生长量不断减少,且下降幅度逐渐增加。胁迫处理15 d 时,‘海尔特兹’和‘秋福’的株高生长量均显著低于CK,分别较CK 降低了37.5%、32.76%,复水处理5 d 后,其株高生长量与CK 和干旱组均无显著差异。胁迫处理20、25 d 时,3 个红树莓品种的株高生长量均极显著低于CK,复水处理5 d 后,株高生长量极显著低于CK,与干旱处理无显著差异。胁迫处理25 d 时‘海尔特兹’‘秋福’‘托拉蜜’的株高生长量最小,分别为1.23、1.17、1.07 cm,分别较CK 降低了66.96%、69.56%、66.32%。
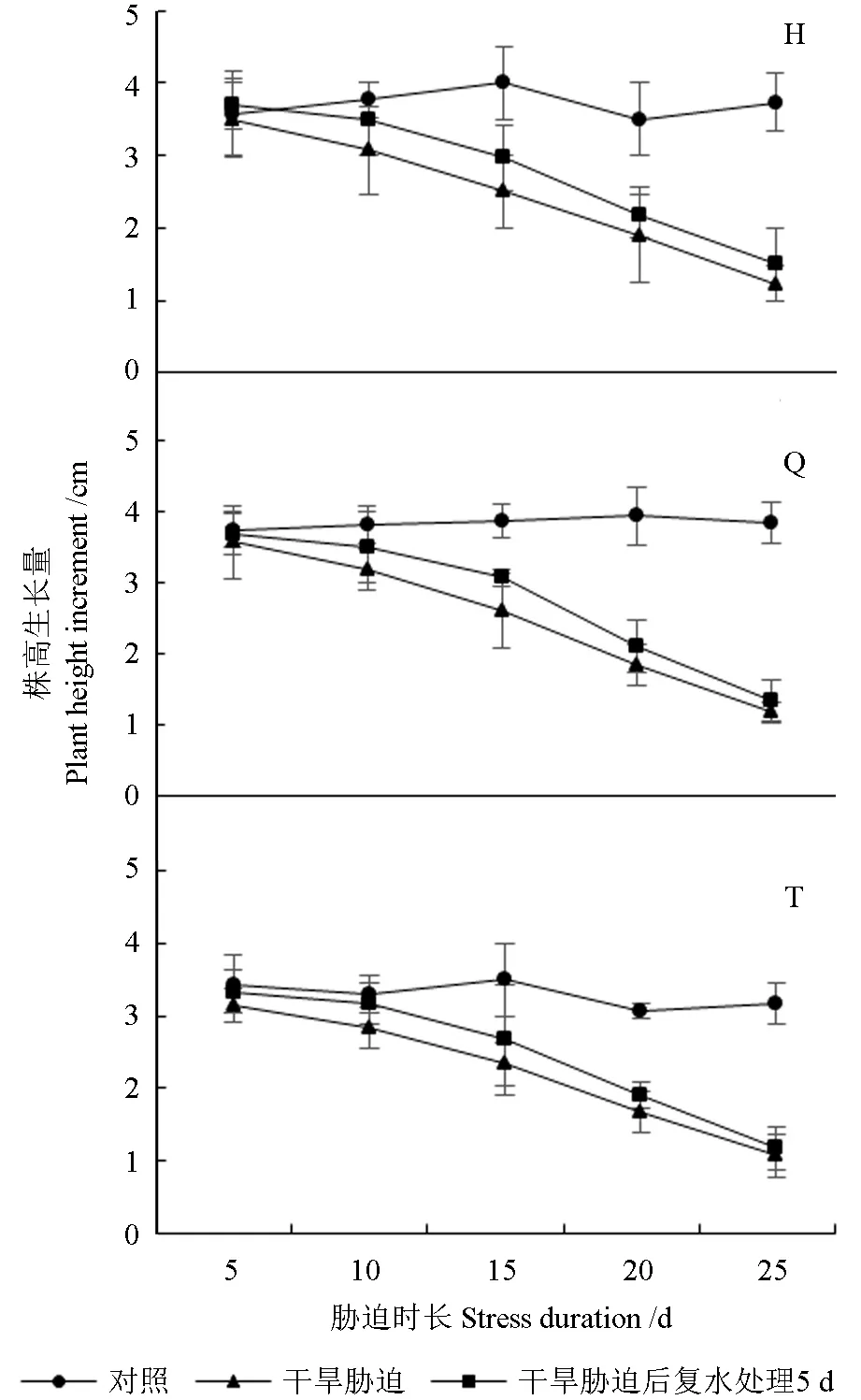
图1 干旱胁迫及复水处理下红树莓的株高生长量Fig. 1 Plant height and growth of red raspberry under drought stress and rehydration treatment
2.1.2 干旱胁迫及复水对单叶面积的影响
干旱胁迫及复水处理下红树莓的单叶面积如图2 所示。由图2 可知,正常管理组3 个红树莓品种的单叶面积均变化不大,且随干旱持续单叶面积均有不断减小的趋势。胁迫处理20 d 时,3 个红树莓品种的单叶面积分别为62.34、60.16、61.14 cm2,均显著低于CK,复水处理5 d 后,单叶面积较干旱处理分别增大了5.76%、6.16%、5.55%,与胁迫组无显著差异,但显著低于CK。胁迫处理25 d 时,3 个红树莓品种的单叶面积均最小,此时‘海尔特兹’‘秋福’‘托拉蜜’的单叶面积分别为55.56、53.91、55.41 cm2,均显著低于CK,复水处理5 d 后,3 个红树莓品种的单叶面积均与干旱处理间无显著差异,但显著低于CK。

图2 干旱胁迫及复水处理下红树莓的单叶面积Fig. 2 Single leaf area of red raspberry under drought stress and rehydration treatment
2.1.3 干旱胁迫及复水对根冠比的影响
干旱胁迫及复水处理下红树莓的根冠比如图3所示。由图3 可知,3 个红树莓品种的根冠比均随干旱持续而增大。胁迫处理15 d 时,‘海尔特兹’和‘秋福’的根冠比分别为0.46、0.42,分别较CK 增加了23.41%、12.76%,均显著高于CK,复水处理5 d 后,根冠比与胁迫处理无显著差异。胁迫处理20、25 d 时,‘海尔特兹’‘秋福’‘托拉蜜’3 个红树莓品种的根冠比均显著高于CK,并在胁迫处理25 d 时达最大值,最大值分别为0.51、0.48、0.46,分 别 较CK 增 加 了31.72%、26.04%、24.57%。胁迫处理25 d 的红树莓经复水处理5 d 后,根冠比较干旱处理分别降低了7.76%、6.31%、5.36%,与干旱处理无显著差异,但均显著高于CK。
