
桉树人工幼龄林根区和非根区土壤属性特征分析
2022-06-29 09:31:56马小芳陈凯鹏陈丹王聪李勇何斌杨梅
农业现代化研究 2022年3期
马小芳,陈凯鹏,陈丹,王聪*,李勇,何斌,杨梅
(1. 广西大学林学院,广西 南宁 530004;2. 南宁师范大学北部湾环境演变与资源利用教育部重点实验室,广西 南宁 530001;3. 中国科学院大气物理研究所,大气边界层物理与大气化学国家重点实验室,北京 100029)
桉树(Eucaiyptusspp.)是桃金娘科桉属植物,常绿乔木,种类多,适应性强,具有调节气候、涵养水源和保持水土的作用。桉树人工林是我国南方重要的速生用材林,广西桉树速丰林面积达到 256 万hm2,年产桉树木材 2 100 万m3以上,约占全国木材总产量的 25%,有力保障了我国木材生产基本储备与供应安全[1]。但是,在桉树带来巨大经济效益的同时,由于桉树生长速度快,轮伐期短(6~7 a),对土壤养分需求量大等特点导致桉树人工林地力衰退、林分产量下降等问题[2]。根区土壤是桉树吸收养分的核心区域,桉树可以通过根系生理活动将部分光合产物传输到土壤中,并从土壤中吸收水分与养分,根区土壤属性特征直接影响桉树的生长状况。由于桉树生长速度较快,幼龄桉树非根区土壤会在桉树种植后第2~3 年转变为根区土壤,该区域土壤理化性质和细菌丰度能直接对桉树生长产生重要影响[3]。因此,研究幼龄桉树根区、非根区土壤属性及微生物群落特征对桉树人工林经营有重要意义。
幼龄期人工林地非根区地上部分通常遍布草本植物,植物群落结构和组成较根区更复杂,这会引起根区、非根区植物吸收养分量、凋落物和根系分泌物差异,进而影响土壤理化性质。Gao 等[4]在苹果树—花生间作系统中发现,苹果非根区植物根系较少,吸收养分量较少,因此土壤有机碳(SOC)和全氮(TN)含量较高。郭新送等[5]在桃园生草还田对土壤微生物的影响研究中发现,桃树非根区生草自然还田可以促进非根区SOC 的积累并减少土壤氮素损失。由于植物根区、非根区土壤物质循环和能量流动,以及植物根系对养分的吸收作用,非根区土壤养分含量增加可能会间接改善根区土壤养分循环状况,从而促进植物生长[6]。因此,研究桉树根区、非根区土壤理化性质对桉树生长具有一定的指导作用。
生态化学计量学是研究生态系统中多种化学元素平衡的科学,C、N、P 是土壤主要的养分元素,其相对组成能影响有机质分解、微生物种群动态和根系养分吸收等一系列生物化学进程[7]。Zheng 等[8]在比较不同气候带的林地和旱地土壤C ∶N ∶P 的研究中,通过与全球尺度上温带森林,热带、亚热带森林[9]或全球森林[10]平均土壤C ∶N ∶P 的比较发现旱地土壤低N ∶P 和高P 含量,表明旱地土壤相对的氮限制。朱育锋等[11]对不同林龄桉树人工林土壤C ∶N ∶P 生态化学计量特征进行研究发现,与我国陆地生态系统土壤C∶N∶P均值相比,土壤C∶N、C ∶P 偏高,N ∶P 偏低,整体呈现高 C、低 N 和低P 的土壤元素格局。桉树根系发达能吸收深层土壤养分,而该研究仅比较了0~10 cm 土壤C ∶N ∶P,且未对桉树根区和非根区进行研究。研究桉树人工林土壤化学计量比可以揭示桉树生长过程中土壤养分的限制作用。目前,仅有针对不同林龄桉树人工林土壤C、N、P 化学计量特征的少量研究,而对桉树幼龄林根区、非根区土壤化学计量学特征的研究较少。
