
杜撒♂×大长撒♀F1代ARHGEF37基因多态性及其对生长性状的影响
2022-07-14 09:47朱怡轩闵鹏聪陈强王孝义鲁绍雄严达伟李明丽
南方农业学报 2022年4期
朱怡轩 闵鹏聪 陈强 王孝义 鲁绍雄 严达伟 李明丽


摘要:【目的】揭示ARHGEF37基因多態性及其对猪生长性状的影响,为利用该基因标记推进滇撒猪配套系的进一步选育及开发利用提供参考依据。【方法】以274头杜撒♂×大长撒♀ F1代猪为试验材料,采用PCR扩增产物直接测序法检测ARHGEF37基因外显子8多态性,采用最小二乘模型分析各SNP位点基因型及单倍型组合对5个生长性状的影响。【结果】在杜撒♂×大长撒♀ F1代猪ARHGEF37基因外显子8上检测出C1262G、T1293C、G1395A和C1398T 4个SNPs位点,分别以CC、TT、GA、CC基因型、等位基因C、T、G、C及单倍型CTGC的频率最高;除T1293C位点外,其余各位点的杂合度在0.4552~0.4940,遗传多样性较丰富。C1262G位点GG基因型个体达100 kg体重日龄(258.93±2.95 d)显著高于CG基因型个体(249.87±2.68 d)(P<0.05,下同),CG基因型个体30~60 kg日增重(462.44±15.44 g)显著高于CC基因型个体(409.29±14.30 g);T1293C位点TT基因型个体30~60 kg日增重(445.04±9.70 g)极显著高于TC基因型个体(329.33±42.26 g)(P<0.01),TT基因型个体30~100 kg日增重(516.96±6.44 g)显著高于TC基因型和CC基因型个体(470.92±28.08 g和480.64±16.94 g)。此外,达60 kg体重日龄和达100 kg体重日龄均以CTGC/GTGC组合最短(161.84±5.07 d和241.00±5.11 d),CTGC/CCGC组合最长(188.80±11.33 d和272.60±11.42 d);30~60 kg日增重和30~100 kg日增重分别以CTGC/GTAT组合和CTGC/GTGC组合最高(479.56±20.66 g和545.42±19.76 g),均以CTAC/CCGC组合最低(275.00±84.33 g和437.00±55.88 g)。【结论】ARHGEF37基因的G1395A和C1398T位点的遗传多样性较丰富;C1262G位点的CG基因型、T1293C位点的TT基因型及CTGC/GTGC单倍型组合对杜撒♂×大长撒♀F1代生长性状的影响较大,携带有CTGC单倍型个体的各生长性状均处于较高水平。
关键词: 杜撒♂×大长撒♀ F1代猪;ARHGEF37基因;外显子8;单核苷酸多态性;生长性状
中图分类号: S828.89 文献标志码: A 文章编号:2095-1191(2022)04-0918-08
Polymorphism of ARHGEF37 gene and its effect on growth traits in F1 generation pigs of (Duroc×Saba)♂×[Yorkshire×(Landrace×Saba)]♀
ZHU Yi-xuan, MIN Peng-cong, CHEN Qiang, WANG Xiao-yi,
LU Shao-xiong, YAN Da-wei, LI Ming-li*
(College of Animal Science and Technology, Yunnan Agricultural University, Kunming,Yunnan 650201, China)
Abstract:【Objective】To reveal the effect of ARHGEF37 gene polymorphisms on the growth traits of pigs, so as to provide a reference for further breeding, development and the utilization of Diansa pig matching line with this gene marker. 【Method】The study used 274 F1 generation pigs (Duroc×Saba)♂×[Yorkshire×(Landrace×Saba)]♀ (hereafter DS♂×YLS♀) to screen for polymorphisms in the ARHGEF37 gene by the direct sequencing of PCR amplified products. The effects of genotypes and haplotype combinations of each SNP locus on five growth traits were analyzed by the least square model. 【Result】It was found that 4 SNPs loci were located on exon 8 of ARHGEF37 gene: C1262G, T1293C, G1395A and C1398T The frequency of CC,TT,GA and CC genotype was the highest,followed by allele C,T,G,C and haplotype CTGC. Except for the T1293C locus,the remaining showed a heterozygosity between 0.4552-0.4940, indicating more abundant genetic diversity. The age to 100 kg body weight in the C1262G GG genotype (258.93±2.95 d) was significantly higher than that in the CG genotype (249.87±2.68 d)(P<0.05,the same below). Measurements of the average daily weight gain in pigs of 30-60 kg showed that the CG genotype (462.44±15.44 g) was significantly higher than that of the CC genotype (409.29±14.30 g) and the TT genotype (445.04±9.70 g) at T1293C was extremely significantly higher than that of the TC genotype (329.33±42.26 g)(P<0.01). The average daily weight gain in pigs between 30-100 kg in the TT genotype (516.96±6.44 g) was significantly higher than that of the TC and CC genotypes (470.92±28.08 g and 480.64±16.94 g, respectively). In addition, the time to achieve 60 and 100 kg body weight was the shortest in the CTGC/GTGC combination(161.84±5.07 d and 241.00±5.11 d) and the longest in the CTGC/CCGC combination(188.80±11.33 d and 272.60±11.42 d). The average daily gains between 30-60 kg and between 30-100 kg were the highest in the CTGC/GTAT and CTGC/GTGC combinations (479.56±20.66 g and 545.42±19.76 g, respectively), and the lowest in the CTAC/CCGC combination (275.00±84.33 g and 437.00±55.88 g). 【Conclusion】The genetic diversity of G1395A and C1398T of ARHGEF37 gene is more abundant. The CG genotype of the C1262G locus, TT genotype of the T1293C locus and the combination of CTGC/GTGC haplotype have more influence on the growth traits of the F1 generation of DS♂×YLS♀ pigs. The growth traits of individuals carrying the CTGC haplotype are all at a relatively high level.
