山羊STEAP1 基因和EIF2B4 基因多态性与繁殖性能的关联分析
2022-07-13 11:22:00范业锴刘玉芳江炎庭欧阳依娜张伟峰洪琼花储明星
中国畜牧杂志 2022年7期
范业锴,刘玉芳,陶 林,江炎庭,欧阳依娜,张伟峰,洪琼花*,储明星*
(1.中国农业科学院北京畜牧兽医研究所,农业农村部动物遗传育种与繁殖重点实验室,北京 100193;2.河北工程大学生命科学与食品工程学院,河北邯郸 056001;3.云南省畜牧兽医科学院,云南昆明 650224)
过去几千年,人类通过对野生动物的驯养和驯化逐步积累了育种经验。山羊是人类最早驯化的野生动物之一,也是一种重要的经济动物,其分布遍及全世界。山羊按其经济用途可分为乳用、肉用、绒用3 类。繁殖性能是肉用山羊重要的经济性状,但产羔数、初生窝重和断奶窝重等繁殖性状因为遗传力低、分子机制尚不明确使常规育种方法进程缓慢,难以在短时间内快速改良。分子标记辅助育种具有准确度高、操作性强的优点,可以缩短育种年限,加快育种进程,提高育种效率。因此,繁殖相关基因候选分子标记的发掘具有重要意义。
人前列腺6-跨膜上皮抗原()家族由和组成,它 们具有作为金属离子还原酶的天然活性,在金属离子的代谢过程中发挥重要作用。它们参与了广泛的生物过程,如胞内和胞外途径中的分子运输以及细胞增殖和凋亡。1999 年,Hubert 等人发现前列腺六次跨膜上皮抗原l(Six-Transmembrane Epithelial Antigen of the Prostate 1,)在前列腺癌中该基因高表达,并指出在其他癌症细胞系中存在差异表达。后续研究证明,不仅在前列腺恶性肿瘤中异常高度表达,而且在肺癌、尤文氏肉瘤、结肠癌、胃癌、乳腺癌中亦存在高度表达。焦志等研究证明在人的卵巢癌细胞中高度表达。因此,基因可能在卵巢细胞异常增殖过程起重要作用。
真核生物起始因子2B(EIF2B)是蛋白质合成所必需的一种GTP 交换因子,由5 个不同的亚基组成(EIF2B1、EIF2B2、EIF2B3、EIF2B4 和EIF2B5),是EIF2B 的第4 个亚单位,在RNA 到蛋白质翻译过程中发挥重要功能。基因可以促进GDP 和GTP 的相互转换,该基因主要作用于少突胶质细胞和星形胶质细胞,促进细胞内翻译过程。因此,基因可能在调控山羊繁殖性能的主效基因翻译过程中起重要作用。
