
豆科作物氮素高效利用机制研究进展
2022-07-08 09:26:06刘颖张佳蕾李新国张正万书波
中国油料作物学报 2022年3期
刘颖,张佳蕾,李新国,张正,万书波
(山东省农业科学院农作物种质资源研究所/山东省作物遗传改良与生态生理重点实验室/农业农村部华东地区作物栽培科学观测实验站,山东 济南,250100)
氮素是限制作物生长和产量形成的重要元素,目前农作物高产所需氮源主要来自工业氮肥,其过量施用一直是我国农业发展中的突出问题[1,2]。与工业固氮相比,豆科根瘤菌共生固氮体系是农作物获取氮素最有效、最经济的途径,共生固氮体系作为农业生态系统重要的氮素输入途径,不仅为作物提供清洁、高效的氮素营养,还改良了土壤结构、提升了土壤肥力[3,4]。因此,研究豆科作物氮素高效利用机理,对降低农业生产成本,促进农业可持续发展、保护生态环境具有重要意义[5]。
虽然豆科作物高效的生物固氮能力对减少氮肥投入、提高产量做出了重要贡献,但外源氮素与结瘤固氮拮抗作用一直是研究难点,特别是高浓度氮对结瘤和根瘤固氮的抑制作用[6,7]。当土壤有效氮含量较高时,豆科植物固氮量受抑制,导致“氮阻遏”的出现,即在土壤有效氮含量较高时,固氮酶活性受抑制的情况[8]。目前,豆科作物如何感知环境中氮素浓度的变化,进而做出精细调控的机制尚不清楚。因此,揭示外源氮对根瘤固氮的调控机制,解析根瘤固氮与外源氮的协同效应,对发挥生物固氮潜力具有重要意义。本文总结了豆科作物共生固氮体系的建立、豆科植物-根瘤菌对外源氮的响应、微生物和根瘤菌共生效率的关系,系统阐述了豆科作物氮素高效利用的机理,拟为豆科作物高产高效提供一定的理论依据。
1 共生固氮体系的建立
豆科植物-根瘤菌共生固氮是目前效率最高的生物固氮方式,固氮量占生物固氮总量的一半以上,多年研究测定大豆、菜豆等豆科植物年固氮量为每公顷50~300 kg,三叶草、苜蓿可达100~500 kg[9]。共生固氮是非常复杂且精细调控的过程,自1990 年起,已鉴定了近200 个基因参与共生固氮的调控,包括根瘤菌诱导信号通路的激活和根瘤原基形成所需转录因子的表达过程,以及特殊的根瘤结构的形成[10,11]。与此同时,根毛卷曲将根瘤菌包裹在里面,沿着侵染线被释放到宿主细胞,内化的细菌被宿主植物生物膜包裹成细胞,形成类菌体。固氮作用主要发生在成熟根瘤的类菌体中,由固氮酶将N2转化为NH4+[12]。多数豆科植物能通过这个结瘤机制形成具有固氮能力的根瘤,然而固氮植物在生态系统中所占的比例很小,固氮作用也受各种因素的影响[13,14]。一般来说,豆科作物根瘤固氮对氮素积累的贡献率仅占40%~60%,外源氮的供应对根瘤数量、形态建成以及固氮能力也有较大的影响[15]。
2 豆科植物-根瘤菌对外源氮的响应
豆科植物-根瘤菌共生固氮无法满足豆科高产栽培对氮素的需求,加入外源氮非常必要,但肥料氮的过量加入会对根瘤固氮产生强烈的抑制,即“氮阻遏”现象,高浓度外源氮对根瘤固氮的抑制作用是制约氮素高效利用的重要原因[16]。目前在豆科植物氮素的需求定量方面还缺乏研究,同时也包括最为关键的能引发根瘤菌“氮阻遏”效应的氮素阈值研究,导致现在的施肥技术难以精准吻合豆科植物对氮素的需求,因此,合理施用氮肥成为影响豆科植物发挥固氮功能的一个关键因素。
2.1 氮肥施用量对根瘤固氮的影响