Key words: F1 generation of DS♂×YLS♀ pigs; ARHGEF37 gene; exon 8; single nucleotide polymorphism;growth trait
Foundation items:National Key Research and Development Program(2018YFD0501206); Yunnan Industrial Technology Leading Talents Special Project(YNWR-CYJS-2018-056); Yunnan Science and Technology Planning Project (202104BI090022); Yunnan Major Science and Technology Special Project(202102 AE090039)
0 引言
【研究意义】滇撒猪配套系是我国第一个以纯地方猪种(撒坝猪)选育、以专门化品系为基础育成的猪配套系,母系父本品系和终端父本品系分别以长白猪和大约克为育种素材,历时13年培育成功(袁跃云等,2014)。该配套系充分聚合了撒坝猪肉质优良、耐粗饲及引进品种生长速度快、瘦肉率高等特性,在区域养猪生产中已得到广泛应用(连林生等,2006)。经过持续系统选育的撒坝猪专门化母系有效保持了撒坝猪原有的优点,各项性能得以明显提升,与国外猪种杂交的杂种优势较突出,但目前关于滇撒猪配套系主要经济性状的分子遗传机制尚不清楚。因此,进一步对相关候选基因,尤其是基于基因组学发掘的新候选基因开展针对性研究,深入揭示滇撒猪配套系主要经济性状的分子遗传机制,对促进该配套系的选育提高具有重要意义。【前人研究进展】Rho鸟嘌呤核苷酸交换因子37(Rho guanine nucleotide exchange factor 37,ARHGEF37)基因是在弥漫性B细胞淋巴瘤(Diffuse B lymphoma)中发现的一种长度为45 kb的DNA序列,命名为DbI(Rossman et al.,2005)。鸟嘌呤核苷酸交换因子(ARHGEF)主要包括两大类,即DbI家族ARHGEF与非DbI ARHGEF,其中关于DbI家族ARHGEF的研究较多。研究表明,ARHGEF参与生物胚胎发育、Rho GTPase活性调节及神经元的形成等过程,对哺乳动物的细胞功能也有重要作用。ARHGEF可影响生物的胚胎发育,与生物体内多个器官的發育及分化均有关系。研究人员将斑马鱼的遮蔽蛋白A(Obscurin A)结构域中与ARHGEF相互作用的区域去除后,斑马鱼的胚胎发育出现明显受阻,其骨骼肌、心脏及大脑均出现发育迟缓或不完全现象(Raeker et al.,2010)。ARHGEF还在细胞生长分化过程中发挥多种重要作用,如细胞骨架重组、细胞内微管形成、细胞黏附作用及细胞极性等。ARHGEF对Rho GTPase活性具有调节作用。Rho GTPase在调节细胞骨架形成、基因转录、细胞周期及膜泡运输中起重要作用,与肿瘤及神经性疾病的发生也有密切关系(刘明涛等,2014),且在转录调控和细胞转化中发挥重要作用(丁美娟等,2020)。ARHGEF是一类通过调节Rho GTP酶的GDP/GTP交换,从而调节Rho GTPase活性的调节蛋白。在细胞的信号转导过程中,ARHGEF起着信号转换器或分子开关的作用,通过作用于细胞骨架或靶蛋白而产生多种生物效应(Van Aelst and D’Souza-Schorey,1997;O'Connor and Mercurio,2001;Wheeler and Ridley,2004)。正常情况下,Dbl家族成员处于自我抑制状态,一般在出现N末端或C末端截断突变后被激活,通过将Rho-GDP转变为Rho-GTP而激活Rho家族以控制细胞的生长发育和浸润转移等(唐秋琳和毕锋,2017)。