课题组前期对云上黑山羊(以努比山羊为父本、云岭山羊为母本培育而成的我国第一个肉用黑山羊品种)重测序数据表明,云上黑山羊群体和基因存在多态性位点,因此,推测和基因可能是影响山羊繁殖性能的重要候选基因。本实验通过对和基因重测序结果的分析,发现了基因突变位点g.72016288 T>G,基因g.45853158 C>T、g.45857371 T>A、g.458 57371 T>A,采用MassARRAYSNP 分型技术对和的基因型进行检测,分析候选基因多态性位点与山羊繁殖性状的关联性。本研究通过分析和基因在云上黑山羊、济宁青山羊和辽宁绒山羊群体中的遗传多态性及其与繁殖性能的关联性,以期为通过分子标记辅助选择选育多羔山羊新品种提供参考。
1 材料与方法
1.1 样品采集和表型记录 本研究共采集了768 只母羊(2~5 岁,544 只云上黑山羊、133 只济宁青山羊和91只辽宁绒山羊)的颈静脉血液(10 mL/只),用酚-氯仿法提取基因组DNA。云上黑山羊来源于云南红河哈尼族彝族自治州,拥有至少1 胎产羔数记录,部分拥有产羔窝重和断奶窝重(3 月龄)记录。济宁青山羊来源于农业农村部济宁青山羊保种基地(山东省嘉祥县),辽宁绒山羊来源于辽宁绒山羊育种中心(辽宁省辽阳市)。济宁青山羊和辽宁绒山羊没有具体产羔数记录。本研究将济宁青山羊视为高繁殖力群体,将辽宁绒山羊视为低繁殖力群体。
1.2 基因分型 利用MassARRAY技术对和基因候选多态性位点(g.45853158 C>T、g.45857371 T>A、g.45857371 T>A 和g.72016288 T>G)进行分型。分型引物信息见表1。

表1 引物序列
1.3 统计分析 群体遗传学参数的计算方法参考Zhang等。计算产羔数和窝重的平均数后,基因型的效应采用单因素方差分析。多重比较采用Tukey 法。云上黑山羊低产和高产基因型差异显著性检验、Hardy-Weinberg 平衡检验、候选基因多态性位点基因型与品种(高繁品种:济宁青山羊,低繁品种:辽宁绒山羊)之间的关系检验、产羔数和窝重的基因型效应分析等数据统计分析由SPSS(V.20.0)完成,产羔数和窝重用平均值±标准误表示。