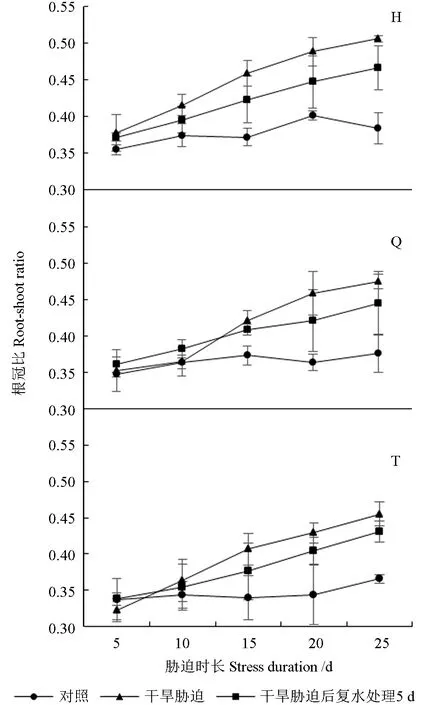
图3 干旱胁迫及复水处理下红树莓的根冠比Fig. 3 Root shoot ratio of red raspberry under drought stress and rehydration treatment
2.1.4 干旱胁迫及复水对叶绿素含量的影响
干旱胁迫及复水处理下红树莓的叶绿素含量如图4 所示。由图4 可知:试验期间,对照组3 个红树莓品种的叶绿素含量较稳定;随着干旱持续,3 个红树莓品种的叶绿素含量先上升后下降。胁迫处理15 d 时,‘海尔特兹’‘秋福’‘托拉蜜’的叶绿素含量均达最大值,分别为2.58、2.53、2.48 mg/g,分别较CK 增加了13.17%、4.44%、5.83%,其中‘海尔特兹’的叶绿素含量显著高于CK。胁迫处理20 d 时,3 个红树莓品种的叶片发黄萎蔫,此时‘秋福’的叶绿素含量为2.30 mg/g,较CK 降低了7.40%,与CK 存在显著差异。胁迫处理25 d 时,3 个红树莓品种的叶片边缘及前端枯萎,‘海尔特兹’‘秋福’‘托拉蜜’的叶绿素含量分别为2.13、2.02、2.00 mg/g,分别较CK降低了15.86%、19.41%、11.22%,与CK 有显著差异,复水处理5 d 后,3 个红树莓品种的叶绿素含量变化不大,显著低于CK。

图4 干旱胁迫及复水处理下红树莓的叶绿素含量Fig. 4 Chlorophyll content of red raspberry under drought stress and rehydration treatment
2.2 干旱胁迫及复水对红树莓根系MAD 含量和抗氧化酶活性的影响
2.2.1 干旱胁迫及复水对根系对MAD 含量的影响
干旱胁迫及复水处理下红树莓根系的MAD 含量如图5 所示。由图5 可知,对照处理下,3 个红树莓品种根系的MAD 含量相对稳定。整个干旱胁迫过程中,3 个红树莓品种根系的MAD 含量均随干旱持续而增加。胁迫处理10 d 时,‘秋福’和‘托拉蜜’根系的MAD 含量显著高于CK,复水处理5 d 后,根系MAD 含量与CK 无显著差异。胁迫处理15、20 d 时,3 个红树莓品种根系的MAD 含量均显著高于CK 和复水处理,复水处理5 d 后,3 个红树莓品种根系的MAD 含量均有不同程度降低,且均显著高于CK。胁迫处理25 d 时,3 个红树莓品种根系的MAD 含量均达最大值,‘海尔特兹’‘秋福’‘托拉蜜’根系的MAD 含量最大值分别为5.88、6.15、6.52 μmol/g,分别较CK 增加了36.04%、41.27%、37.92%,复水处理5 d 后,根系MAD 含量分别为5.53、5.87、6.03 μmol/g,显著高于CK,分别较胁迫处理降低了5.96%、4.59%、7.49%,与胁迫处理间无显著差异。