ARHGEF作为Rho GTPase家族的上游信号调节蛋白,通过微管依赖的激活机制和非微管依赖的激活机制进行活化,从而发挥多种生物学功能。ARHGEF还可影响神经元的形成。已有研究在灵长类前额叶皮层中发现ARHGEF亚细胞的分布能明显影响神经元的激活(Meiri et al.,2012),并认为这种影响机制很可能是因为ARHGEF的表达调节微管与肌动蛋白之间存在的耦合作用,由于ARHGEF的不平衡分布将会影响细胞骨架的形成,神经细胞轴突和树突的伸展也因此会受到较大影响。【本研究切入点】部分基于全基因组的研究结果及本课题组前期开展简化基因组测序结果提示,ARHGEF37基因可能是影响猪生长性状的潜在候选基因,但目前尚无关于ARHGEF37基因多态性及其对猪经济性状影响的相关报道。因此,有必要开展相关研究,揭示并证实ARHGEF37基因的遗传效应。【拟解决的关键问题】将ARHGEF37基因作为杜撒♂×大长撒♀ F1代猪生长性状相关的候选基因,通过分析ARHGEF37基因外显子8区域的SNP变异位点与5个生长性状间的关联程度,以期探索此基因对杜撒♂×大长撒♀ F1代猪群生长性状的影响,为利用ARHGEF37基因标记推进滇撒猪配套系的进一步选育及开发利用提供参考依据。
1 材料与方法
1. 1 试验样品
试验样品来自云南省楚雄州种猪种鸡场,选择生长发育正常、健康的以杜洛克♂×撒坝猪♀(杜撒)公猪为父本、大白猪♂×(长白猪♂×撒坝猪♀)母猪为母本的杜撒♂×大长撒♀ F1代猪274头(其中母猪138头、公猪136头)。将采集的猪耳组织泡入装有75%酒精的EP管中,置于-80 ℃超低温冰箱中保存备用。
1. 2 生长性能测定
猪体重达30 kg左右开始测定其生长性能指标,至110 kg左右时结束。分别测定达60 kg体重日龄、达100 kg体重日龄、100 kg活体背膘厚、30~60 kg日增重和30~100 kg日增重共5个生长性能指标,均按照NY/T 822—2019《种猪生产性能测定规程》执行。
1. 3 基因多态性检测
1. 3. 1 DNA提取与检测 采用北京擎科(昆明)生物技术有限公司的DNA提取试剂盒(TSINGKE)提取基因组DNA,再通过1%琼脂糖凝胶电泳和超微量检测仪(NanoDrop 2000)完成待测DNA样品的定性和定量分析。检测合格的DNA条带清晰明亮,无明显拖带,符合扩增要求,置于-20 ℃冰箱中保存备用。
1. 3. 2 引物合成和PCR扩增 通过Ensembl数据库(http://asia.ensembl.org/index.html)获取猪ARHGEF37基因外显子8序列,利用Primer Premier 5.0设计1对引物,序列为F:5'-TTTGACGACCACCCGATAC-3';R:5'-CTGCTGCGTTTGAGTCCC-3',预扩增片段大小857 bp。PCR反应体系25.0 μL:2×Taq PCR Mix 12.5 μL,正、反向引物各0.5 μL,DNA模板1.0 μL,灭菌水10.5 μL。扩增程序:94 ℃预变性5 min;94 ℃ 30 s,62 ℃ 30 s,72 ℃ 30 s,进行35个循环;72 ℃延伸7 min。
1. 3. 3 测序与基因分型 检测合格的PCR产物经直接测序后,使用BioEdit v7.0.9完成序列比对,再结合其序列峰图找出SNP位点,各SNP位点的位置及其基因型由CDS序列和相应的碱基名称命名。
1. 4 统计分析
根据ARHGEF37基因分型结果,分别统计试验猪群中不同SNP位点相应基因型的个数,并进行群体遗传学分析,再使用Phase 2.0对ARHGEF37基因各SNP位点完成单倍型分析,最后采用最小二乘模型对杜撒♂×大长撒♀ F1代猪的生长性状进行关联分析:
Yij=μ+Gi+Sj+eij
式中,Yij为性状观察值;μ为群体均值;Gi为该SNP位点的第i基因型效应或单倍型组合效应;Sj为第i性别效应,eij为随机误差,假定服从N(0,σ[2e])分布。