1.4 山羊和基因的生物信息学分析 山羊的基因序列和翻译蛋白序列来自NCBI(登录号:XM_005679303.3,XP_005679360.3)。山羊的基因来自NCBI(登录号:XM_018055503.1,XP_017910992.1)。利用Param tool(https://web.expasy.org/protparam/)用于预测目的蛋白理化性质;利用Prot Scale 在线工具(https:∥web.expasy.org/protscale/)对其目的蛋白的疏水性进行分析;采用NopoPro 在线工具(https://www.novopro.cn/tools/signalp)进行目的蛋白信号肽预测;采用Detaibio 在线工具(http://www.deta ibio.com/tools/index.php?r=transmembrane%2Findex)进行目的蛋白跨膜区预测;SCRATCH Protein Predictor(http://scratch.proteomics.ics.uci.edu/index.html)用于预测目的蛋白的二硫键;NetOGlyc 4.0 Server(http://www.cbs.dtu.dk/services/NetOGlyc/)用于预测目的蛋白的糖基化;NetPhos 3.1 Server(http://www.cbs.dtu.dk/services/NetPhos/)用于预测目的蛋白的磷酸化位点。采用SOPMA 在线工具(https://npsa-prabi.ibcp.fr/cgibin/ npsa_automat.pl?page=npsa_sopma.html)进行目的蛋白二级结构分析。
2 结果
2.1和基因多态性分析 基因分型结果表明,4 个候选位点均存在多态性(图1)。群体遗传学分析表明,基因g.45857345 C>T 位点在云上黑山羊、济宁青山羊、辽宁绒山羊群体中均为中度多态(0.25
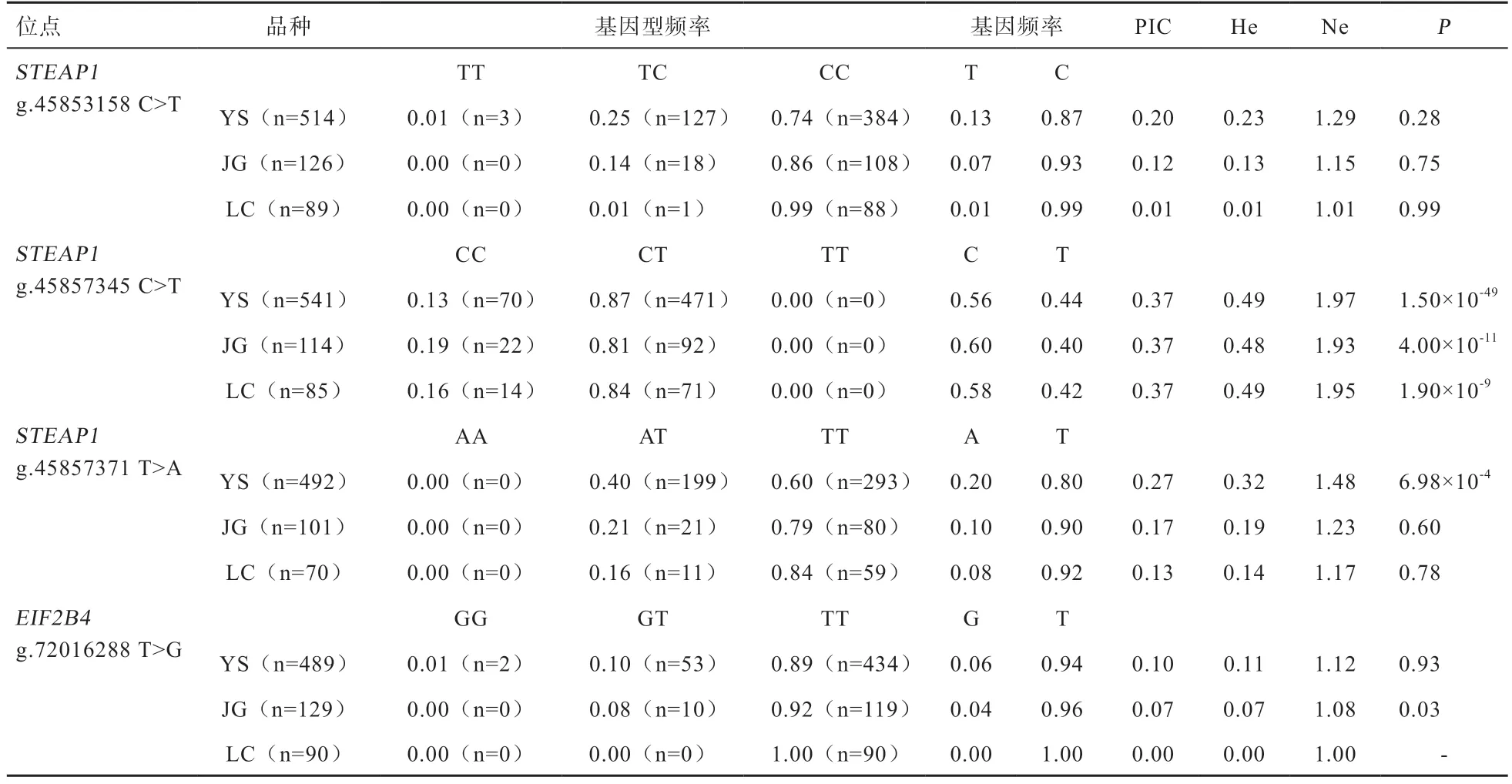
表2 STEAP1、EIF2B4 基因4 个多态位点在不同山羊品种中的群体遗传学分析
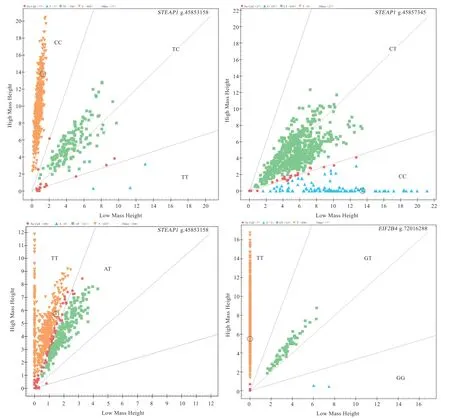
图1 基因分型结果
