
9 种莎草科植物种子萌发期抗旱性评价
2022-06-29 00:49:24李欣勇张靖雪王文强虞道耿
草业科学 2022年6期
李欣勇,张靖雪,王文强,盛 伟,虞道耿
(中国热带农业科学院热带作物品种资源研究所, 海南 海口 570100)
干旱导致的作物减产,超过其他环境因子胁迫所造成作物减产的总和[1]。随着全球气候变暖现象的持续,会增加全球高温时期及干旱面积,植物将会面临更严峻的干旱胁迫压力[2]。种子萌发是植物生活史中最脆弱的阶段,易受环境因子影响[3],种子萌发期对逆境的适应能力会决定植物种群分布和繁衍[4]。水分是一切生命活动所必需的,也是种子萌发的基础[5],干旱胁迫达到一定阈值时,会显著抑制种子萌发和幼苗生长[6]。因此,研究植物种子萌发期的抗旱性,将有助于推测植物在幼苗生长阶段的抗旱性,为提高该植物的建植率、培育抗旱新品种及新品种推广提供参考数据[7]。
莎草科(Cyperaceae)是陆地生态系统的重要构成者,广泛分布于各大洲,尤以热带和亚热带地区种类较多,具有水土保持、土壤改良、水源净化等作用,也是天然草地资源中饲用价值较高、耐牧性极强的一类优良牧草[8]。如高秆莎草(Cyperus exaltatus)、风车草(C. alternifolius)可作观赏植物;羽状穗砖子苗(Mariscus javanicus)可治理盐碱地;蕨状薹草(Carex filicina)、十字薹草(C. cruciata)、毛轴莎草(C.pilosus)等植物根、茎、叶的通气组织十分发达,具有固土保水、净化水体的作用[9]。目前对莎草科植物的研究主要集中在田间杂草种群调查[10-11]、净化水体[12-13]、湿地群落构建[14]、修复土壤重金属污染[15]、休眠类型[16]和萌发条件[17-20]等方面,对莎草科种子萌发期抗旱性的研究鲜有报道。因此,本研究通过不同浓度聚乙二醇(PEG-6000)溶液模拟干旱环境,对9 种莎草科植物种子萌发期的抗旱性研究和评价,以期为莎草科植物的种植和利用提供理论依据。