统计分析由SAS 9.0的GLM过程完成(鲁绍雄和林连生,2003),结果以最小二乘均数±标准误的形式表示,且进行差异显著性检验,多重比较在α=0.05和α=0.01水平上进行。
2 结果与分析
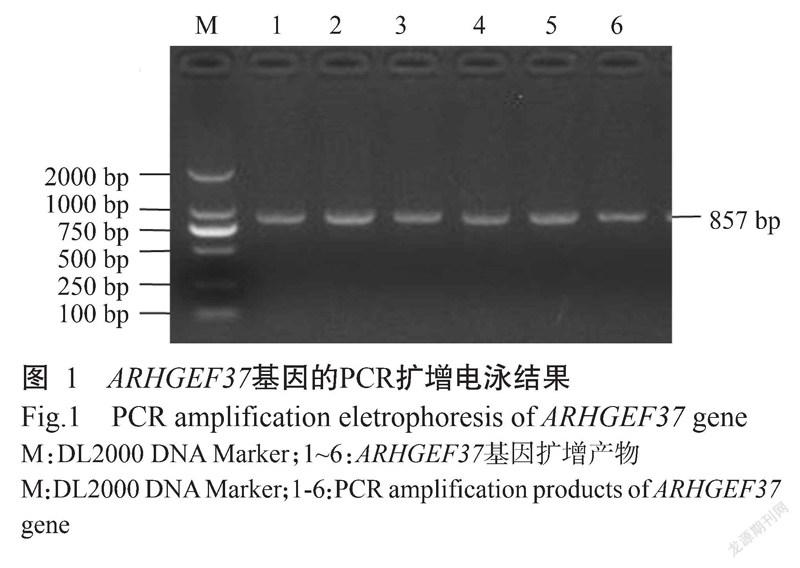
2. 1 目的片段扩增与SNP位点检测结果
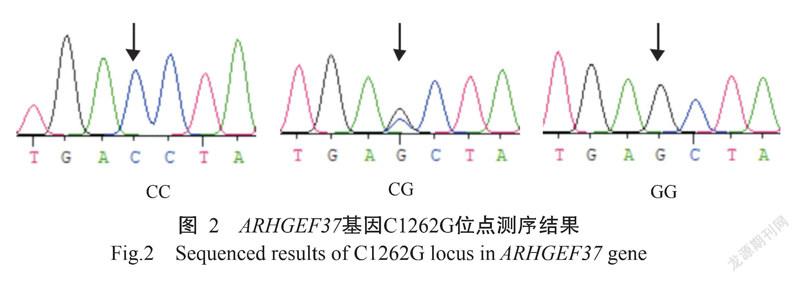
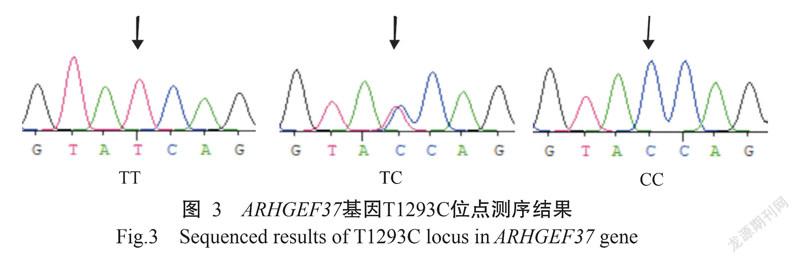
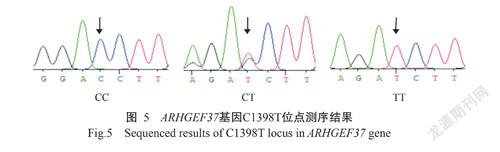
ARHGEF37基因的PCR扩增电泳结果(图1)显示,扩增条带单一、大小为857 bp,与预期结果一致,满足DNA测序要求。PCR产物测序在ARHGEF37基因第8外显子区域发现4个SNPs位点:即CDS序列1262 bp处C→G(C1262G)为错义突变,导致ARHGEF37蛋白第421位的氨基酸由苏氨酸(Thr)突变为丝氨酸(Ser);1293 bp处T→C(T1293C)、1395 bp处G→A(G1395A)及1398 bp处C→T(C1398T),均为同义突变,分别对应编码络氨酸(Tyr)、甘氨酸(Gly)和天冬氨酸(Asp)。4个SNPs位点均检测出3种基因型,即C1262G位点存在CC、CG和GG基因型(图2),T1293C位点TT、TC和CC基因型(图3),G1395A位点GG、GA和AA基因型(图4),C1398T位点存在CC、CT和TT基因型(图5)。
2. 2 ARHGEF37基因多态性分析结果
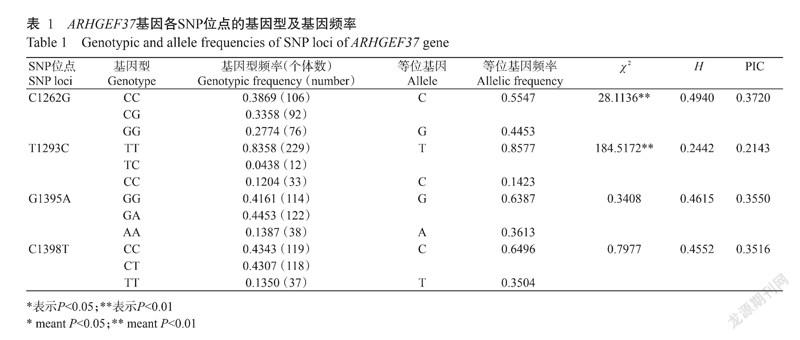
杜撒×大长撒F1代ARHGEF37基因C1262G、T1293C、G1395A和C1398T位点的等位基因频率和基因型频率见表1。由表1可知,C1262G、G1395A和C1398T位点均为中度多态,多态信息含量(PIC)分别为0.3720、0.3550和0.3516。其中,G1395A和C1398T位点遗传多样性较丰富,杂合度(H)分別为0.4615和0.4552,且达Hardy-Weinberg平衡状态(P>0.05),C1262G位点未达Hardy-Weinberg平衡状态(P<0.01)。T1293C位点为低度多态(PIC为0.2143),未达Hardy-Weinberg平衡状态(P<0.01)。此外,等位基因频率显示,C、T、G、C分别是对应位点的优势等位基因。
2. 3 ARHGEF37基因多态位点的单倍型分析结果
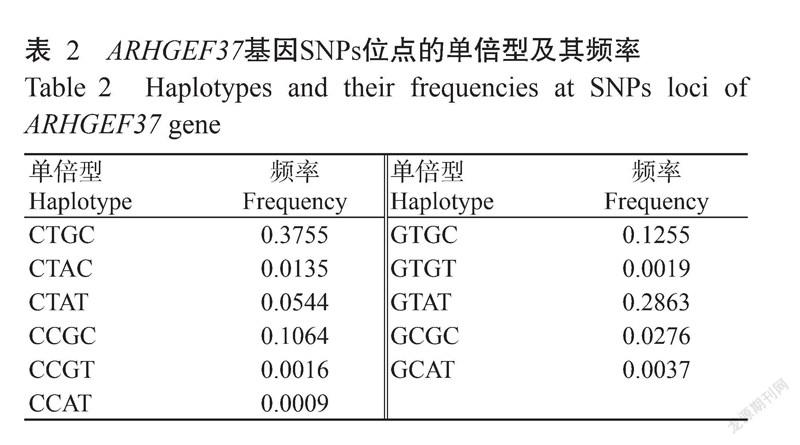
通过Phase 2.0进行单倍型分析,结果(表2和表3)显示,C1262G、T1293C、G1395A和C1398T 4个SNPs位点共计11种单倍型,20种单倍型组合。单倍型以CTGC的频率最高(0.3755),GTAT的频率次之(0.2863),CCAT的频率最低(0.0009)(表2)。单倍型组合则以CTGC/CTGC组合的频率最高(0.1861),CTGC/GTAT的频率次之(0.1825),10种单倍型组合CTAC/GCGC、GCAT/GCAT、GTGC/GTGT、CTAT/CCGT、CTAC/GTAT、CCGC/CCAT、CTGC/CTAC、CTAT/CCGC、CCGC/GTGC和CTAC/CTAC均只检测出1个个体(表3)。
2. 4 ARHGEF37基因各SNP位点不同基因型的生长性状差异
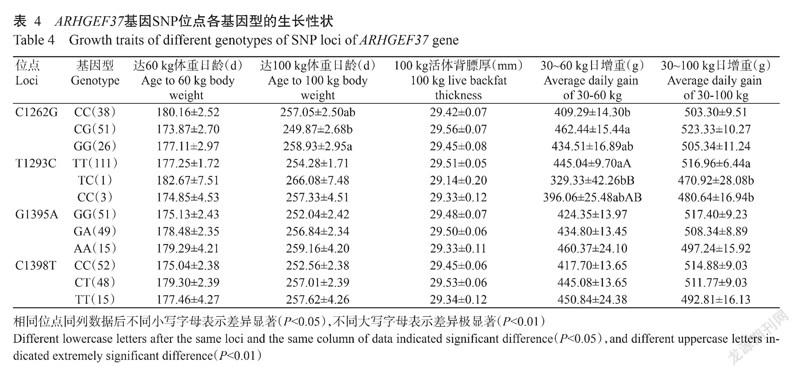