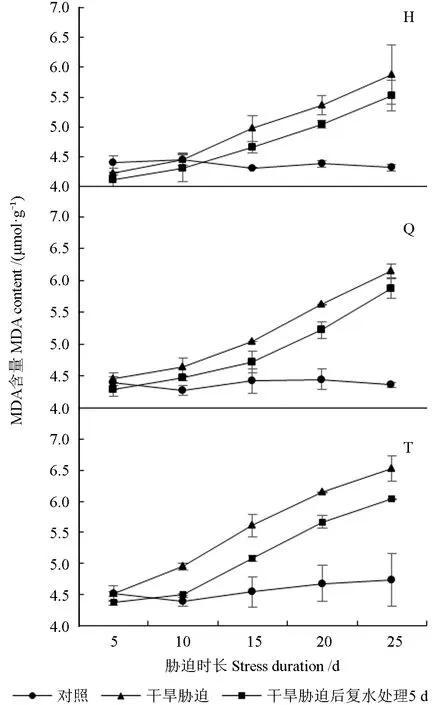
图5 干旱胁迫及复水处理下红树莓根系的MDA含量Fig. 5 MDA content of red raspberry roots under drought stress and rehydration treatment
2.2.2 干旱胁迫及复水对根系SOD 活性的影响
干旱胁迫及复水处理下红树莓根系的SOD 活性如图6 所示。由图6 可知,对照处理下,3 个红树莓品种根系的SOD 活性均相对稳定。胁迫处理过程中,3 个红树莓品种根系的SOD 活性先上升后下降。‘海尔特兹’和‘秋福’根系的SOD 活性均在处理15 d 时达最大值,最大值分别为6.28、6.06 U/(g·min), 分 别 较CK 升 高 了15.34%、10.71%,均极显著高于CK,复水处理5 d 后,‘海尔特兹’根系的SOD 活性为6.10 U/(g·min),较干旱处理降低了2.97%,显著低于干旱处理,‘秋福’根系的SOD 活性为5.93 U/(g·min),较CK 降低了2.16%,与干旱处理无显著差异。‘托拉蜜’根系的SOD 活性在胁迫处理10 d 时达最大值,为5.96 U/(g·min),较CK 升高了8.06%,极显著高于CK,复水处理5 d 后,根系SOD 活性较胁迫处理降低了3.70%,显著低于干旱胁迫处理。

图6 干旱胁迫及复水处理下红树莓根系的SOD活性Fig. 6 SOD activity of red raspberry roots under drought stress and rehydration treatment
2.2.3 干旱胁迫及复水对根系POD 活性的影响
干旱胁迫及复水处理下红树莓根系的POD 活性如图7 所示。由图7 可知,正常管理组3 个红树莓品种根系的POD 活性相对稳定。干旱胁迫条件下,随干旱持续3 个红树莓品种根系的POD活性均呈现先升后降的趋势。胁迫处理10 d 时,3 个红树莓品种根系的POD 活性均显著高于CK,复水处理5 d 后,根系POD 活性与CK 和干旱胁迫处理均无显著差异。