2.2 候选多态性位点与繁殖性能的关联分析 如表3 所示,g.45853158 C>T 和g.72016288 T>G 在不同繁殖力品种(济宁青山羊和辽宁绒山羊)中的基因型分布存在极显著差异。g.45857345 C>T、g.45857371 T>A 在不同繁殖力品种(济宁青山羊和辽宁绒山羊)中的基因型分布差异不显著。
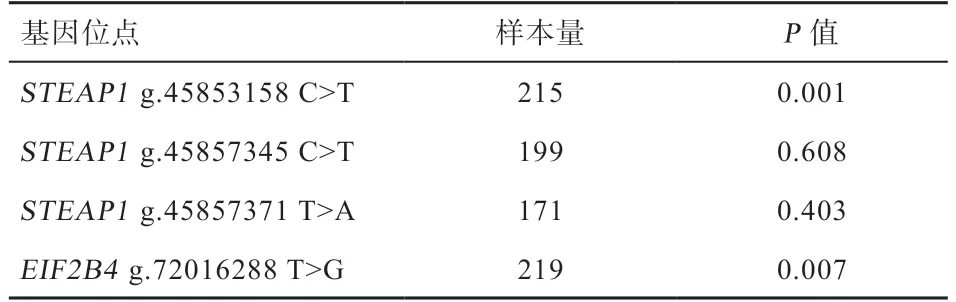
表3 候选位点基因型与不同繁殖力品种的卡方检验
由表4 可知,云上黑山羊高产群体和低产群体中所有候选多态性位点的基因型频率和基因频率均无显著差异。

表4 STEAP1、EIF2B4 基因多态位点在云上黑山羊高产、低产中的基因型频率和等位基因频率
表5 中,所有基因候选多态性位点均对产羔数无显著影响,所有基因候选多态性位点对初生窝重和断奶窝重无显著影响。
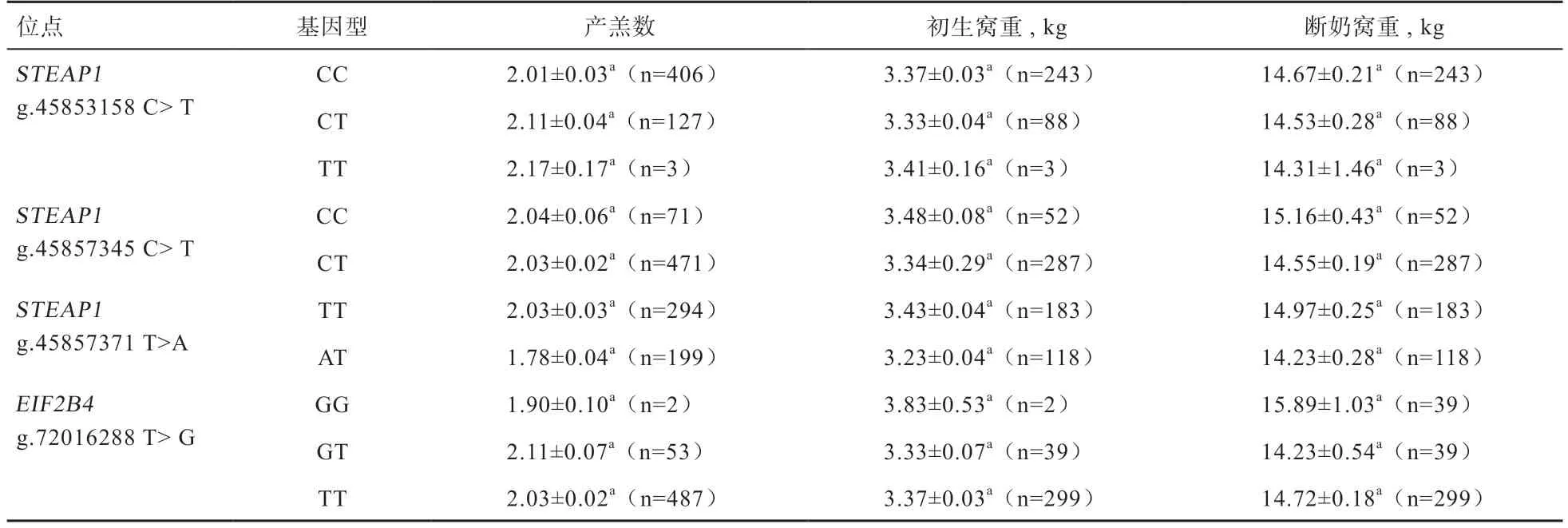
表5 STEAP1 和EIF2B4 对云上黑山羊繁殖性能的影响(均值±标准误)
2.4 STEAP1 和EIF2B4 蛋白的生物信息学分析STEAP1 蛋白的理化性质预测表明:该蛋白由339 个氨基酸组成,其分子量为40.03 KDa,等电点为9.928 2,分子式为CHNOS,总原子数为5 479;从蛋白组成来看,其主要由20 种氨基酸组成,其中亮氨酸(Leu)含量最高为13.9%,而半胱氨酸(Cys)含量最低为0.3%,蛋白在动物体内的半衰期为30 h,不稳定指数为51.60,脂溶性指数为114.13,亲水平均值为0.103,故该蛋白是一种不稳定脂溶性的疏水蛋白;从信号肽和跨膜区预测结果显示,该蛋白属无信号肽结构(图2A),存在6 个跨膜区(图2B);二硫键预测结果显示,该蛋白存在3 个半胱氨酸(Cys),蛋白序列没有形成二硫键。
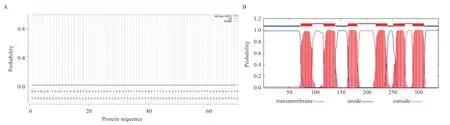
图2 山羊STEAP1 蛋白的信号肽(A)和跨膜结构域(B)预测