杜撒×大长撒F1代ARHGEF37基因C1262G、T1293C、G1395A和C1398T位点各基因型的生长性状见表4。由表4可知,C1262G位点GG基因型个体达100 kg体重日龄(258.93±2.95 d)最长,显著高于CG基因型个体(P<0.05,下同),CG基因型个体达100 kg体重日龄(249.87±2.68 d)最短,且CG基因型个体30~60 kg日增重(462.44±15.44 g)最高,显著高于CC基因型个体(409.29±14.30 g);T1293C位点TT基因型个体30~60 kg日增重(445.04±9.70 g)最高,极显著高于TC基因型个体(329.33±42.26 g)(P<0.01,下同),TT基因型个体30~100 kg日增重(516.96±6.44 g)最高,显著高于TC基因型和CC基因型个体(470.92±28.08 g和480.64±16.94 g)。
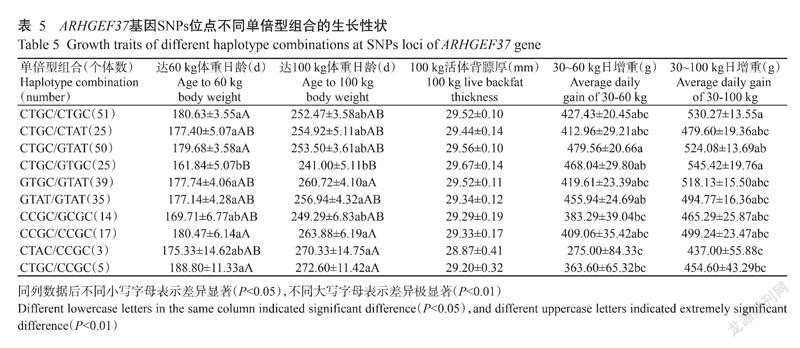
2. 5 ARHGEF37基因不同单倍型组合的生长性状差异
杜撒×大长撒F1代ARHGEF37基因C1262G、T1293C、G1395A和C1398T位点各单倍型组合的生长性状见表5。由表5可知,除100 kg活体背标厚外,4个SNPs突变位点的不同单倍型组合在其他生长性状上均表现出显著或极显著差异。其中,达60 kg体重日龄和达100 kg体重日龄均以CTGC/GTGC组合最短(161.84±5.07 d和241.00±5.11 d),CTGC/CCGC组合最长(188.80±11.33 d和272.60±11.42 d);30~60 kg日增重和30~100 kg日增重分别以CTGC/GTAT组合和CTGC/GTGC组合最高(479.56±20.66 g和545.42±19.76 g),均以CTAC/CCGC组合最低(275.00±84.33 g和437.00±55.88 g)。
3 讨论
DNA分子标记技术的飞速发展和应用,为人们从分子水平揭示包括猪在内的畜禽主要经济性状的遗传基础提供了有力工具,而候选基因法已成为检测猪经济性状相关基因或分子标记的重要途径之一。目前有关滇撒猪配套系生长性状的候选基因研究主要集中在其母系母本撒坝猪专门化品系上。鲁绍雄等(2007)研究表明,肌细胞生成素(Myogenin,MYOG)基因Msp I酶切位点多态性对撒坝猪4月龄体重和45日龄~6月龄日增重有显著影响,且均以AB基因型个体最高,AA基因型个体最低。李国治等(2008)研究报道,心脏脂肪酸结合蛋白(H-FABP)基因的5'上游区1125~1817区段的Hinf I酶切位点不同基因型对撒坝猪生长性状无显著影响。丁艳等(2012)的研究结果显示,胰岛素样生长因子1(Insulin-like growth factor 1,IGF-1)基因Hha I酶切位点AB基因型为撒坝猪120日龄体重、180日龄体重和45~180日龄阶段日增重的增效基因型。钱锦花和连林生(2014)研究表明,撒坝猪SLA-DQA基因EcoR I酶切位点对0~45日龄日增重和70日龄~4月龄日增重有显著影响,其中AB基因型个体的0~45日龄日增重最大,70日龄~4月龄日增重则以BB基因型个体最高。
ARHGEF对哺乳动物的细胞功能有显著影响,如在细胞骨架、细胞屏障功能、细胞分裂、细胞周期、树突棘形态及癌细胞的增殖、凋亡和侵袭转移等过程中发挥重要作用(王莉和李晓东,2015)。猪的ARHGEF37基因位于2号染色体上,共有12个外显子和11个内含子。Fernández等(2012)利用PorcineSNP60 BeadChip芯片对利比利亚猪×长白猪杂交的F3代群体进行全基因组连锁分析,发现猪的2号染色体112~117 cM区域存在显著影响生长和胴体组成的QTL。基于课题组前期试验筛选,ARHGEF37基因第8外显子的遗传变异较丰富,具有一定代表性。本研究通过PCR产物直接测序,在杜撒×大长撒F1代ARHGEF37基因第8外显子上发现4个SNPs突变位点:C1262G、T1293C、G1395A和C1398T,且均检测出3种基因型,分别以CC、TT、GA、CC基因型及C、T、G、C等位基因的频率最高。