细菌在土壤微生物中数量最多、分布最广,主要参与土壤有机物质的矿化、无机养分的循环与转化等过程,是养分元素生物地球化学循环的主要驱动者[3]。Zhou 等[12]报道了荒漠豆科植物根区细菌多样性及结构差异,研究发现土壤含水量(SWC)、SOC、TN、全磷(TP)含量高的根区土壤细菌丰度更高。Elliott 等[13]通过研究不同施氮量对植物根区细菌数量的影响发现,高施氮量处理会提高细菌丰度。Wang 等[14]对不同耕作方式下根区土壤细菌丰度变化的研究表明,免耕保留了凋落物和地下根系,能提高土壤有机质含量,从而增加根区土壤细菌丰度。段春燕等[15]在不同林龄桉树人工林土壤 养分含量及生物学特性的研究中发现幼龄林(1 a)SOC、TN、TP、TK 含量和细菌数量随土壤深度增加而降低,但该研究仅比较了不同土壤深度间土壤养分含量和细菌数量的差异,未对根区与非根区土壤进行研究,且采用稀释平板计数法测定细菌数量存在可培养微生物少、分析精度低等缺点。幼龄期(1~2 a)是桉树生长最为脆弱、对环境变化最敏感的阶段[16],该阶段生长状况对桉树人工林生产力有重要影响[17]。因此,桉树人工幼龄林根区和非根区细菌丰度特征的研究值得探讨。鉴于此,本文基于广西东门林场林业科学研究所试验基地,研究冬季和夏季桉树幼龄林阶段根区、非根区0~20 cm、20~40 cm 深度土壤细菌丰度和理化属性特征,探讨桉树人工幼龄林土壤碳氮磷丰缺状况,以期为桉树人工林的抚育管理优化和地力维持提供决策参考。
1 材料与方法
1.1 研究区概况
研究区位于广西壮族自治区崇左市扶绥县东门林场(107°15′~108°00′ E,22°17′~22°30′ N),该区域属于典型的亚热带季风气候,年均温度为21.2~22.3 ℃,1 月平均气温12.8~13.5 ℃,7 月平均气温27.9~ 28.3 ℃。年降雨量为 1 100~1 300 mm,主要集中在 6—8 月份。研究区域的桉树人工林均采用尾巨桉 (E. urophylla × E. grandis)在2019 年5 月进行造林,株行距为 2 m×3 m。造林时每株桉树施1.0 kg 生物有机肥和0.5 kg 复混肥作为基肥。造林后连续追肥两年,2020 年5 月每株桉树分别追施0.5 kg 生物有机肥和复混肥,2021 年5 月每株桉树追施1.5 kg 生物有机肥和0.5 kg 复混肥。其中,生物有机肥的N、P2O5和K2O 含量分别为10.52 g/kg、14.29 g/kg 和13.12 g/kg,复混肥的N、P2O5和K2O 含量分别为208.99 g/kg、32.10 g/kg 和96.67 g/kg。桉树林下灌木层优势种有潺槁、柘树、三叉苦、木姜子、雀梅、盐肤木、石岩枫、大青、白背桐和毛桐等;草本层优势种有五节芒、黄茅、飞机草、海金沙和铁线蕨 等。一年除草两次,分别在5 月和10 月进行。研究区域以丘陵为主,海拔81~96 m,坡度<5°,土壤为砂页岩发育而来的砖红壤。
1.2 样地设置与样品采集