EIF2B4 蛋白的理化性质预测表明,该蛋白由539个氨基酸组成,其分子量为58.89 KDa,等电点为9.41,分子式为CHNOS,总原子数为8 357;从蛋白组成来看,其主要由20 种氨基酸组成,其中亮氨酸(Leu)含量最高为11.1%,而色氨酸(Trp)含量最低为0.4%,蛋白在动物体内的半衰期为30 h,不稳定指数为48.44,脂溶性指数为87.05,亲水平均值为-0.378,故该蛋白是一种不稳定脂溶性的亲水蛋白;从信号肽和跨膜区预测结果显示,该蛋白属无信号肽结构(图3A)、无跨膜区(图3B);二硫键预测结果显示,该蛋白存在11 个半胱氨酸(Cys),蛋白序列没有形成二硫键。
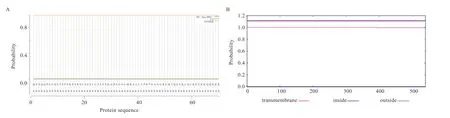
图3 山羊EIF2B4 蛋白的信号肽(A)和跨膜结构域(B)预测
基因的3 个多态性位点均为错义突变,基因的多态位点属于同义突变。基因多态位点对应蛋白变化为g.45853158 C>T 使295 位Val>Lle,g.45857345 C>T 使49 位Arg>His,g.45857371 T>A 使40 位Gln>His,3 个多态位点中只有g.45853158 C>T 符合Hardy-Weinberg 平衡且在不同繁殖力品种羊中差异极显著,为探究g.45853158 C>T 基因突变前后蛋白功能和结构的变化,对其突变前后糖基化位点、磷酸化位点、蛋白质二级结构变化进行了预测,预测结果发现:突变前后STEAP1 编码蛋白的糖基化和磷酸化位点未发生变化,突变前(图4A)和突变后(图4B)STEAP1 编码蛋白的二级结构中突变后-转角区域减少,-螺旋区域增多,延伸链区域减少,无规卷曲区域减少。
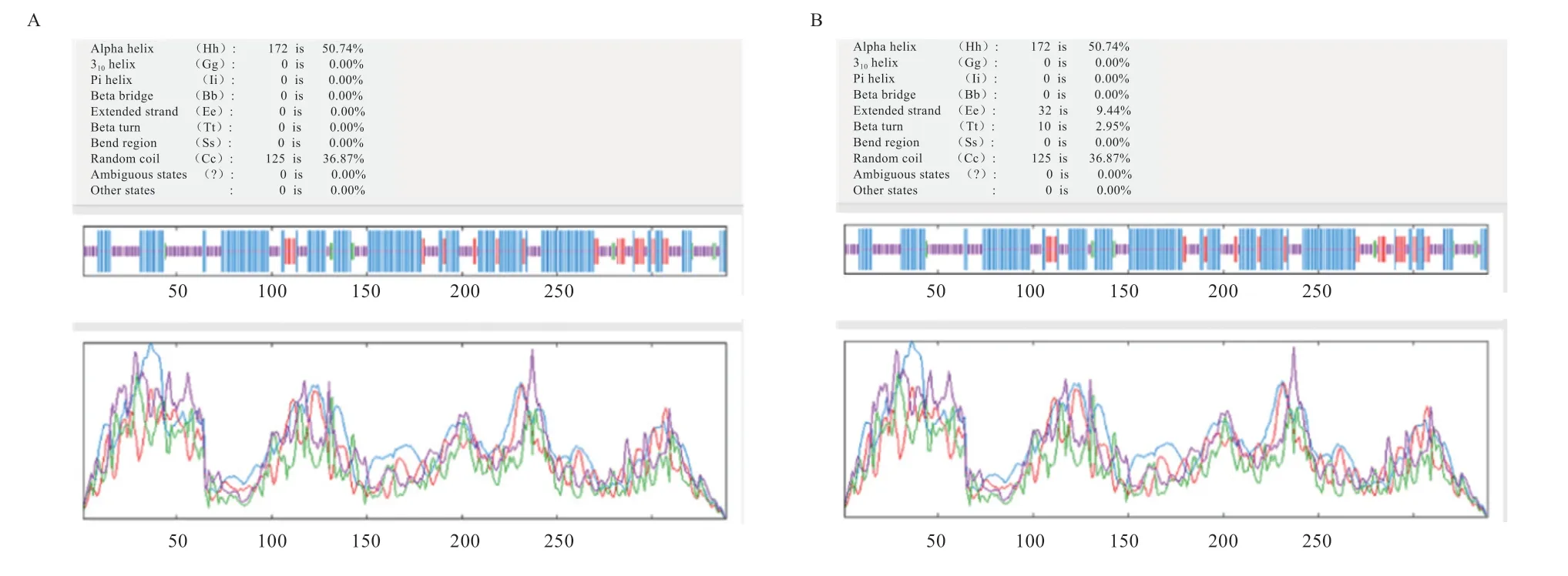
图4 山羊STEAP1 突变前编码蛋白二级结构(A)和突变后编码蛋白二级结构(B)
3 讨论
山羊的产羔性能受微效多基因控制,遗传机理尚未明确。基因多态性指同一物种的不同基因组DNA等位序列之间的差异,多态性位点可能造成mRNA 转录水平和蛋白质翻译水平发生改变,进而影响生物性状。前人研究发现,基因在人卵巢中高度表达,推测该基因可能对山羊的繁殖性能有着重要影响,本研究对课题组前期重测序数据进行整合分析发现,基因在山羊中存在g.45853158 C>T、g.45857371 T>A 和g.45857371 T>A 3 个多态位点,且均为错义突变,对该基因多态性位点和山羊繁殖性能进行关联分析,发现基因g.45853158 C>T 突变在不同繁殖性能群体中有显著差异。