由χ2检验可知,C1262G和T1293C位点未达Hardy-Weinberg平衡状态,而G1395A和C1398T位点达Hardy-Weinberg平衡状态,可能是选种选育过程中人工选择所致(李金泽等,2008)。此外,根据Botstein等(1980)提出的标准,C1262G、G1395A点和C1398T位点均为中度多态(0.25≤PIC≤0.50),T1293C位点为低度多态(PIC<0.25)。G1395A和C1398T位点杂合度较丰富(H为0.4615和0.4552),且遗传多样性也较丰富,说明该基因的选择潜力较大,可作为有效遗传标记应用于开展其与经济性状的关联分析。
不同位点各基因型及其单倍型组合的关联分析结果表明,ARHGEF37基因的4个SNP突变位点对杜撒×大长撒F1代的部分生长性状有不同程度的影响:C1262G位点以杂合型CG型相对于突变型GG型显著缩短达100 kg体重日龄,杂合型CG型相对于野生型CC型显著提高30~60 kg日增重;T1293C位点以野生型TT型相对于杂合型TC型显著提高30~60 kg日增重,野生型TT型相对于杂合型TC型和突变型CC型显著提高30~100 kg日增重。除100 kg活体背膘厚外,4个SNPs突变位点的不同单倍型组合在其他生长性状上均表现出显著或极显著差异,其中,达60 kg体重日龄和达100 kg体重日龄均以CTGC/GTGC单倍型组合最短,30~60 kg日增重及30~100 kg日增重分别以CTGC/GTAT组合、CTGC/GTGC组合最高。可见,CTGC/GTGC组合对杜撒×大长撒F1代生长速度的影响最大。同时,携带有CTGC单倍型个体的各生长性状也均处于较高水平,说明CTGC单倍型具有提高杜撒×大长撒F1代猪群生长速度的效果,4个SNPs位点对生长性状的影响表现出明显的协同作用。究其原因可能是某些情况下,单个突变位点的作用较小,而多个位点联合可能会对目标性状产生显著影响效果(闫霞,2014)。
综上所述,可推断ARHGEF37基因对杜撒×大长撒F1代猪的生长性状具有显著影响,鉴于当前有关于猪ARHGEF37基因的研究报道十分匮乏,若能综合考虑该基因相邻区域的其他基因或SNP分子标记,并在不同品种的猪群中加以研究确认,将有利于更深入地揭示ARHGEF37基因的真实效应,对育种实际具有重要指导意义。
4 结论
ARHGEF37基因G1395A和C1398T位点的遗传多样性较丰富;C1262G位点的CG基因型、T1293C位点的TT基因型及CTGC/GTGC單倍型组合对杜撒×大长撒F1代猪生长性状的影响较大,携带有CTGC单倍型个体的各生长性状均处于较高水平。
參考文献:
丁美娟,刘艺丹,孔庆霞. 2020. Rho鸟嘌呤核苷酸交换因子15在神经系统疾病中的研究进展[J]. 临床神经病学杂志,33(1):66-68. [Ding M J,Liu Y D,Kong Q X. 2020. Research progress of Arhgef15 in neurological disorders[J]. Journal of Clinical Neurology,33(1):66-68.] doi:10. 3969/j.issn.1004-1648.2020.01.017.
丁艳,鲁绍雄,连林生. 2012. 撒坝猪胰岛素样生长因子-I基因多态性及其与部分生长性状的关联分析[J]. 中国畜牧兽医,39(10):181-184. [Ding Y,Lu S X,Lian L S. 2012. The polymorphism of IGF-I gene and its relationship with some growth traits in Saba pig[J]. China Animal Husbandry & Veterinary Medicine,39(10):181-184.] doi:10.3969/j.issn.1671-7236.2012.10.042.