1 材料与方法
1.1 试验材料
本试验共选用莎草科种质资源9 种(表1)。所有种子采集后贮存于中国热带农业科学院热带作物品种资源研究所低温种子库中。
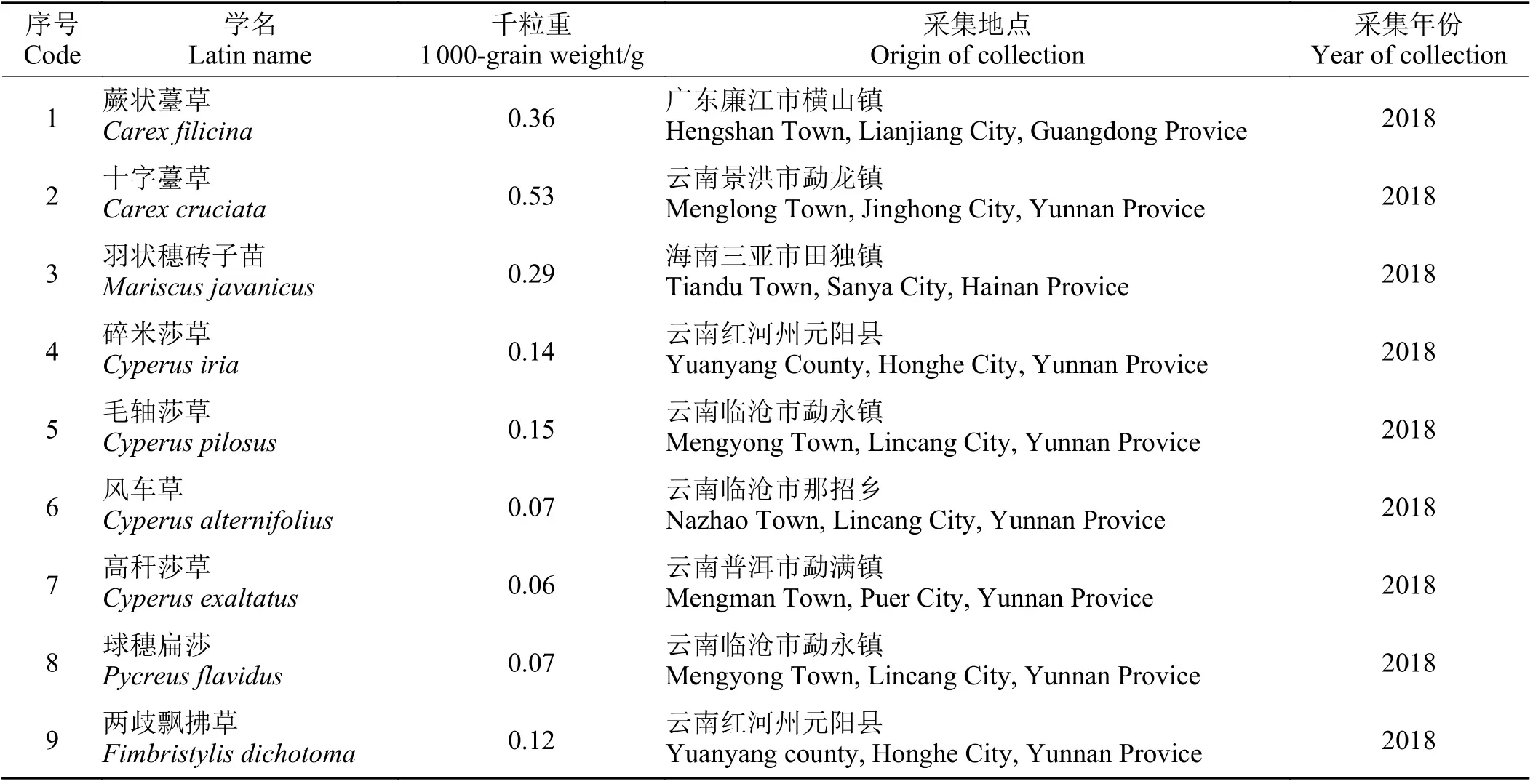
表1 9 种莎草科植物种子信息Table 1 Information of nine Cyperaceae species seeds
1.2 试验方法
1.2.1 种子预处理
每份种质材料挑选大小均匀一致、饱满的种质150 粒,用0.1% HgCl2溶液浸泡消毒10 min,再用蒸馏水冲洗3 次,用滤纸吸干种子表面水分备用。
1.2.2 试验设计
模拟干旱条件设置5 个处理,分别用10 mL 浓度为0 (CK)、5%、10%、15%、20%的聚乙二醇(PEG-6000)水溶液代替标准萌发试验中的蒸馏水。将培养皿置于人工智能气候培养箱中(昼夜温度为30 ℃/20 ℃;光/暗16 h/8 h)。为保持培养皿内水势不变,每2 d 换一次滤纸和溶液,每天调换培养皿在培养箱中的位置,确保所有种子的所处环境相同。每种处理3 个重复,每个重复50 粒种子,每天统计萌发种子数(以胚根突出种皮1 mm 视为萌发),持续28 d。
1.3 指标测定
为了消除种质材料之间本体萌发能力的差异,更好地进行胁迫条件下不同种质材料间的分析,本研究采用相对萌发率、相对萌发速率、相对萌发指数、相对胚芽长和相对胚根长进行萌发能力比较。
1.3.1 萌发率、相对萌发率、萌发速率、相对萌发速率、萌发指数和相对萌发指数
萌发率(germination rate,GR) = 试验结束时已萌发种子数/供试种子总数 × 100%;
