
CO2浓度升高条件下不同氮素水平对小叶章光合特性和生长的影响
2022-07-10 23:04:00王建波王继丰付晓玲钟海秀刘赢男倪红伟
中国农学通报 2022年17期
王建波,王继丰,付晓玲,钟海秀,刘赢男,倪红伟
(1黑龙江省科学院自然与生态研究所,哈尔滨 150040;2黑龙江省林业科学院,哈尔滨 150081)
0 引言
全球气候变化已经被科学家所证实,大气CO2浓度升高是当前全球气候变化的一个主要因素[1]。自工业革命以来,由于化石燃料燃烧等人类活动,大气CO2浓度已经增加了40%以上[2],预计到21世纪末将达到700µmol CO2/mol[3]。CO2是植物光合作用的底物,其浓度升高必将影响到植物的光合作用和生长发育等多个过程。一般认为,CO2浓度升高使植物叶片的光合速率升高。但对于不同的物种,在不同的生态环境条件下,增加的幅度不同。CO2浓度升高的初始阶段将使植物的光合速率显著增加,而后随着时间的推移,其增加速率会逐渐减弱或渐渐消失,有的甚至出现“光合下调”现象[4],也称为“光合适应”[5],这与植物N素的供应有着密切关系。例如,在高大气CO2浓度下,植物在土壤氮素供应较高时不发生光合适应现象,这一方面可能与增施氮素调节了植物体内C/N关系有关[6-7],另一方面也与施氮可能增加了Rubisco等光合酶的含量和活性有关[8]。
氮是植物生长所需要的大量营养元素之一,在大多天然陆地生态系统中,植物的生长发育都受到氮素营养的限制。而全球N沉降也由1961年的14 Tg N/a增加到2000年的68 Tg N/a,预计到2050年全球N沉降量将达到200 Tg N/a[9]。以往对大气CO2浓度升高和氮素营养的研究多集中在栽培植物,且多集中在农田[10-12]、森林[13-14]等生态系统,对于湿地生态系统的研究较少[15-16]。由于天然群落的复杂性,要真正了解不同氮素沉降程度下,CO2浓度升高对于植物所产生的影响以及植物对其作出的响应,有必要在野外自然条件下进行监测和研究。
三江平原是中国面积最大、分布最为集中的湿地区,是具有重要代表性和国际意义的湿地生态系统[17]。已有研究指出氮素是湿地系统中最主要的限制性养分。目前,由于农业生产活动过程中流失的氮素随地表径流、大气沉降等多种途径进入湿地中[18]。为明确高大气CO2浓度下湿地植物光合特性对不同氮量的响应,本研究在野外原位条件下,以三江平原优势植物小叶章为研究对象,采用开顶式气室进行不同CO2浓度处理,施用不同量的NH4NO3,研究小叶章气体交换参数随CO2升高处理时间延长的变化;不同施氮量如何调控CO2浓度升高对小叶章光合生理特性和生长的影响;探讨长期高大气CO2浓度下小叶章叶片的光合参数及其氮素调控机制,这一工作以期为未来湿地植物应对大气CO2浓度升高提供理论依据。