一般认为作物合理施氮量为最高产量施氮量[17],但豆科植物施氮特点和要求与其它非豆科作物不同,其高产施氮量不一定有利于高效固氮,由于结瘤植物的根瘤固氮是消耗能量的过程,当大量氮素进入土壤时,固氮植物会转向耗能较小的方式,即吸收土壤有效氮[18,19]。王树起等[20]等发现随施氮量的渐进式增加,根瘤干重及数量均呈先增加后降低的趋势,可见,较低水平施氮会促进豆科植物自身的固氮能力,且达到最大固氮效率,超过这个临界则产生抑制作用。如石海等[21]等在大豆上的研究结果表明,54 mg·kg-1为促进根瘤形成和发挥最佳固氮功能的临界氮水平。截至目前,我国豆科作物大多以最高产量施氮达到的水平作为目标产量,这样造成氮肥损失略大。因此,准确判断豆科作物经济最佳施氮量及合理的施肥手段是提高豆科作物固氮能力的关键。目前判断临界氮水平的方式一般为植物体内的各种生理生化指标,例如根系形态相关指标、氮代谢关键酶活性、叶绿素含量和光合参数、固氮特性等影响氮素利用的活跃因素。其中,豆科植物固氮特性无疑是更严谨可靠的指标,固氮能力强也意味有较高的固氮效率。此外,15N示踪技术可将外源氮与其它来源的氮区分开,能更准确地研究豆科植物体内不同的氮素来源与分配,使得植物固氮特性成为研究氮素利用率的关键指标。
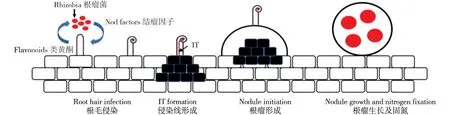
图1 根瘤菌-豆科植物共生体系的建立Fig.1 The establishment of symbiosis system of legume plant-Rhizobium
2.2 氮素形态对根瘤固氮的影响
氮素有多种化合态形式,不同形态氮素对豆科植物共生固氮体系结瘤固氮影响并不一致。以往研究表明[22]铵态氮在根毛变形前抑制结瘤,硝态氮则在根毛变形后至皮层细胞分裂前起抑制作用。与铵态氮相比,硝态氮对豆科植物结瘤的抑制作用更强,表现为铵态氮培养的植物的根瘤生长及固氮酶活性明显高于硝态氮培养[23~25]。但Yamashita等[26]研究表明,1 mmol·L-1硝态氮、铵态氮对大豆根瘤生长和固氮活性的抑制作用相当。多数学者认为硝态氮对结瘤性状及固氮酶活性的抑制作用高于铵态氮,说明不同氮素形态对豆科作物根瘤形成生物抑制存在多种作用机制,氮素浓度可在一定程度上影响两者的抑制效果。
2.3 氮肥施用时期对根瘤固氮的影响
豆科作物不同生育期对氮素的响应具有特异性,乔云发等[27]认为生育前期施氮可抑制根瘤固氮,生育后期追施氮肥则减轻抑制作用。郑永美等[28]研究则发现前期施用少量氮肥可提高根瘤固氮能力。在低氮条件下,根瘤主要吸收外源氮,这样避免了根瘤发育与宿主植物竞争氮素,达到了促进根瘤发育的目的。但在豆科作物生长初期,正处于根瘤形成和发育阶段,过低的土壤氮水平影响植株生长,进而影响结瘤过程和固氮能力[29]。董守坤等人[30]研究也表明,根瘤生长需要一定量的“启动氮”,且氮浓度为50 mg·L-1时利于根瘤生长,在高于50 mg·L-1受到明显抑制。生育中期以直接吸收根瘤固氮为主,较高浓度的外源氮对根瘤固氮产生抑制作用,生育后期,特别是生殖器官形成期,由于肥料氮的流失及根瘤的衰老,根瘤氮和土壤氮的供应不能满足花生的生长需求,及时补充氮肥不仅可以缓解根瘤衰老,还促进干物质向荚果转化[31]。因此,氮肥按需施用、精准投入是解决豆科植物氮素营养的关键。
3 根际微生物与根瘤菌共生效率的关系
豆科植物-根瘤菌共生是植物-微生物共生的一种重要类型,根瘤菌通过改变土壤微环境间接影响豆科作物的生长,同样,豆科植物根际存在的特定微生物群落也影响根瘤菌的结瘤及共生固氮效率。以大豆为例,根际微生物群落主要以中华根瘤菌和慢生根瘤菌组成,加上土壤中细菌、真菌等微生物,共同组成了复杂的群落[32]。Pérez-Jaramillo等[33]对菜豆核心根际微生物群研究也发现,菜豆核心微生物群由61 个变形杆菌组成,其中根瘤菌属为最丰富的菌,其次为慢生根瘤菌、伯克霍尔德菌、新生根瘤菌和鞘氨醇单胞菌。对天然土壤进行核心微生物群分析发现,变形杆菌、酸杆菌、疣状菌、拟杆菌、放线菌和氯曲菌是最丰富的门。在变形杆菌属中,拉尔斯通菌是最丰富的属,其次才是伯克霍尔德菌和根瘤菌。土壤中存在数十亿种的微生物,根瘤菌可与其宿主豆科植物的根际微生物竞争以建立共生关系,而且与天然土壤相比豆科植物选择性地从土壤中富集与养分循环有关的根瘤菌,其根系分泌物的类型及总量均影响了微生物群落结构[34]。另外,研究发现一些根际微生物与根瘤中的慢生根瘤菌和中华根瘤菌的组成有关,一种根际细菌-芽孢杆菌可显著影响根瘤菌的结瘤与固氮,Han 等[35]进一步研究发现,蜡状芽孢杆菌特异性促进和抑制根瘤菌的生长,并缓解了盐碱条件对中华根瘤菌结瘤的影响。因此,根际微生物群落在大豆根瘤菌-宿主相互作用中起关键作用,并为通过使用合成细菌群落提高这种共生体系的共生效率提供了框架,有益微生物和根瘤菌的联合使用可有效促进豆科植物的结瘤共生,为以后合成细菌群落提高大豆-根瘤菌共生固氮效率提供了科学依据。