相对萌发率(relative germination rate,RGR) = (胁迫处理萌发率/对照萌发率) × 100%;
萌发速率(germination speed,GS) = 1/T50。
式中:T50为种子萌发率达到最终萌发率的50%所需的时间。
相对萌发速率(relative germination speed,RGS) =(胁迫处理萌发速率/对照萌发速率) × 100%;
萌发指数(germination index,GI) = ∑ (Gt/Dt)。
式中:Dt为萌发天数,Gt为Dt相对应的每天萌发种子数。
相对萌发指数(relative germination index,RGI) =(胁迫处理萌发指数/对照萌发指数) × 100%。
1.3.2 胚芽长和胚根长
第15 天每盘随机选取10 株植株测量胚芽和胚根长度,取平均数。
相对胚芽长 = (胁迫处理胚芽长度/对照胚芽长度) × 100%;
相对胚根长 = (胁迫处理胚根长度/对照胚根长度) × 100%。
1.4 数据分析
本研究采用隶属函数法对不同莎草科种质资源的抗旱性进行评价。
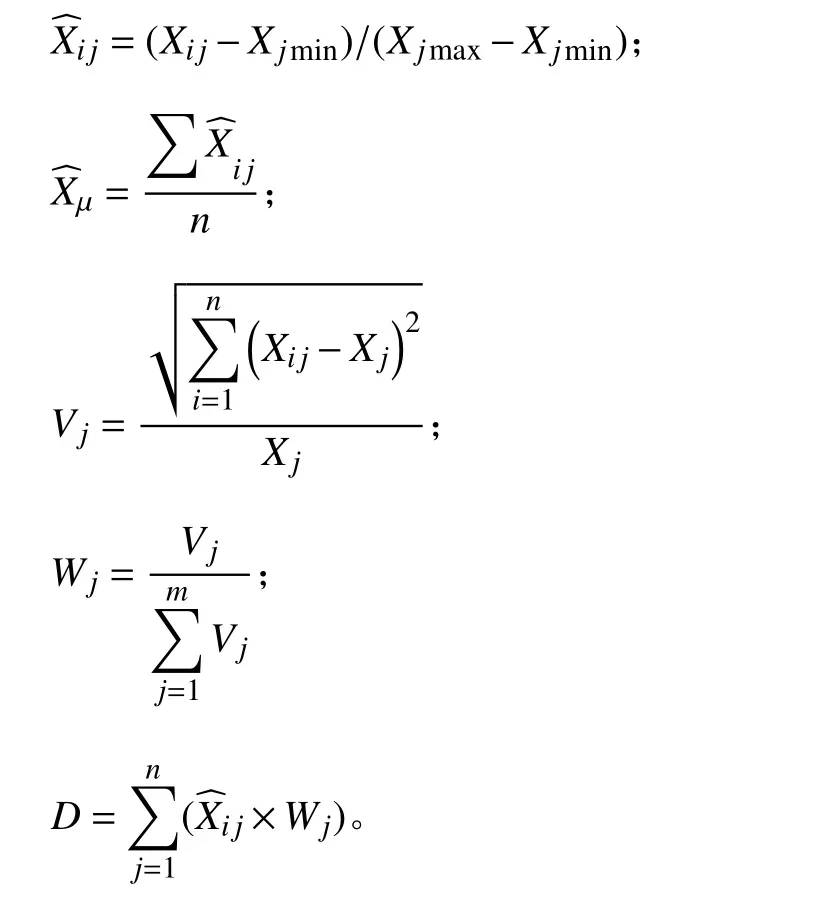
式中:Xij为第i种植物第j个指标的测定值;Xjmax和Xjmin分别为所有植物中第j个指标的最大值和最小值;Xμ为第i种植物第j个指标的隶属函数平均值。Vj为标准差系数,Wj为权重系数,D为综合抗旱值,D越大,其综合抗旱性越强。
所有数据的统计分析均采用SPSS 19.0 进行,多重比较采用Duncan 法,Word 2013 作表,所有数值均以平均值 ± 标准误 (Mean ± SE) 表示。
2 结果与分析
2.1 不同浓度PEG 对相对萌发率的影响