1 材料与方法
1.1 研究地点
本研究地区位于中国东北三江平原——洪河国家自然保护区,黑龙江省科学院自然与生态研究所三江平原湿地生态定位研究站内(47°49′N、133°40′E)。三江平原是由黑龙江、松花江和乌苏里江冲击而成,地势平坦,地面坡降小,土质粘重,渗透困难,发育有淡水沼泽、沼泽化草甸、河流、湖泊等湿地类型。近年来由于气候变化和大规模的农业活动,区域降水量减少、地下水位下降,湿地退化严重。小叶章(Calamagrostis angustifolia)是三江平原典型草甸、沼泽化草甸的建群植物和优势植物,沼泽植被中的优势种、亚优势种或重要的伴生种,常伴生有毛果苔草(Carex lasiocarpa)、芦苇(Phragmitescommunis)、狭叶甜茅(Glyceria spiculosa)、灰脉苔草 (Carex appendiculata)、二歧银莲花(Anemone vivularis)、五 脉 山 黧 豆 (Lathyrus quinqunervuius)等植物。
1.2 试验方法
采用开顶式气室(open top chamber,OTC)模拟大气CO2浓度升高。OTC气室为正八边形棱柱状,框架高1.8 m,顶45°角,边长1.5 m,框架之间为高透光钢化玻璃构成。充气系统包括气瓶(液态CO2,纯度为99.9%)、流量计、鼓风机和PVC管。PVC管(内径为7.5 cm)围绕箱体一周,其高度随植物生长可进行调整,PVC管朝箱体中央一侧每隔1 cm开直径为0.1 cm的小孔。气瓶排出的CO2在鼓风机作用下通过PVC管上的小孔均匀分布在开顶箱内,通过流量计调节流量,使箱体内形成相对均一的CO2浓度。在整个生长季保持连续通气,每天7:00—18:00充气,雨天停止充气。试验设置3个处理:高浓度CO2(700±20 μmol/mol,E2)、中浓度 CO2(550 ±20 μmol/mol,E1)和 背 景 CO2(380±20 μmol/mol,E0)。设置 3 个氮素供应水平 N0[0 g/(m2·a)]、N1[4 g/(m2·a)]和 N2[8 g/(m2·a),每个处理3次重复。因此,试验共计9个处理。根据天气情况分别于CO2浓度处理0、25、52、72、86、112天测定气体交换参数,于处理72天取样测定叶片可溶性糖、淀粉、可溶性蛋白含量,在处理86天测定小叶章株高等。