胁迫处理15 d 时,‘海尔特兹’根系的POD 活性显著高于CK,‘秋福’和‘托拉蜜’根系的POD 活性极显著高于CK,复水处理5 d 后,‘海尔特兹’根系的POD 活性与CK 和干旱处理均无显差异,‘秋福’和‘托拉蜜’根系的POD 活性显著高于CK,与干旱处理无显著差异。胁迫处理20 d 时,3 个红树莓品种根系的POD 活性均达最大值,最大值分别为114.56、88.75、128.27 U/(g·min),分 别 较CK 增加了1.55、0.99、1.67 倍,复水处理5 d 后,POD活性均有不同程度降低,‘海尔特兹’和‘秋福’根系的POD 活性均极显著高于CK,与干旱处理无显著差异,‘托拉蜜’根系的POD 活性较干旱处理降低了14.41%,显著低于干旱处理,极显著高于CK。胁迫处理25 d 时,3 个红树莓品种根系的POD 活性均急剧下降,复水处理5 d 后,根系POD 活性显著高于CK 和干旱胁迫处理。
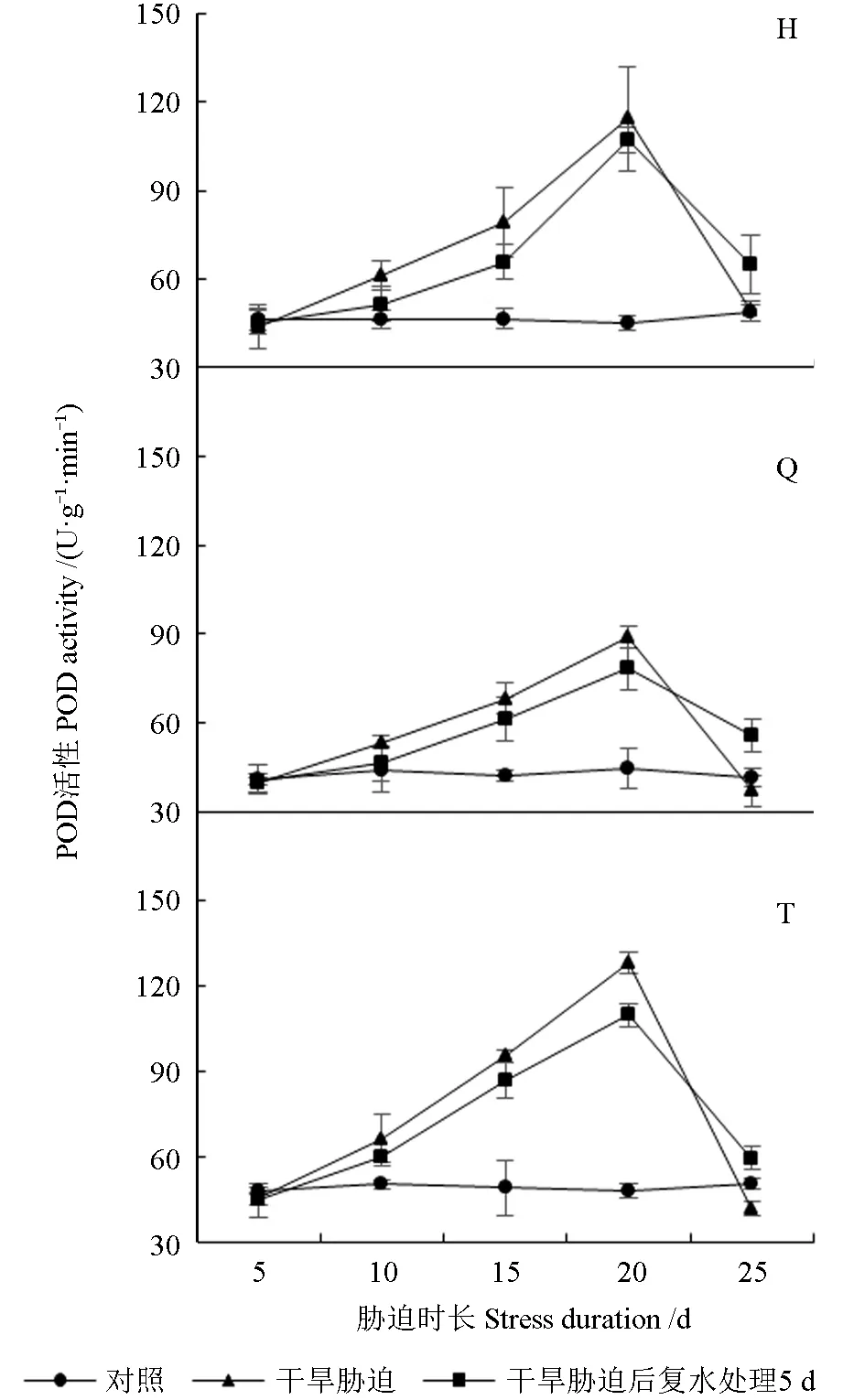
图7 干旱胁迫及复水处理下红树莓根系的POD活性Fig. 7 POD activity of red raspberry roots under drought stress and rehydration treatment
2.2.4 干旱胁迫及复水对根系CAT 活性的影响
干旱胁迫及复水处理下红树莓根系的CAT 活性如图8 所示。由图8 可知,正常管理组3 个红树莓品种根系的CAT 活性相对稳定。随干旱持续,3 个红树莓品种根系的CAT 活性呈现先升高后下降的趋势。胁迫处理15 d 时,‘托拉蜜’根系的CAT 活性达最大值,为5.33 U/(g·min),较CK 上升了130.77%,极显著高于CK,复水处理5 d 后,根系CAT 活性为4.62 U/(g·min),较干旱处理降低了13.33%,与干旱处理无显著差异,极显著高于CK。胁迫处理20 d 时,‘海尔特兹’和‘秋福’根系的CAT 活性最高,最大值分别为6.04、5.69 U/(g·min),分别较CK 升高了142.85%、128.57%,极显著高于CK,复水处理5 d 后,根系CAT 活性分别为5.33、4.98 U/(g·min),分别较干旱处理降低了11.77%、12.50%,与胁迫处理无显著差异,分别较CK 升高了114.28%、100%,显著高于CK。

图8 干旱胁迫及复水处理下红树莓根系的CAT活性Fig. 8 CAT activity of red raspberry roots under drought stress and rehydration treatment
2.3 各红树莓品种抗旱性的综合评价