微生物间合作与竞争可能有利于豆科植物-根瘤菌的结瘤,根瘤菌与豆科植物建立共生关系的前提是选择性识别、侵染,而侵染取决于根瘤菌与微生物的竞争力,还受不稳定环境因素的影响。正如Mason 等[36]发现大豆快生和慢生根瘤菌在不同pH值土壤中生长的结瘤率有很大的差异。Zahran[37]也认为酸性土壤不利于豆科植物的生长,特别是土壤pH<5 时,植物的结瘤效率大大降低,这与Wood等[38]研究结果基本一致。另外,长期大量氮肥输入可能引起土壤酸化,固氮菌处于酸性环境会导致固氮酶水解过程的发色团发生变化,影响固氮反应的正常运行。Fan 等研究表明[39],长期施肥可以通过降低与氮素固定效率呈正相关的关键物种和系统聚类固氮微生物的相对丰度,从而间接降低氮素固定效率。目前普遍认为,长期施肥增加了微生物总量、加剧微生物间的竞争,此时通过耗能的方式固氮不利于固氮菌的存活,根瘤菌的丰度随之降低,进而降低共生固氮量。
4 豆科氮素高效利用的机制研究
4.1 生理机制
氮素高效利用主要取决于氮素吸收、运输和利用能力,其中吸收利用效率起决定性的作用,而根系的发育能力和活性直接影响植物对氮素的吸收利用[40]。魏海燕等[41]研究表明,与低氮效率水稻品种相比,高氮效率水稻的根系发育能力更强,在玉米的研究中也发现根干重和根表面积存在差异是影响氮效率的原因[42]。以上研究均说明根系形态参数是评价植株氮效率主要的筛选指标,可见从根系角度研究氮效率差异,可在一定程度上为揭示氮高效机理提供依据。
植物吸收的氮肥需进一步同化成谷氨酸、谷氨酰胺等有机氮才能被植物利用,而豆科植物根瘤固氮中的固氮酶可将N2还原为氨,直接被植物吸收利用[43]。固氮酶活性是衡量豆科作物固氮效率的重要指标,与根瘤数、鲜重、根瘤固氮量、供氮比例、豆血红蛋白含量呈正相关,增强根瘤菌侵染能力和根瘤生理活性,可显著提高固氮酶活性[44]。另外,外源NO3--N 只有通过硝酸还原酶(NR)的催化还原为亚硝酸,由亚硝酸还原为氨才能被植物利用。NR 是植物NO3--N同化的关键酶,在植物氮代谢起着重要作用,氮高效品种对NO3--N 的同化能力相对较强[45]。
植物吸收同化NH4+-N的途径有两种:谷氨酰胺合成酶(GS)-谷氨酸合成酶(GOGAT)途径和谷氨酸脱氢酶(GDH)途径。GS、GOGAT 和GDH 是NH4+-N同化的关键酶,谷氨酰胺合成酶与氮效率间关系密切,呈显著正相关关系[46]。由此可见,氮素高效吸收利用各环节紧密相连,提高根系对氮素的吸收能力、向收获器官运输能力以及地上部分对氮素的利用能力是提高氮肥利用率的重要措施。
4.2 分子机制
4.2.1 结瘤自主调控(AON)控制结瘤 豆科植物适当的结瘤数目能保证氮素的高效利用,根瘤数目过多将会额外消耗能量,对植物正常的生长发育造成不利影响,根瘤数目过少则不能维持生物固氮的需求,因此,豆科植物形成了一套系统性调控根瘤数量的负反馈调节机制,称为结瘤自我调节(AON),主要通过地上-地下长距离信号传导[47]。其基本过程如下:首先是激活过程,根毛接收结瘤因