1.3 测定指标
1.3.1 光合参数测定 在每株小叶章上部选取完全伸展的、具有相似生理生态特征的叶片,用Li-6400便携式光合系统 (Li-Cor,Inc,美国)于晴天9:00—11:00测定叶片净光合速率(Pn)、气孔导度(Gs)、胞间CO2浓度(Ci)和蒸腾速率(Tr)。测定时采用CO2小钢瓶控制叶片叶室 CO2浓度,E0、E1和 E2气室测定时叶室 CO2浓度分别设置为380、550、700 μmol/mol。测定时利用内置红蓝光源,光量子通量密度设置为1200 μmol/(m2·s),叶室内温度设25℃。
1.3.2 可溶性糖、淀粉、可溶性蛋白、氮素含量测定 于处理72天取样测定叶片可溶性糖、淀粉、可溶性蛋白、氮素含量。选取中上部完全展开的功能叶作为测定材料;采用蒽酮比色法测定可溶性糖和淀粉含量;采用考马斯亮蓝染色法测定可溶性蛋白含量;采用微量凯式定氮法测定氮素含量。
1.4 数据分析
采用Sigmaplot 10.0进行数据处理和图表绘制,用SPSS 16.0统计软件进行方差分析和差异显著性分析。
2 结果分析
2.1 不同处理对小叶章气体交换参数的影响
由图1可知,不同处理小叶章净光合速率随着时间进展均表现出先增加后降低的趋势。小叶章光合速率在72天达到最大值,而后各处理小叶章净光合速率逐渐下降。N0和N1条件下的小叶章光合速率在处理25天之前表现为E1和E2显著高于E0处理,说明短期CO2浓度升高可以提高小叶章光合作用;此后E1、E2和E0两者差距逐渐缩小,72天之后E1和E2条件下小叶章净光合速率显著低于E0;而N2条件下,E1和E2小叶章净光合速率在72天后较E0条件下略微升高。

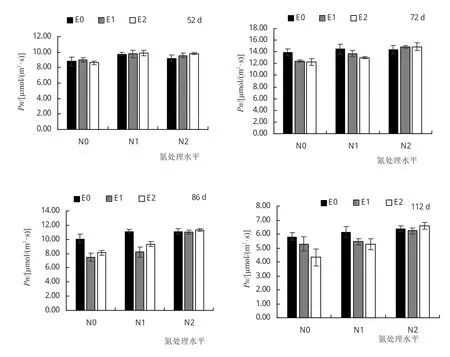
图1 不同处理对小叶章净光合速率的影响
具体而言,N0条件下,与 E0处理相比,E1和 E2处理的净光合速率分别下降11%和12%(P<0.05);在N1条件下,E1和E2处理的净光合速率分别下降5%(P>0.05)和10%(P<0.05);在N2条件下,E1和E2处理净光合速率有稍增加的趋势,但变化不显著(P>0.05,表1)。说明小叶章在CO2熏蒸一段时间后出现光合下调现象,而施加一定浓度的氮对小叶章气体交换有一定的促进作用。在3个CO2浓度下,施氮均增加了小叶章叶片净光合速率。