采用隶属函数法可直观全面地进行品种综合特性评价,该方法被广泛应用于指标评价。3 个红树莓品种幼苗抗旱性的综合评价结果见表1。由表1可知,3 个红树莓品种按照抗旱性由强到弱排列依次为‘海尔特兹’‘秋福’‘托拉蜜’。

表1 干旱胁迫条件下不同红树莓品种各指标的隶属函数值Table 1 Subordinate function values of different indicators of different red raspberry varieties under drought stress
3 结论与讨论
本研究结果表明,干旱胁迫下红树莓幼苗通过抑制生长、调节光合色素合成及抗氧化酶活性协同作用适应干旱环境。胁迫15 d 的3 个红树莓品种经复水处理5 d 后,幼苗生长与正常供水条件下无明显差异,指标可基本恢复至对照水平;胁迫超过15 d 的红树莓复水后,植株叶片变黄、枯萎,生长状况不良,无法恢复至正常状态。3 个红树莓品种按照抗旱性由强到弱排列依次为‘海尔特兹’‘秋福’‘托拉蜜’。
干旱会直接抑制植物的生长,植物通过调整形态构造和各器官的生长速度来适应干旱胁迫[16]。本研究结果表明,随着干旱胁迫持续,3 个红树莓品种的株高生长量和单叶面积均呈现逐渐下降的趋势,这与阿日文[18]、张彦妮等[19]的研究结果一致。王林龙等[20]经研究发现,干旱胁迫增加了不同种源油蒿的根冠比。本研究结果显示,3 个红树莓品种的根冠比均随干旱胁迫持续而增大,这与何静丹等[21]的研究结果相吻合。造成红树莓根冠比增大的原因可能是干旱胁迫下红树莓将较多生长所需养分分配到地下部,从而使得干旱对根系生长的抑制作用小于地上部。在受到干旱胁迫后,红树莓通过加快根系的生长,吸收更多的有机质和水分,同时地上部器官干物质的量逐渐减少,植株生长速度降低,叶面积减少,从而降低地上部水分的消耗。
干旱胁迫下叶绿素的含量不仅与光合能力有关,还与植物的耐旱性有一定关系[22]。干旱胁迫下植物体内叶绿素含量的变化,指示植物对水分胁迫的敏感性,并直接影响光合产量。本研究结果表明,随着干旱胁迫程度的加重,叶绿素含量表现出先增加后降低的变化趋势,这与任迎虹等[23]关于桑树的研究结果一致。胁迫前期,叶绿素含量升高,可能是由于红树莓幼苗在受到短期的干旱胁迫后产生了应激反应,通过增加叶绿素含量来缓解和降低干旱胁迫对红树莓幼苗本身的伤害;胁迫后期,叶绿素含量降低,可能是叶绿素合成受阻或叶绿素降解所致。