李国治,连林生,鲁绍雄,严达伟,苟潇,赵桂英. 2008. 云南撒坝猪H-FABP基因多态性及与部分生产性能的关系[J]. 家畜生态学报,29(6):13-16. [Li G Z,Lian L S,Lu S X,Yan D W,Gou X,Zhao G Y. 2008. The relationship between polymorphisms of H-FABP gene and production traits in Yunnan Saba pigs[J]. Acta Ecologiae Animalis Domastici,29(6):13-16.] doi:10.3969/j.issn.1673-1182. 2008.06.003.
李金泽,曹桂荣,岳敏,张建明,刘运忠,顾为望. 2008. 西藏小型猪H-FABP基因的PCR-RFLP研究[J]. 中国实验动物学报,16(3):201-205. [Li J Z,Cao G R,Yue M,Zhang J M,Liu Y Z,Gu W W. 2008. Polymorphism of heart fatty acid-binding protein gene in Tibet minipig populations detected by PCR-RFLP[J]. Acta Laboratorium Animalis Scientia Sinica,16(3):201-205.] doi:10.3969/j.issn.1005- 4847.2008.03.010.
连林生,鲁绍雄,严达伟,连槿,李明丽,苟潇,王开荣,赵中保,任文辉,陈文芳. 2006. 滇撒猪配套系选育[C]//中国畜牧兽医学会. 中国畜牧兽医学会2006学术年会论文集(上册). 北京:中国农业出版社. [Lian L S,Lu S X,Yan D W,Lian J,Li M L,Gou X,Wang K R,Zhao Z B,Ren W H,Chen W F. 2006. Breeding of Dian Sa pig matching line[C]. Chinese Animal Husbandry and Veterinary Association. Proceedings of 2006 Annual Conference of Chinese Animal Husbandry and Veterinary Association(Volume I). Beijing:China Agriculture Press.]
刘明涛,杨军,王志钢. 2014. Rho鸟嘌呤核苷酸交换因子家族及其在疾病发生中的作用[J]. 生物技术通报,34(1):63-67. [Liu M T,Yang J,Wang Z G. 2014. RhoGEF family and the role in occurrence and development of diseases[J]. Biotechnology Bulletin,34(1):63-67.] doi:10. 13560/j.cnki.biotech.bull.1985.2014.01.032.
鲁绍雄,丁艳,李明丽,连林生,严达伟,李国治. 2007. 撒坝猪MYOG基因Msp I多态性及其与部分生长性状的关系[J]. 云南农业大学学报,22(3):385-388. [Lu S X,Ding Y,Li M L,Lian L S,Yan D W,Li G Z. 2007. The MspI polymorphism of MYOG gene and its relationship with some growth traits in Saba pig[J]. Journal of Yunnan Agricultural University,22(3):385-388.] doi:10.3969/j.issn. 1004-390X.2007.03.017.