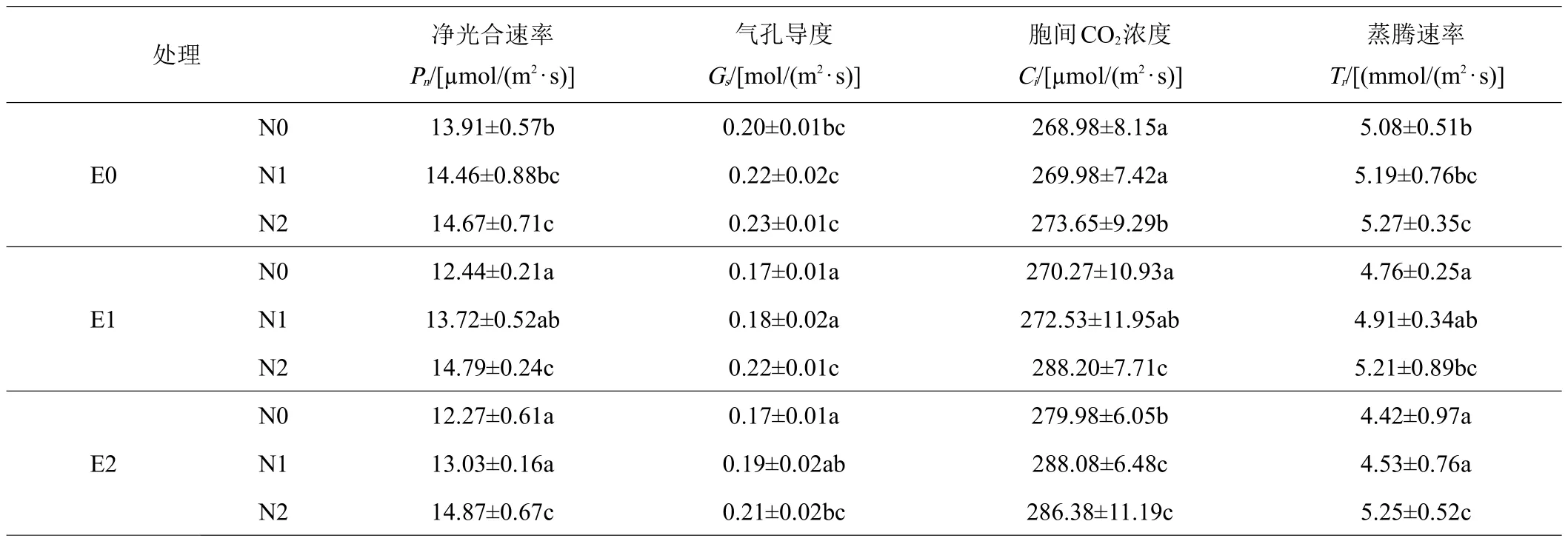
表1 不同处理对小叶章气体交换参数的影响
CO2熏蒸72天后,N0条件下,E1气孔导度、蒸腾速率较E0条件下降低15%(P<0.05)和6%,E2二者则分别降低15%和13%(P<0.05);而N1条件下,E1处理气孔导度、蒸腾速率较E0条件下降低18%和5%,E2处理二者则分别降低14%和13%(P<0.05),相比较而言,N0和N1条件下,CO2浓度升高对气孔导度的影响更显著;N2条件下,CO2升高条件下气孔导度、蒸腾速率与E0处理相比变化不显著(P>0.05,表1)。说明施用高浓度的氮肥,一定程度会缓解CO2浓度升高对小叶章气体交换参数的负效应。
2.2 不同处理对小叶章可溶性蛋白、氮素、可溶性糖和淀粉含量的影响
由图2a可知,N0条件下,E1和E2小叶章叶片可溶性蛋白含量较E0分别低7%和9%(P<0.05),E2处理差异达到显著水平;N1条件下,E1和E2处理可溶性蛋白较E0分别增加7%和8%,但未达到显著水平;N2条件下,E1和E2处理叶片可溶性蛋白含量较E0分别增加13%和18%(P<0.05),差异达到显著水平。在相同CO2浓度下,施氮也显著增加了小叶章叶片可溶性蛋白含量。CO2浓度升高显著影响了小叶章叶片N素含量(图2b)。N0条件下,E1和E2处理N素含量较E0处理分别降低了4%和9%(P<0.05);N1条件下,E1和E2处理小叶章叶片N素含量较E0处理显著降低(P<0.05);N2条件下,E1和E2处理小叶章叶片N素含量较E0处理变化不大(P>0.05)。
3个氮处理条件下,CO2浓度升高均促进了小叶章叶片可溶性糖含量的增加(图2c)。N0、N1和N2条件下,E1小叶章叶片可溶性糖含量较E0分别增加24%(P<0.05)、6%和16%(P<0.05);E2叶片可溶性糖含量则较E0分别增加11%(P<0.05)、3%和24%(P<0.05),除了在N1条件下未达到显著水平,其余均达到显著水平。3个氮处理条件下,CO2浓度升高对小叶章叶片淀粉含量影响均不显著(图2d)。
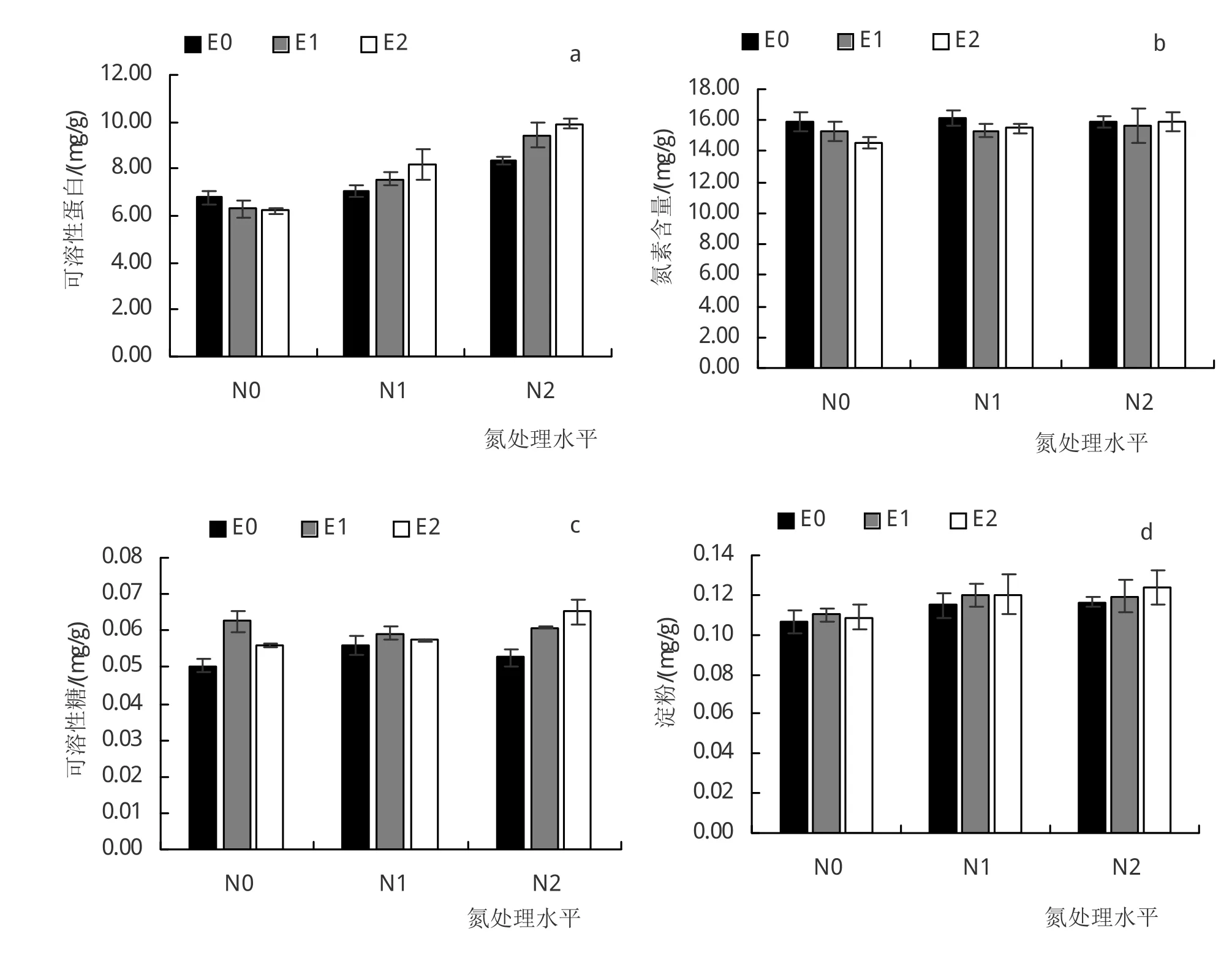
图2 不同处理对小叶章叶片可溶性蛋白、N素含量、可溶性糖、淀粉含量的影响