研究以桉树幼龄林(1~2 年树龄)为研究对象,选取5 个立地条件、栽植树种和营林措施基本一致,且周围干扰较少的幼龄林尾巨桉人工林样地,样地大小为400 m2(20 m×20 m)。于2020 年1 月初(冬季)和2020 年7 月初(夏季)采集根区、非根区0~20 cm、20~40 cm 深度土壤样品。在每块样地按“S”形随机设置5 个采样点,每个采样点选取2 株长势中等的桉树,在植株垂直冠幅下(约距离桉树树干10~40 cm)使用直径为5 cm 的土钻采集0~20 cm(R1)和20~40 cm(R2)深度根区土壤;在距离桉树树干最远点(无桉树根系分布的位置,约1.8 m 处)使用直径为 5 cm 的土钻采集0~20 cm(NR1)和20~40 cm(NR2)深度非根区土壤[18]。挑出肉眼可见的动植物残体和石头,然后将每块样地5 个采样点的根区、非根区不同深度的土样分别混匀并装入聚乙烯封口袋中。每个样品分成3 份,1 份土样迅速用锡箔纸包好,放入纤维袋子,投入到液氮罐中,运送到实验室,置于-80 ℃冰箱保存,用于后续测定土壤细菌丰度;1 份土样放入4 ℃冰箱保存,用于测定土壤DOC 和DON 含量;1 份置于室外风干,而后研磨过100 目筛,分别装入塑料封口袋保存,用于测定TN、TP、TK 和SOC 含量。
1.3 土壤理化性质测定
土壤理化性质按照《土壤农化分析》进行测定[19]。 采用环刀法测定非根区土壤容重(BD);烘干法测定SWC;土壤TN 采用硫酸消解-连续流动化学分析仪(AA3,SEAL Analytical Ltd.,Germany)测定;通过0.5 mol/L K2SO3浸提-连续流动化学分析仪测定DON;0.5 mol/L K2SO3浸提-TOC 仪(MULTIN/C3100)测定DOC;SOC 采用重铬酸钾氧化-外加热法测定;TP 采用 NaOH 熔融-钼锑抗显色-紫外分光光度法测定;采用 NaOH 熔融-原子吸收法测定TK。
1.4 土壤DNA 提取与定量PCR
土壤微生物的总DNA 采用土壤DNA 提取试剂盒(UltraClean DNA Isolation kit,MOBIO)进行提取,提取方法严格按照操作说明进行。提取出来的DNA经1%琼脂糖凝胶电泳和NanoDrop 核酸蛋白测定仪(NanoDrop ND-1000,Germany)进行浓度和纯度检测。实时荧光定量PCR 在耶拿荧光定量PCR仪(QTOWER3G,Germany)上进行。使用细菌通用 引 物1369F (5’-CGGTGAATACGTTCYCGG-3’)和1492R(5’- GGWTACCTTGTTACGACT-3’)细菌16S rRNA 进行扩增[20]。选取含有目的基因片段的质粒,梯度浓度稀释后,进行标准曲线的绘制。标准曲线R2为0.999,扩增效率为1,最后根据标准曲线计算细菌拷贝数。
1.5 数据处理与分析
所有数据统计分析采用R 软件。运用单因素方差分析和独立样本T 检验对冬季和春季根区与非根区不同土壤深度土壤理化性质和细菌丰度差异显著性进行分析(P<0.05)。通过三因素方差分析检验根区、土壤深度、季节及其交互作用对土壤理化性质、细菌丰度的影响。运用皮尔逊(Pearson)相关分析来分析土壤理化性质和细菌丰度之间的关系。图表数据为平均值±标准误(n=5)。
2 结果与分析
2.1 土壤理化性质分析