鲁绍雄,连林生. 2003. SAS统计分析系统在畜牧科学中的应用[M]. 昆明:云南科技出版社. [Lu S X,Lian L S. 2003. Application of SAS statistical analysis system in animal husbandry science[M]. Kunming:Yunnan Science and Technology Press.]
钱锦花,连林生. 2014. 撒坝猪SLA-DQA基因多态性与生长性状的相关性分析[J]. 畜牧与兽医,46(5):59-61. [Qian J H,Lian L S. 2014. The polymorphism of SLA-DQA gene and its associations with growth traits in Saba pig[J]. Ani-mal Husbandry & Veterinary Medicine,46(5):59-61.]
唐秋琳,毕锋. 2017. 鸟苷酸交换因子ARHGEF10功能鉴定及其与肿瘤恶性表型间的关系研究[J]. 实用肿瘤学杂志,31(6):481-488. [Tang Q L,Bi F. 2017. Identification of guanine nucleotide exchange factor ARHGEF 10 and its relationship with tumor malignant phenotype[J]. Practical Oncology Journal,31(6):481-488.] doi:10.11904/j.issn. 1002-3070.2017.06.001.
王莉,李晓东. 2015. GEF-H1的生物学作用及其与肿瘤的关系[J]. 解剖科学进展,21(1):75-79. [Wang L,Li X D. 2015. The biological functions of GEF-H1 and it’s crucial determinant of tumor pathogenesis[J]. Progress of Anatomical Sciences,21 (1):75-79.]
闫霞. 2014. 猪CCK和CCKAR基因多态性及其与生长性状的关联研究[D]. 昆明:云南农业大学. [Yan X. 2014. Single nucleotide polymorphisms(SNP)of CCK and CCKAR gene and their associations with growth traits in swine[D]. Kunming:Yunnan Agricultural University.]
袁跃云,孙利民,叶绍辉. 2014. 云南省畜禽遗传资源志[M]. 昆明:云南科技出版社. [Yuan Y Y,Sun L M,Ye S H. 2014. Genetic resources of livestock and poultry in Yunnan Province[M]. Kunming:Yunnan Science and Technology Press.]
Botstein D,White R L,Skolnick M,Davis R W. 1980. Construction of a genetic linkage map in man using restriction fragment length polymorphisms[J]. American Joural of Human Genetics,32(3):314-331. doi:10.1016/0165-1161(81)90274-0.
Fernández A I,Pérez-Montarelo D,Barragán C,Ramayo-Caldas Y,Ibáñez-Escriche N,Castelló A,Noguera J L,Silió L,Folch J M,Rodríguez M C. 2012. Genome-wide linkage analysis of QTL for growth and body composition emplo-ying the PorcineSNP60 BeadChip[J]. BMC Genetics,13(1):41. doi:10.1186/1471-2156-13-41.
Meiri D,Marshall C B,Greeve M A,Kim B,Balan M,Suarez F,Bakal C,Wu C J,Larose J,Fine N,Ikura M,Rottapel R. 2012. Mechanistic insight into the microtubule and actin cytoskeleton coupling through dynein-dependent RhoGEF inhibition[J]. Molecular Cell,45(5):642-655. doi:10.1016/j.molcel.2012.01.027.
O'Connor K L,Mercurio A M. 2001. Protein kinase A regulates Rac and is required for the growth factor-stimulated migration of carcinoma cells[J]. Journal of Biological Chemistry,276(51):47895-47900. doi:10.1074/jbc.M107 235200.
Raeker M O,Bieniek A N,Ryan A S,Tsai H J,Zahn K M,Russell M W. 2010. Targeted deletion of the zebrafish obscurin A RhoGEF domain affects heart,skeletal muscle and brain development[J]. Developmental Biology,337(2):432-443. doi:10.1016/j.ydbio.2009.11.018.
Rossman K L,Der C J,Sondek J. 2005. GEF means go:Turning on RHO GTPases with guanine nucleotide-exchange factors[J]. Nature Reviews Molecular Cell Biology,6(2):167-180. doi:10.1038/nrm1587.
Van Aelst L,D’Souza-Schorey C. 1997. Rho GTPases and signaling networks[J]. Genes & Development,11(18):2295-2322. doi:10.1101/gad.11.18.2295.
Wheeler A P,Ridley A J. 2004. Why three Rho proteins? RhoA,RhoB,RhoC,and cell motility[J]. Experimental Cell Research,301(1):43-49. doi:10.1016/j.yexcr.2004. 08.012.
收稿日期:2021-07-21
基金項目:国家重点研发计划项目(2018YFD0501206);云南省产业技术领军人才专项(YNWR-CYJS-2018-056);云南省科技计划项目(202104BI090022);云南省重大科技专项(202102AE090039)
通讯作者:李明丽(1969-),https://orcid.org/0000-0002-3073-4035,教授,主要从事动物生产与环境卫生研究工作,E-mail:xiaolucao@126.com
第一作者:朱怡轩(1996-),https://orcid.org/0000-0001-6932-276X,研究方向为动物生产,E-mail:1418247662@qq.com