不同浓度PEG 胁迫对9 种莎草科植物的种子相对萌发率有显著影响(P< 0.05) (表2)。多数表现为随PEG 浓度的增加,萌发率呈下降趋势,但下降程度因物种不同而不同。蕨状薹草、十字薹草、碎米莎草和两歧飘拂草(Fimbristylis dichotoma)的种子相对萌发率在PEG 浓度≥5%时显著低于对照(P<0.05),其中十字薹草在15% PEG 浓度时的相对萌发率仅为4.63%,在20% PEG 浓度时不萌发。5% PEG浓度会促进羽状穗砖子苗和高杆莎草种子萌发,其种子相对萌发率高于对照。羽状穗砖子苗在15%PEG 浓度时种子相对萌发率仍有90.28%,但在20%PEG 浓度时却不萌发。毛轴莎草和球穗扁莎(Pycreus flavidus)的种子相对萌发率仅在PEG 浓度≥15%时显著低于对照(P< 0.05)。20% PEG 浓度时,蕨状薹草和毛轴莎草的相对萌发率仍有70%以上,显著高于其他物种(P< 0.05)。
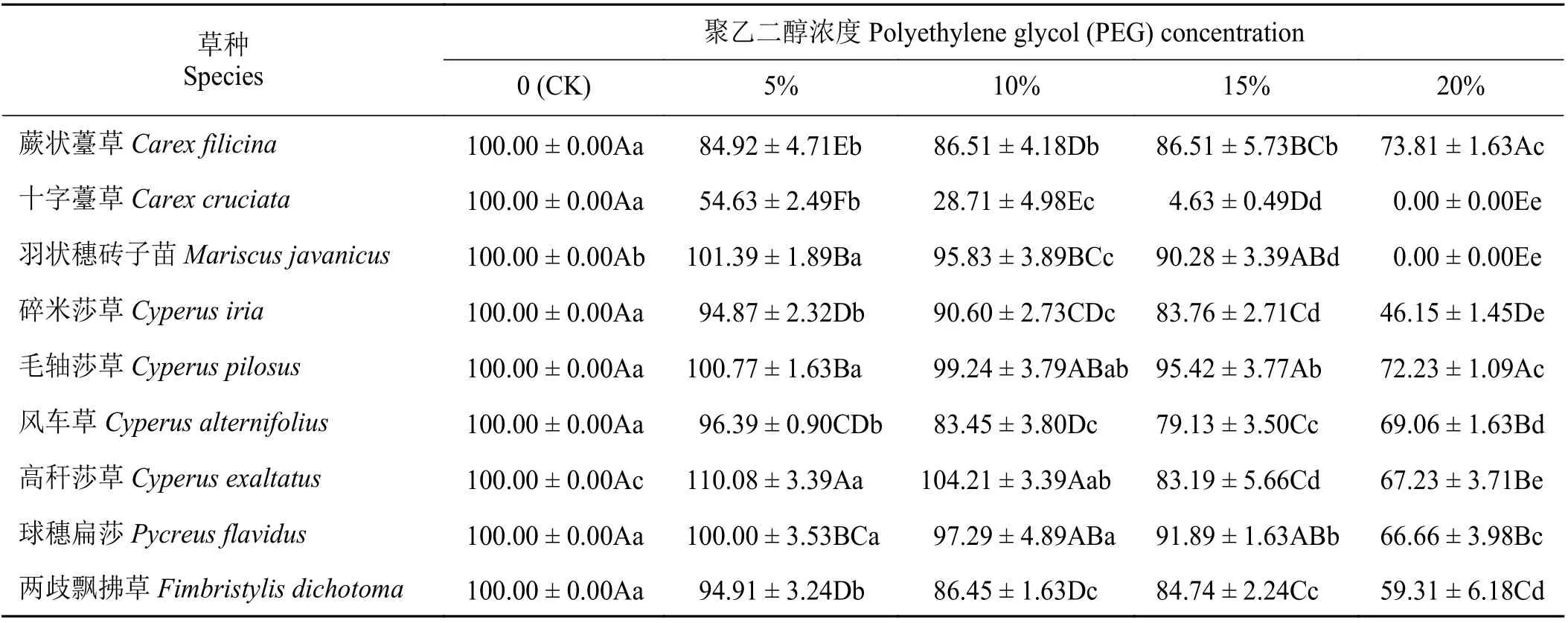
表2 PEG 胁迫对9 种莎草科植物相对萌发率的影响Table 2 Effect of PEG treatment on the seed relative germination rate of nine Cyperaceae species%
2.2 不同浓度PEG 对相对萌发速率和相对萌发指数的影响
不同浓度PEG 胁迫对9 种莎草科植物的种子萌发速率和萌发指数有显著影响(P< 0.05),多数表现为随PEG 浓度的增加,萌发速率和萌发指数呈下降趋势,但下降程度因物种不同而不同(表3、表4)。总体来说,随PEG 浓度的增加,相对萌发指数的下降速度均比相对萌发速率快,说明种子萌发指数对干旱胁迫更敏感。碎米莎草、风车草和球穗扁莎在5% PEG 浓度时的相对萌发速率显著高于对照(P< 0.05),说明轻微干旱胁迫反而能提高其萌发速率。20% PEG 浓度时,两歧飘拂草的相对萌发速率和相对萌发指数均为最大,显著高于其他物种(P< 0.05)。