分析结果表明,与冬季相比,夏季根区和非根区的SWC、DON 和SOC 含量显著提高,增幅分别为 18.0%~55.6%、33.4%~63.3%和7.7%~17.5%(表1)。 夏季充足的水热条件较冬季更适宜土壤微生物生长,从而加速动植物残体分解使夏季土壤DON 和SOC 含量增加。夏季土壤TK 含量较冬季显著降低,降幅为18.0%~20.5%。夏季非根区0~20 cm 和20~40 cm 土壤DOC 含量较冬季分别显著增加了20.8%和23.9%,这是由于夏季非根区草本植物根系分泌物较冬季多。与冬季相比,夏季根区0~20 cm、20~40 cm 和非根区0~20 cm 土壤TN 含量分别显著降低了21.2%、25.5%和12.5%。夏季根区和非根区0~20 cm 土壤TP 含量较冬季分别显著减少了16.7%和14.6%。

表1 土壤理化属性Table 1 Soil physicochemical properties
在冬季和夏季,非根区0~20 cm 土壤DOC、DON、SOC、TN、TP 含量较根区分别显著增加了15.3%和42.7%、63.7%和43.8%、19.9%和23.8%、13.9% 和26.5%、22.1% 和25.1%;非 根 区20~40 cm 土壤DOC、SOC、TN 含量较根区显著增加了33.7% 和54.7%、11.1% 和18.9%、9.5% 和51.5%。说明桉树人工幼龄林根区土壤养分含量低于非根区。在冬季和夏季,土壤DOC、DON、SOC、TN和TP 含量均表现为R2<R1、NR2<NR1(P<0.05);土壤BD 表现为R1<R2、NR1<NR2(P<0.05,表1)。结果表明,土壤DOC、DON、SOC、TN 和TP 含量随土壤深度增加而降低。
2.2 土壤C ∶N ∶P 化学计量特征
在夏季,与R2 相比,R1 和NR2 的C ∶N 显著降低(图1)。在冬季,桉树人工幼龄林各土壤深度根区和非根区C ∶N 没有显著差异。与冬季相比,夏季桉树人工幼龄林各土壤深度根区和非根区C ∶N和C∶P显著增加。在冬季和夏季,NR2的N∶P和C∶P显著高于NR1 和R2。在夏季,与R2 相比,R1 的N ∶P显著增加。夏季R2 的N ∶P 显著低于冬季。

图1 土壤C ∶N ∶P 化学计量特征(C ∶N ∶P 的摩尔数据)Fig. 1 Soil C ∶N ∶P stoichiometry characteristic
在冬季和夏季,桉树人工林根区、非根区0~ 20 cm 土壤C ∶N ∶P 化学计量比的范围为52 ∶4.8 ∶1~ 73 ∶4.5 ∶1(图2)。桉树人工幼龄林冬季根区和非根区0~20 cm 土壤C ∶N ∶P 化学计量比分别为61 ∶5.5 ∶1 和61 ∶5.2 ∶1,夏 季 根 区 和 非 根 区0~20 cm 土 壤C ∶N ∶P 化学计量比分别为79 ∶5.2 ∶1 和78 ∶5.2 ∶1,冬季根区和非根区土壤C ∶N ∶P 均低于Cleveland 和Liptzin 等[10]报告的全球森林土壤C ∶N ∶P 平均值(212 ∶14.6 ∶1)。冬季桉树人工林根区和非根区土壤C ∶N ∶P 化学计量比小于夏季。夏季桉树人工林根区土壤C ∶N ∶P 化学计量比较非根区高。

图2 基于不同季节根区、非根区土壤C ∶N ∶P(C ∶N ∶P 的摩尔数据)三元相图Fig. 2 Ternary plots of C ∶N ∶P (based on original C ∶N ∶P molar data) in root zone and non-root zone of different seasons