2.3 不同处理对小叶章株高的影响
由图3可见,N0条件下,E1和E2小叶章株高较E0处理降低9%和18%(P<0.05);而N1条件下,E1和E2小叶章株高较 E0降低 4%(P<0.05)和 2%(P>0.05);N2条件下,二者株高则有稍增加的趋势。在CO2浓度升高条件下,小叶章株高均随着施氮量的增加而增加。
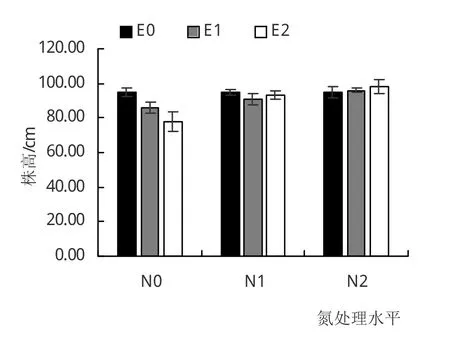
图3 不同处理对小叶章株高的影响
3 结论与讨论
通常情况下,短时间CO2浓度升高会促进植物光合作用,但随着时间延长,CO2浓度升高对光合效率的积极作用会慢慢减弱,甚至出现光合适应现象[19]。一般来说,可利用养分(nutrient availability)在植物和生态系统对CO2升高响应过程中起着重要作用[20]。氮是生态系统中影响植物生长的最重要因素。Sefcik等[21]研究发现,高CO2处理的植物发生光合适应一般受到氮素限制,高氮会削弱植物在高CO2条件下的光合作用下调[6]。
本研究结果表明,CO2浓度升高短期内可促进小叶章光合作用,但随着处理时间的延长,未施氮条件下,CO2浓度升高处理第72天,CO2升高处理的净光合速率显著低于自然CO2浓度条件下小叶章的净光合速率;而高氮条件下小叶章在CO2浓度升高条件中则未出现明显的光合下调现象,这与Hymus[22]等的研究相似,即在N供应缺乏时植物在富CO2下出现光合作用下调,而正常供N时,则未出现光合下调。
植物发生光合下调的机理目前还不十分清楚。一些研究表明,CO2浓度升高可能影响植物碳水化合物的汇和源间的平衡。当碳水化合物的汇规模较小或碳水化合物的传输受限制,那么植物叶片可能就会积累过多碳水化合物,最终反馈抑制光合效率导致光合下调[23-25]。但也有研究认为,终端光合产物合成速率是高CO2条件下植物光合作用下降的原因,淀粉大量积累不能成为植物光合下调的主要因素[26]。
本研究中,长期CO2浓度升高条件下出现光合下调的小叶章其可溶性糖含量较自然CO2浓度增加,但未出现光合下调的小叶章叶片可溶性糖含量也显著增加,这与赵光影等[15]的研究较一致,而与王晓等[13]的研究结果不同。有研究发现,CO2浓度升高处理不同物种、不同熏蒸时间植物叶片碳水化合物也有所差异。例如有研究发现CO2浓度升高条件下,小麦叶片淀粉和可溶性糖含量未显著增加[27];CO2浓度升高对植物叶片可溶性糖含量的影响与季节变化和土壤营养有关[28]。光合适应并不总是与高浓度CO2条件下可溶性糖积累相伴出现[29]。本研究说明光合下调的原因可能并不是由于光合产物积累的,可能是由于光合关键酶活性的降低影响光合作用。