植物遭受逆境时,细胞产生大量羟基自由基和超氧阴离子自由基,这些自由基和细胞膜中的不饱和脂肪酸发生膜脂过氧化反应生成MDA[24]。对芍药[25]、板栗[26]的研究结果表明,随着干旱胁迫程度的加剧,MDA 含量呈现逐渐增加的趋势。本研究结果表明,随干旱胁迫持续,根系MDA 含量逐渐增加,这与彭少兵[27]对不同品种树莓、黑莓的研究结果一致。干旱胁迫后复水可以缓解干旱胁迫造成的膜伤害与膜脂过氧化作用的影响,使植物细胞膜脂过氧化产物MDA 积累降低。胁迫5、10、15 d 的红树莓经复水处理5 d 后,根系MDA 含量均与对照无显著差异,胁迫20、25 d 的红树莓经复水处理5 d 后,根系MDA 含量极显著高于对照。这可能是因为短期干旱胁迫引起的清除氧自由基酶类活性的丧失是可逆的,复水后即可恢复,而长期干旱胁迫引发了抗氧化酶的不可逆转失活,即使复水也不能完全激活,使过剩的氧自由基不能及时被清除,从而使根系中有较多的MDA。
在正常生长发育过程中,植物体内活性氧的产生和清除处于平衡,当逆境胁迫来袭时,活性氧自由基增加,其浓度超过植物正常的生理代谢水平,影响植物生长,此时植物体内抗氧化酶活性提高,有效清除活性氧,维持活性氧自由基的动态平衡。左小容等[28]的研究结果显示,在干旱前期SOD、POD、CAT 活性均高于对照,在干旱后期酶活性下降,经过复水后幼苗的抗氧化酶活性与对照处理无明显差异。本研究结果表明,随干旱胁迫的持续,根系中SOD、POD、CAT 活性呈现先升高后降低的变化趋势,这与张希吏[29]的研究结果相吻合。3 个红树莓品种的根系SOD、POD、CAT 活性在不同时期达到最大值,这说明一定时间范围内,干旱胁迫能够提高植物中抗氧化酶的活性,这是植物对外界干旱环境的一种应激反应,但随着胁迫程度的增加,红树莓根系中SOD、POD、CAT 活性均迅速下降,此时干旱程度已经超过了植物对损伤的抵御能力,抗氧化酶活性降低,细胞过氧化作用增强,产生大量自由基。
本研究中未能揭示不同红树莓品种对干旱胁迫及复水处理的分子响应机制。为全面掌握不同红树莓品种的抗旱性及其对复水处理的应答机制,后续将对相关作用的重要差异表达基因、代谢通路等进行检测分析,为使用分子育种方法筛选培育抗旱性强的红树莓品种提供参考。
猜你喜欢
世界最新医学信息文摘(2021年12期)2021-06-09 08:37:06
金桥(2020年8期)2020-05-22 06:22:32
电子制作(2019年11期)2019-07-04 00:34:32
商周刊(2018年24期)2019-01-08 03:30:40
电子制作(2017年17期)2017-12-18 06:40:43
当代医药论丛(2017年22期)2017-04-12 06:29:38
中国酿造(2016年12期)2016-03-01 03:08:19
食管疾病(2015年3期)2015-12-05 01:45:09
中国品牌(2015年11期)2015-12-01 06:21:05
哈尔滨医药(2015年2期)2015-12-01 03:57:34