2.3 土壤细菌丰度
冬季和夏季细菌丰度分别为1.63×109~3.87×109copies/g 和5.49×109~8.20×109copies/g(图3)。NR1的细菌数量较R1、NR2 分别显著增加了113.9%和49.3%、138.1%和24.2%(P<0.05),这是因为桉树幼龄林非根区凋落物量较根区多,且主要分布在表层土壤。夏季R1、NR1、R2 和NR2 的细菌丰度较冬季分别显著增加了203.5%、111.9%、233.7%和306.2%,说明夏季充足的水热条件更适宜细菌生长。

图3 土壤细菌16S rRNA 基因拷贝数Fig. 3 Abundance of the 16S rRNA gene
2.4 根区、土壤深度和季节对土壤属性的影响
三因素方差分析结果表明,根区、土壤深度、季节均显著影响DOC、DON、SOC、TN、TP 含量和土壤细菌丰度。此外,根区对C ∶N、N ∶P 和C ∶P均有显著影响;土壤深度对BD、N ∶P 和C ∶P 影响显著;季节对SWC、TK、C ∶N 和C ∶P 有极显著影响(表2)。根区和土壤深度的交互作用对SWC、DON、SOC、TP、C ∶N、N ∶P、C ∶P 和土壤细菌丰度均产生显著影响;根区和季节的交互作用显著影响DOC、SOC、TN 和C ∶N;土壤深度和季节的交互作用显著影响TN 和TP 含量。根区、土壤深度和季节三者的交互作用显著影响TN 含量和C ∶N。
2.5 土壤理化性质、C ∶N ∶P 化学计量特征与细菌丰度的关系
运用Pearson 相关性分析发现,SWC 与TN、TK含量显著负相关,与C ∶N、C ∶P 和细菌丰度显著 正相关。BD 与DOC、DON、SOC、TN、TP 含量显著负相关。DOC 含量与DON、SOC、TN、TP 含量、C ∶P、细菌丰度显著正相关。DON 含量与SOC、TP 含量、细菌丰度显著正相关,与TK 含量显著负相关。SOC 含量与TN、TP 含量、C ∶N、细菌丰度显著正相关,与TK 含量显著负相关。TN 含量与TP、TK 含量和N ∶P 显著正相关,与C:N 显著负相关。TP含量与N∶P和C∶P显著负相关。TK含量与C∶N、C ∶P 和细菌丰度显著负相关。C ∶N 与C ∶P 和细菌丰度显著正相关,与N ∶P 显著负相关。N ∶P 与C ∶P显著正相关。C ∶P 与细菌丰度显著正相关(图4)。

表2 土壤理化性质和细菌丰度对根区(R)、土壤深度(D)、季节(S)响应的三因素方差分析(F 值)Table 2 Significant effects of root zone (R), soil depth (D), and season (S) on soil physicochemical properties and bacterial abundances (three-way ANOVA root zone × soil depth × season)
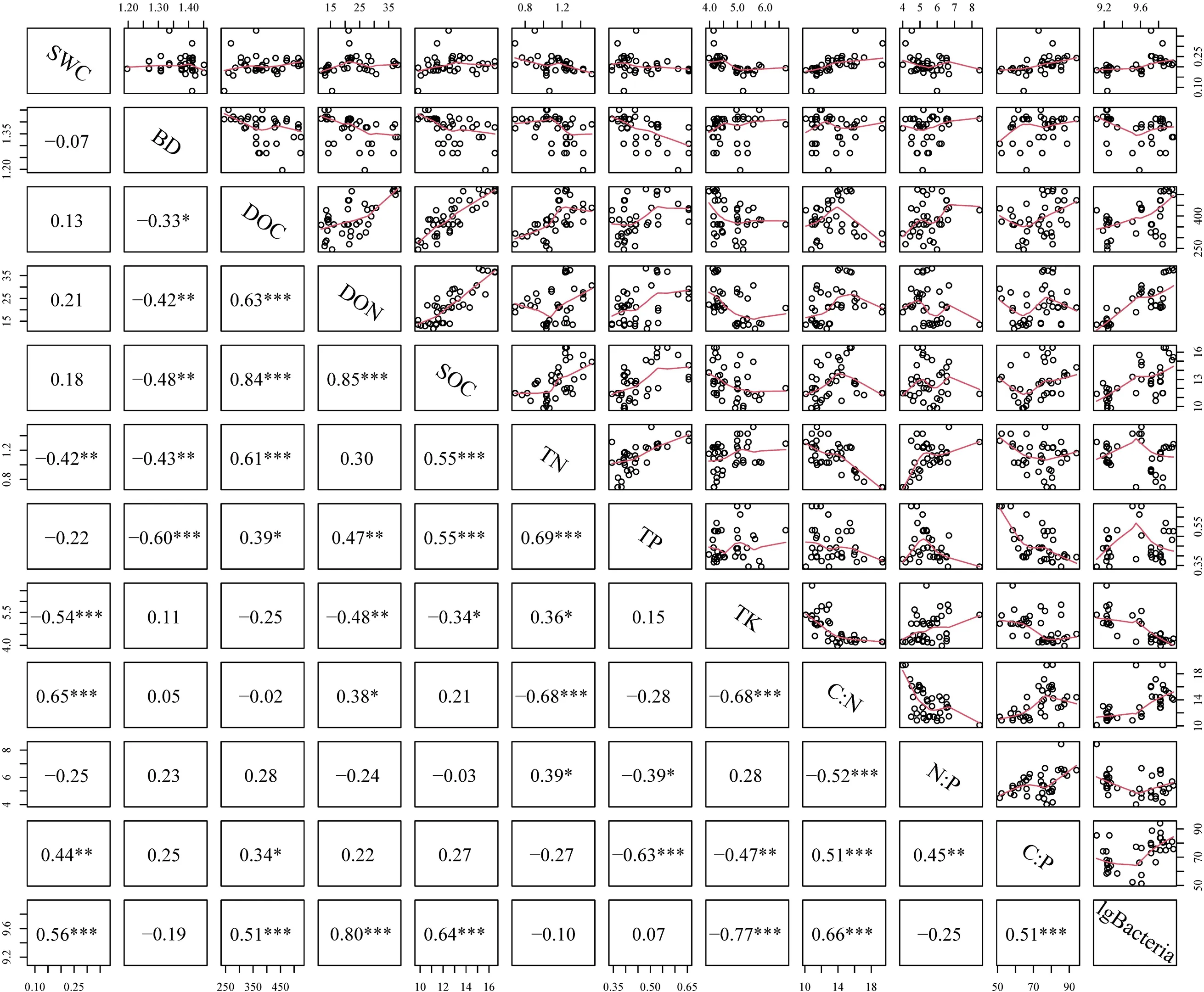
图4 桉树人工幼龄林土壤理化性质与细菌丰度之间的Pearson 相关性分析矩阵Fig. 4 Matrix of Pearson’ s correlation analysis of soil physicochemical properties and bacterial abundances in a young Eucalyptus plantation.