表3 PEG 胁迫对9 种莎草科植物相对萌发速率的影响Table 3 Effect of PEG treatment on the seed relative germination speed of nine Cyperaceae species%
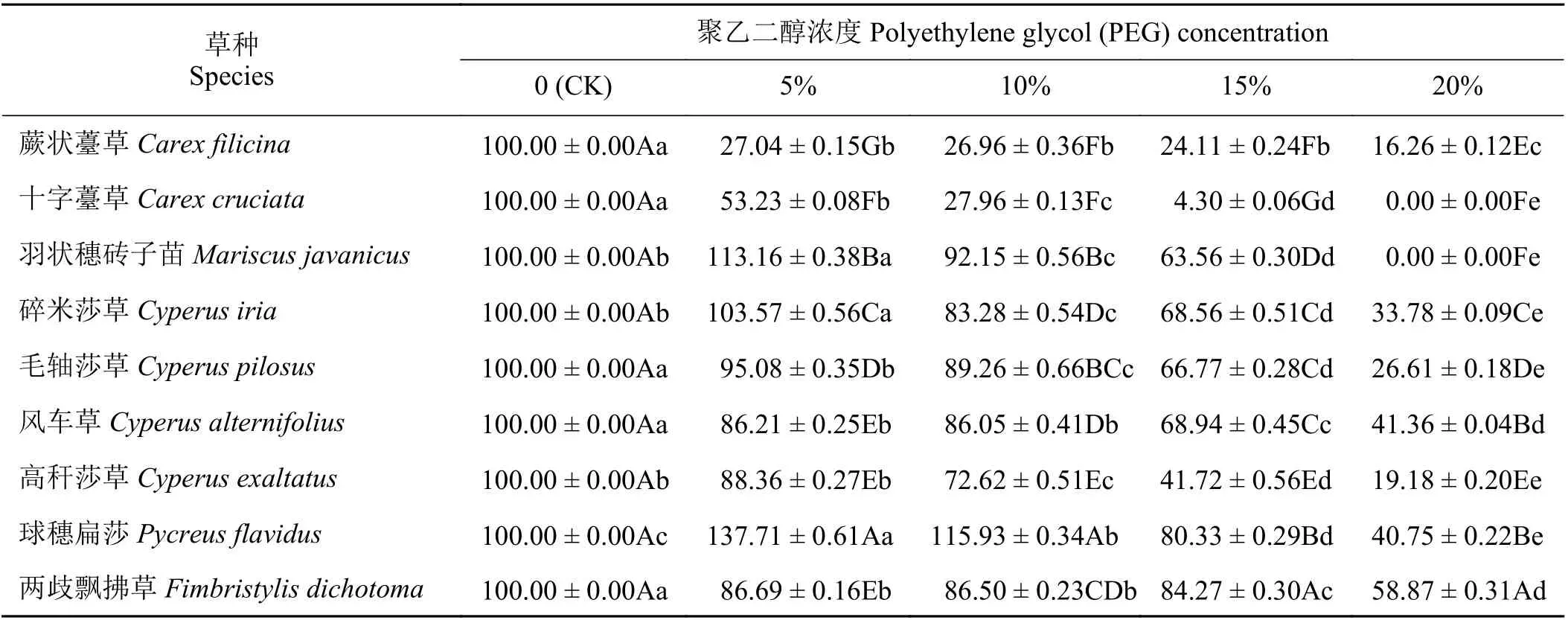
表4 PEG 胁迫对9 种莎草科植物相对萌发指数的影响Table 4 Effect of PEG treatment on the seed relative germination index of nine Cyperaceae species%
2.3 不同浓度PEG 对相对胚芽和相对胚根的影响
随PEG 浓度的增加,9 种莎草科植物的相对胚芽长和相对胚根长均呈不同程度的下降(表5、表6)。毛轴莎草的胚芽长在10% PEG 浓度时显著高于对照(P< 0.05),其余8 种莎草科植物的胚芽长均在5% PEG 浓度时出现明显下降,显著低于对照(P< 0.05)。20% PEG 浓度时,所有植物的胚芽生长都受到显著抑制,其中碎米莎草的胚芽下降最慢,为对照的68.82%,显著高于其他物种(表5)。5% PEG浓度胁迫有利于羽状穗砖子苗、毛轴莎草、风车草和两歧飘拂草的胚根生长,其胚根长显著高于对照(表6) (P< 0.05);其余物种的胚根对干旱胁迫较敏感,5% PEG 浓度胁迫时其胚根长已显著低于对照(P< 0.05);20% PEG 浓度时,碎米莎草和毛轴莎草的胚根长下降最慢,分别是对照的81.79%和81.48%,显著高于其他物种(P< 0.05) (表6)。所有植物的胚芽生长受干旱胁迫的影响程度明显大于胚根生长。