Rubisco是叶片中最丰富的蛋白质,占全部叶片(可溶性蛋白质)的30%~50%,因此可溶性蛋白的降低在一定程度上可以表明Rubisco活性的下降[30]。本研究中,未施氮条件下,长期CO2浓度升高条件下小叶章叶片可溶性蛋白含量显著降低,因此推测出小叶章Rubisco含量也降低,进而影响小叶章光合效率,最终表现为小叶章株高的下降。徐明仪[16]在相同CO2浓度栽培试验研究中发现,高CO2浓度处理24 h后的小叶章,其叶片有3个编码rbc S的基因表达上调,这些基因表达水平的上调可以提高Rubisco的含量,进而短期内促进了小叶章光合能力的提高。Takatani等[31]发现长期CO2浓度升高条件下编码Rubisco的2个亚基(rbcS、rbcL)基因表达下调,最终影响光合作用。此外,也可能由于小叶章在高CO2浓度下,改变了调节光合适应有关基因的表达或者加速叶片衰老,因此导致植物发生光合下调[32-33]。而高氮条件下可改善小叶章光合关键酶活性,调节长期CO2浓度升高条件下对小叶章光合作用的影响,促进植物生长。
植物光合特性和生长对于CO2浓度升高的响应和适应是复杂的,氮素对其的调控作用不仅存在物种之间的差异,而且可能受生物和非生物因素的影响,未来还需从植物碳氮代谢以及基因表达方面进行长期系统的研究。
猜你喜欢
High Technology Letters(2021年4期)2022-01-09 02:08:16
河北果树(2020年4期)2020-11-26 06:05:00
海峡姐妹(2019年1期)2019-03-23 02:42:40
中成药(2018年9期)2018-10-09 07:18:46
新课程·下旬(2017年7期)2017-08-14 11:12:50
西北林学院学报(2015年5期)2015-01-03 05:12:20
植物营养与肥料学报(2011年5期)2011-11-06 07:30:52
植物营养与肥料学报(2011年2期)2011-10-26 03:52:30
植物营养与肥料学报(2011年2期)2011-10-26 03:52:10
植物营养与肥料学报(2011年3期)2011-10-24 06:15:00