3 讨论
3.1 桉树人工林根区和非根区土壤理化性质的季节变化
桉树幼龄林时期由于郁闭度低,非根区土壤生长杂草较根区多,因此非根区的植物残体与根系分泌物较根区多[14],非根区土壤DOC、SOC、TN 含量显著大于根区土壤。由于非根区的植物残体与根系分泌物较根区多的缘故,非根区0~20 cm 土壤 DON、TP 大于根区土壤,但是非根区20~40 cm 土壤 DON、TP 与根区土壤并没有显著差异(表1)。这是因为DON 和土壤磷的移动性较弱,使非根区土壤 DON、TP 与根区土壤的显著差异主要发生在0~20 cm 土壤深度[21]。主要由于酸性土壤钾的水溶性较强[22],因此根区和非根区0~20 cm 土壤TK 含量与20~40 cm 土壤无显著差异。
由于夏季降雨较冬季多,所以夏季的SWC 更高(表1)。夏季较冬季有更合适的温度和SWC(21.4%~26.3%)使土壤微生物数量增加(如细菌),同时桉树幼龄林时期根区的易降解有机物质较少(主要为桉树根系分泌物和枯枝落叶),而非根区易降解有机物质较多(主要为杂草根系分泌物和残体)[23],因此夏季非根区DOC 含量较冬季显著增加,而夏季与冬季根区DOC 含量无显著差异(表1 和表2,图3)。虽然桉树幼龄林时期根区的易降解有机物质较少,非根区易降解有机物质较多,但是由于桉树根区微生物释放枯枝落叶中氮较碳更容易[14],因此夏季根区和非根区土壤DON 含量均较冬季显著增加。与之相似,夏季土壤微生物较冬季增加(如细菌),导致根区与非根区动植物残体分解形成SOC,因此夏季SOC 含量较冬季显著增加。该结果在Pearson 相关性分析中被验证:SWC、DOC、DON、SOC 与细菌数量显著正相关。
夏季充分的水分和光照下桉树和杂草生长较冬季旺盛,它们通过根系促进微生物矿化土壤氮为可利用态,然后吸收了大量土壤氮素[24];同时,由于桉树根系较草本植物更发达,能延伸到并显著促进更深土壤层次微生物矿化有机质释放氮素供其吸收利用[17],因此除非根区20~40 cm 土壤,夏季根区与非根区土壤TN 含量较冬季显著减少(表1 和表2,图3)。该结果在Pearson 相关性分析中被验证:SWC 与TN 显著负相关。与之相似,由于桉树幼龄林需求氮素较磷素多,磷素吸收主要集中在0~20 cm 土壤深度,且磷素在酸性土壤中移动性较差[21];杂草由于根系较浅,主要吸收0~20 cm 土壤磷素,因此夏季桉树根区与非根区0~20 cm 土壤TP 较冬季更低(表1 和表2)。与冬季相比,夏季适宜的温度和SWC 更有利于微生物矿化作用使土壤钾有效化[25]。土壤有效态钾在酸性土壤(pH=4.5)易随水流失,且被生长旺盛的植物吸收[22],因此夏季TK含量显著低于冬季。该结果在Pearson 相关性分析中被验证:TK 与SWC、细菌丰度显著负相关。
3.2 桉树人工林根区和非根区土壤C ∶N ∶P 化学计量特征