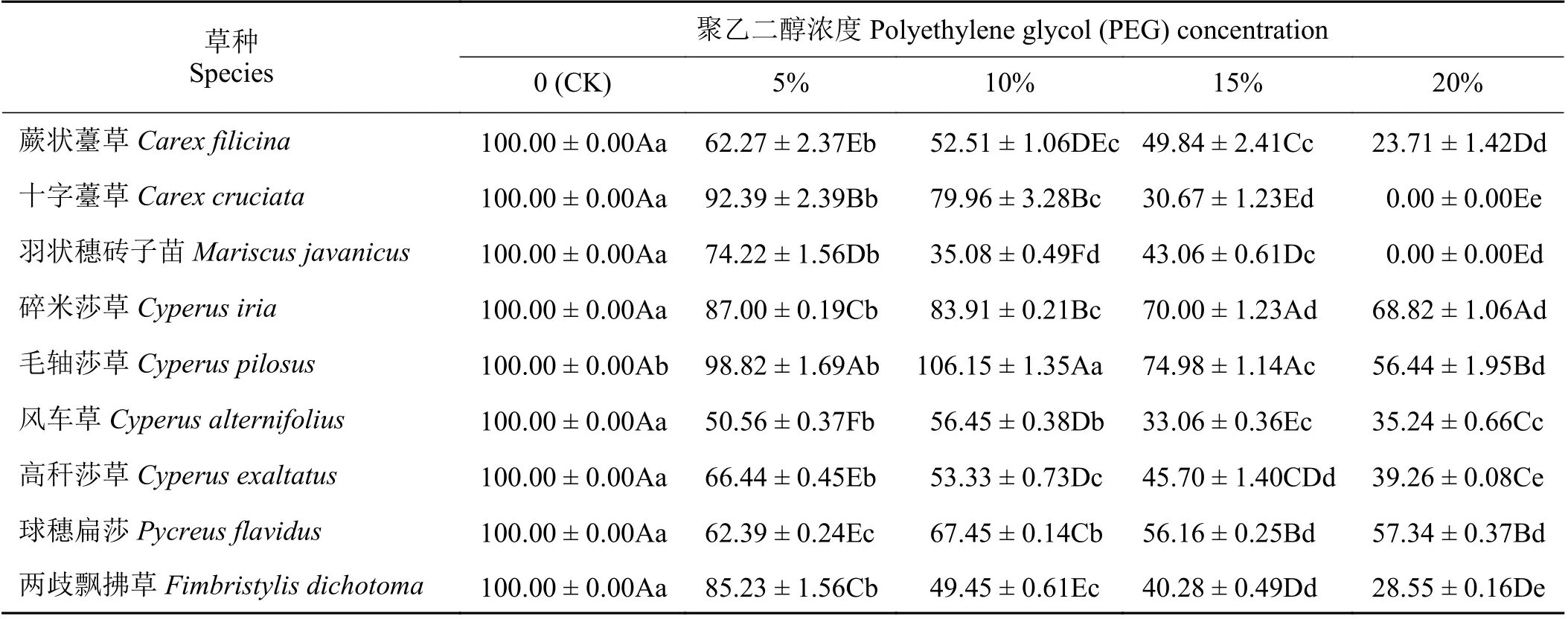
表5 PEG 胁迫对9 种莎草科植物相对胚芽长的影响Table 5 Effect of PEG treatment on the relative plumule length of nine Cyperaceae species%

表6 PEG 胁迫对9 种莎草科植物相对胚根长的影响Table 6 Effect of PEG treatment on the relative radical length of nine Cyperaceae species%
2.4 9 种莎草科植物萌发期抗旱性综合评价
为了全面综合反映植物的抗旱能力,采用相对萌发率、相对萌发速率、相对萌发指数、相对胚根长和相对胚芽长这5 项抗旱指标,计算9 种莎草科植物萌发期的综合抗旱能力(D值),以评价其抗旱性。结果表明,9 种莎草科植物5 项抗旱指标的隶属函数综合评价值为0.24~0.63 (表7),9 种莎草科植物种子萌发期抗旱性表现为毛轴莎草 > 碎米莎草 >高秆莎草 > 蕨状薹草 > 羽状穗砖子苗 > 风车草 >球穗扁莎 > 两歧飘拂草 > 十字薹草。

表7 9 种莎草科植物各指标隶属函数值及综合评价值Table 7 Subordinate function value and D-value of nine Cyperaceae species
3 讨论
种子萌发通常是植物生活史最为脆弱的阶段,容易受外界环境因子的影响[3]。植物种子萌发期对干旱胁迫的适应性与植物抗旱性相关,能在一定程度上反映该物种在干旱环境中的生存能力[21]。本研究结果表明,当PEG 浓度 > 5%时,9 种莎草科植物的种子萌发率、萌发速率、萌发指数、胚芽长和胚根长大多数随PEG 浓度的增加呈显著下降趋势,说明随着干旱胁迫程度的增加,对种子萌发的抑制作用也越强。