李晓琴等研究发现,mRNA 到蛋白质的翻译速率与蛋白质二级结构之间有一定关系:高翻译速率时编码螺旋、无规卷曲较多;低翻译速率时编码无规卷曲较多、编码螺旋较少;折叠结构则随翻译速率表现出明显的振荡。同时,密码子的使用在不同片段内一般也是不均匀的:在螺旋片段内,结构尾部偏向使用高翻译速率密码子;中部倾向使用中翻译速率密码子;而头部使用的密码子翻译速率偏低。
对山羊基因突变前后蛋白理化性质和空间结构分析后发现,g.45853158 C>T 使山羊基因编码蛋白空间二级结构-转角区域减少,-螺旋区域增多,延伸链区域减少,无规卷曲区域减少,推测其为影响基因g.45853158 C>T 在不同繁殖力群体有显著差异的重要因素。
前人研究发现,基因是真核细胞翻译起始因子,对人体所有细胞中启动蛋白质合成过程中RNA 到蛋白质的翻译有着重要作用且基因可以促进GDP 和GTP 的相互转换,推测该基因可作为繁殖相关细胞的翻译起始因子;对重测序数据进行整合分析发现,基因在山羊中存在g.72016288 T>G,为同义突变;对位点和对应繁殖性能进行整合分析后发现,g.72016288 T>G 突变对山羊繁殖性能无显著影响。
4 结论
基因的位点g.45853158 C>T 在不同繁殖性能群体中有显著差异,突变造成STEAP1 编码蛋白二级空间结构发生变化,可作为云上黑山羊产羔性能的分子标记,但不适合进行窝选。
猜你喜欢
今日农业(2022年14期)2022-09-15 01:43:28
云南畜牧兽医(2022年4期)2022-08-19 01:56:34
云南畜牧兽医(2021年6期)2021-12-24 02:14:02
系统工程学报(2021年4期)2021-12-21 06:21:08
云南畜牧兽医(2021年5期)2021-10-27 07:27:50
新世纪智能(高一语文)(2020年12期)2020-06-01 08:14:20
新农业(2016年23期)2016-08-16 03:38:12
新农业(2016年22期)2016-08-16 03:34:42
兽医导刊(2016年6期)2016-05-17 03:50:32
中国医药导报(2015年27期)2015-02-28 22:08:01