与全球森林土壤C ∶N ∶P 平均值相比[10],冬季根区、非根区土壤C ∶N ∶P 明显偏低,这可能与桉树人工幼龄林土壤C、N 含量较低有关。夏季桉树生长速度快,根系发达,吸收氮素较非根区草本植物快,且能通过根系促进微生物矿化有机质从而吸收土壤氮素[24],导致夏季根区0~20 和20~40 cm 土壤TN 含量减少,而0~20 cm 土壤的氮素由土壤表层动植物残体补充较20~40 cm 土壤快,因此夏季根区20~40 cm 土壤C ∶N 显著高于夏季根区0~20 cm、非根区0~20 和20~40 cm 土壤。冬季根区土壤有机质含量较低,依据全国第二次土壤普查技术规程规定的养分分级标准[26],评价结果表明桉树幼龄林冬季根区土壤有机质含量处于四级较缺乏水平(10~20 g/kg)。本研究桉树人工幼龄林冬季根区0~20 cm 土壤C ∶N 均值为11.1,低于全球森林土壤C ∶N 平均值(14.5)[10],因此,桉树人工幼龄林冬季根区土壤可能缺乏碳。参照全国土壤养分分级标准[26],桉树幼龄林夏季根区土壤TN 含量为四级较缺乏水平(0.75~1.0 g/kg)。夏季根区0~20 cm 土壤C ∶N 为15.3,高于全球森林土壤C ∶N 平均值,这可能与桉树人工林土壤TN 含量偏低有关,且桉树速生阶段(如幼龄林阶段)需吸收大量氮素促其生长[25],因此夏季桉树生长可能受到氮素限制。该结果在三元相图中得到验证。
与全球森林土壤N ∶P 平均值(14.6)相比[10],本研究中桉树根区、非根区0~20 cm 土壤N ∶P 均较低。可能的原因:1)桉树幼龄林和草本植物生长旺盛,吸收养分以氮素为主,因此消耗了土壤大量氮素[27];2)与土壤磷素相比,长期桉树短周期连栽会消耗更多氮素[28];3)由于与南方红壤属于酸性土壤,氮素以淋失和排放等方式损失多,使土壤氮含量偏低[29]。因此,与土壤TP 相比,土壤TN 含量降低更多,使土壤N ∶P 偏低。结合夏季根区土壤TN 含量处于较缺乏水平的现状,推断夏季根区土壤可能缺氮。该结果在三元相图中得到验证,这与Smethurst 等[30]对不同林龄桉树人工林的氮素管理研究结果一致。夏季桉树人工幼龄林生长过程中表现出相对的氮限制,可根据桉树幼龄时期对养分需求的特性合理施用氮肥改善土壤养分供给,同时在桉树人工林非根区种植固氮绿肥(如紫云英等豆科绿肥)来提升土壤氮含量[31-32]。
由于非根区草本植物根系及根系分泌物多分布在0~20 cm 土壤,20~40 cm 土壤微生物活性较低,矿化能力弱,能积累较多SOC,同时磷素在酸性土壤中的移动性较弱[21,23],多滞留在表层土壤,而桉树根系可分布于0~20 cm 和20~40 cm 土壤,因此非根区20~40 cm 土壤C ∶P 显著高于根区0~20 cm、20~40 cm 和非根区0~20 cm 土壤。夏季SWC显著高于冬季,根区、非根区动植物残体较冬季更易分解形成SOC,且夏季植物生长需要吸收更多的土壤磷素[24],因此夏季土壤C ∶P 均显著高于冬季(表1,图1)。该结果在Pearson 相关性分析中被验证:C ∶P 与SWC、lgBacteria 显著正相关。与全球森林土壤C ∶P 平均水平(211.7)[10]相比,本研究根区和非根区0~20 cm 土壤C ∶P 均较低。C ∶P 低的原因可能是长期短周期种植桉树,导致桉树人工林土壤退化,SOC 含量降低[2]。根据全国土壤养分 分级标准,冬季根区土壤有机质含量处于四级较缺乏水平,综合以上分析推断桉树人工幼龄林冬季根区土壤存在相对的碳限制。该结果在三元相图中得到验证。桉树人工幼龄林土壤C ∶P 低,细菌生长相对受碳限制,土壤磷释放潜力大。Fan 等[33]和Costa 等[34]通过比较不同林龄桉树叶片N ∶P 发现中龄林和近熟林N ∶P>16,表明桉树生长后期可能出现P 限制[35]。因此在桉树人工林经营中可通过施加有机肥、土壤调理剂等措施提高南方酸性土壤pH 来提高土壤有效磷。