这与在马齿苋(Portulaca oleracea)[22]、紫花苜蓿(Medicago sativa)[23]、沙冬青(Ammopiptanthus mongolicus)[24]等的研究结果一致。当PEG 浓度≤5%,羽状穗砖子苗和高秆莎草的种子相对萌发率显著高于对照;毛轴莎草和球穗扁莎的种子相对萌发率与对照差异不显著。这可能是由于轻度干旱胁迫下,植物种子会通过改变脯氨酸等调节物质来维持相对正常的渗透势,从而保证种子顺利萌发,甚至会促进种子萌发[25],与Hu 等[26]在‘兰箭3 号’箭筈豌豆(Vicia sativa‘Lanjian3’)和窄叶野豌豆(Vicia angustifolia)上的研究结果相似。低浓度(5% PEG浓度)的干旱胁迫能促进羽状穗砖子苗、毛轴莎草、风车草和两歧飘拂草胚根的生长,这可能是由于干旱条件下,植物幼苗为了增强从环境吸水的能力来保证植株生长,会优先增加胚根的生长速度,这与在紫云英(Astragalus sinicus)[27]、垂穗披碱草(Elymus nutans)[28]和黑黄檀(Dalbergia fusca)[29]上的研究结果相似。有意思的是羽状穗砖子苗在15% PEG 浓度时种子相对萌发率仍有90.28%,但在20% PEG 浓度时却不萌发;这说明羽状穗砖子苗种子在萌发期的需水量较少,萌发能力很强,虽然15% PEG 浓度胁迫会延缓羽状穗砖子苗种子萌发,降低其萌发速率和萌发指数,但对其最终萌发率影响不大;20%PEG 浓度已经超出了羽状穗砖子苗种子萌发所需的基础水势,所以种子不萌发[30]。