子(NF)即被激活;其次地上-地下传递系统,根部产生信号分子被运至地上部,被地上部的类受体激酶识别后合成新的信号分子再运送至地下部,其中一类富含亮氨酸的类受体激酶CLE 家族识别接受来自根部信号分子(即RS),CLE-RS 通过木质部转运至地上部,地上部识别后产生一种对于结瘤具有抑制作用的信号(SDI)运输到根部,从而实现对根瘤数目的控制[48]。以往的研究表明[49],AON 途径涉及的基因如果发生突变,会出现超结瘤的表型,表现为过度固氮,消耗掉大量的光合产物,对自身生长发育产生消极影响。最新研究表明[50],GmNINamiR172c-NNC1 分子模块是结瘤与AON 信号通路的主要开关,动态精细控制大豆的最佳结瘤数。这种自调节的机制在结瘤过程中达到豆科植物能承受的阈值时开启,保证了结瘤过程顺利进行,为豆科作物共生固氮和氮素营养高效利用提供重要的保障。
4.2.2 NIN-like proteins(NLPs)转录因子介导硝酸盐抑制结瘤 NIN(nodule inception)是共生和结瘤启动必需的转录因子NIN 的同源蛋白NLP(NINlike protein)在调节硝酸盐抑制结瘤共生的过程中有一定作用。在拟南芥基因组中发现了9个NLP基因[51],作为响应硝酸盐信号的核心转录因子,NLP7在感受到硝酸盐后可从细胞质穿梭到细胞核,调控硝酸盐信号相关基因的表达。Lin等人[52]还发现,蒺藜苜蓿中的5 个NLP基因可与NIN 蛋白相互作用,与拟南芥的NLP7一样,NLP1也能响应硝酸盐信号,从细胞质进入细胞核,通过PB1 结构域与NIN 形成复合体,并抑制NIN 影响下游因子CRE1,NF-YA1等的激活,最终抑制根瘤的形成及固氮。可见NLP1感受硝酸盐信号之后,通过干扰NIN 的功能来抑制共生。此外,Nishida 等[53]在百脉根中也发现了NRSYM1(LjNLP4)不仅参与硝酸盐抑制共生固氮过程,还参与抑制侵染线形成及根瘤发育。以上说明除了AON 途径以外,豆科作物还通过其它途径控制共生固氮过程,其中的分子机制还需要进一步探索。
5 氮素高效利用调控途径
关于豆科作物氮素高效利用的研究,前人已开展了大量工作,对生产技术起到了很大指导作用,而对于不同生育期氮肥精准投入,充分发挥根瘤菌的固氮效应和外源氮的协同效应,全生育期提高氮肥利用率则研究甚少。提高豆科作物氮素利用率是合理施肥实践中的长期任务,因此,确定豆科植物适宜目标产量,应以经济最佳施氮量所得产量为参考,既能充分发挥作物固氮潜力,又尽量保持在最为关键的能引发根瘤菌“氮阻遏”效应的氮素阈值之下,如果目标产量过高,反而会造成减产,导致高投入低产出;如果目标产量过低,则不能发挥土壤、品种的产量潜力。其次,选用氮高效品种,依据不同基因型豆科作物对不同氮源的吸收利用特点及差异,选用根瘤固氮能力较高的氮高效品种。另外,确定合适的施氮时期及施氮量也很重要,应根据豆科作物不同生育期对氮素需求的特异性,明确生育前期能充分发挥根瘤固氮效应和促进植株生长的“起爆氮”最佳质量、生育中期导致“氮阻遏”效应的氮素阈值、补充固氮不足的最少氮素用量以及生育后期延缓植株衰老改善叶片生理机能并能提高荚果饱满度的“后援氮”最优用量。针对不同生育时期外源氮对根瘤固氮的影响,实现氮肥按需施用、最大发挥自生固氮潜能,可突破豆科作物氮肥高效利用理论研究瓶颈,对解决豆科作物氮肥减施增效的重大关键科技问题具有重要意义。
猜你喜欢
山东冶金(2022年1期)2022-04-19 13:40:14
中国南方果树(2022年1期)2022-01-28 07:39:16
农技服务(2021年7期)2021-09-24 04:13:02
上海金属(2021年4期)2021-07-28 12:33:22
上海金属(2020年6期)2021-01-04 12:25:52
科学导报(2020年82期)2020-12-23 05:45:21
河北果树(2020年1期)2020-02-09 12:31:36
科学导报(2020年83期)2020-01-13 05:10:28
植物营养与肥料学报(2019年11期)2019-12-13 05:55:04
现代园艺(2017年21期)2018-01-03 06:42:11