3.3 桉树人工林根区和非根区土壤细菌丰度特征
细菌主要分解较容易降解的有机物质(如DOC、DON 和SOC 的易降解组分)[36],非根区0~20 cm 土壤易降解有机物质较多(主要为杂草残体),而桉树幼龄林时期非根区20~40 cm 土壤(20~ 40 cm 土壤易分解的有机物质一般较0~20 cm 土壤少)和根区0~20 cm、20~40 cm 土壤易降解有机物质较少(主要为桉树枯枝落叶)。因此,桉树非根区0~20 cm 土壤细菌丰度显著大于根区0~20 cm、20~40 cm 和非根区20~40 cm 土壤(表2,图3)。 该结果在Pearson 相关性分析中被验证:DOC、DON、SOC 与细菌丰度显著正相关。与冬季相比,夏季充足的水热条件促进了动植物和微生物生长,进而通过增加凋落物、根系分泌物、加快土壤养分周转速率等方式[14],显著增加桉树根区和非根区0~20 cm 和20~40 cm 土壤DON 和SOC 含量,以及非根区0~20 cm 和20~40 cm 土壤DOC 含量(表1,图3)。夏季增加的SWC、DOC、DON、SOC 含量可为土壤细菌生长提供适宜的条件[12],因此夏季桉树根区和非根区土壤细菌丰度较冬季显著增加(表1 和表2,图3)。该结果在Pearson 相关性分析中被验证:SWC、DOC、DON、SOC 与细菌丰度显著正相关。
4 结论
研究表明,桉树人工幼龄林非根区土壤DOC、SOC、TN 含量高于根区,非根区0~20 cm 深度土壤DON 和TP 含量高于根区。与冬季相比,夏季根区、非根区土壤SWC、DON、SOC 和非根区DOC含量更高。冬季根区土壤SOC 和夏季根区土壤TN含量为四级较缺乏水平,桉树人工幼龄林冬季根区土壤存在碳限制、夏季根区土壤存在氮限制。细菌丰度特征表现为桉树非根区0~20 cm 土壤细菌丰度最高,夏季土壤细菌丰度高于冬季。根区、土壤深度和季节是影响桉树人工幼龄林土壤细菌丰度及理化性质的重要因素。本研究结果可为优化根区水肥管理、非根区绿肥种植等营林措施,进而提高桉树人工林生产力提供参考。
猜你喜欢
中国农业大学学报(2022年11期)2022-11-07 03:20:46
儿童故事画报·发现号趣味百科(2019年9期)2019-02-02 04:12:19
环球时报(2019-01-03)2019-01-03 09:06:44
农业与技术(2018年16期)2018-11-28 11:24:08
现代农业科技(2017年10期)2017-07-12 14:14:49
中国生态农业学报(中英文)(2017年2期)2017-02-16 08:01:30
广西林业科学(2016年2期)2016-03-20 05:53:22
广西林业科学(2016年4期)2016-03-16 05:44:51
中国农业信息(2016年17期)2016-02-06 09:50:05
农业与技术(2014年9期)2014-10-20 08:57:37