植物种子萌发期抗旱性是一个受多种因素影响的复杂生理生化过程[27]。本研究采用萌发率、萌发速率、萌发指数、胚芽长和胚根长5 项指标,结合隶属函数对9 种莎草科植物种子萌发期抗旱性进行综合评价发现,这5 项指标的单一隶属函数排序均不同,这进一步表明植物抗旱性受多种因素影响,只靠单一指标来评价植物抗旱性,结果相对片面。所以本研究采取多指标综合隶属函数评价方法,才能消除单一指标的片面性,研究结果才更加可靠。隶属函数综合评价显示,9 种莎草科植物种子萌发期抗旱性表现为毛轴莎草 > 碎米莎草 > 高秆莎草 >蕨状薹草 > 羽状穗砖子苗 > 风车草 > 球穗扁莎 >两歧飘拂草 > 十字薹草。
4 结论
本研究表明,低浓度(5% PEG 浓度)干旱胁迫能促进羽状穗砖子苗、毛轴莎草、风车草和两歧飘拂草胚根的生长。当PEG 浓度 > 5%时,9 种莎草科植物的种子萌发指标均随PEG 浓度的增加呈下降趋势,且种质之间差异较大。隶属函数综合评价,9 种莎草科植物种子萌发期抗旱性表现为毛轴莎草 >碎米莎草 > 高秆莎草 > 蕨状薹草 > 羽状穗砖子苗 >风车草 > 球穗扁莎 > 两歧飘拂草 > 十字薹草。
猜你喜欢
疯狂英语·新读写(2022年1期)2022-11-23 04:44:18
疯狂英语·读写版(2022年1期)2022-01-29 01:06:57
广西林业科学(2021年6期)2022-01-21 07:39:28
山西农业科学(2021年8期)2021-08-17 06:20:10
文史博览·文史(2021年1期)2021-04-01 14:04:00
青年文学家(2021年4期)2021-03-18 07:00:23
农民致富之友(2020年19期)2020-07-18 15:32:39
山西农业科学(2020年1期)2020-02-27 17:00:54
家庭百事通·健康一点通(2019年11期)2019-09-26 09:27:52
饮食科学(2014年8期)2014-08-22 17:42:07
