
江西柘林水库鱼类群落结构及功能多样性分析
2022-06-28 08:20孟子豪李学梅胡飞飞杜红春朱永久杨德国
生态学报 2022年11期
陈 康,孟子豪,李学梅,胡飞飞,杜红春,刘 璐,朱永久,杨德国,*
1 中国水产科学研究院长江水产研究所,农业农村部淡水鱼类种质资源与生物技术重点实验室, 武汉 430223
2 上海海洋大学,水产与生命学院,上海 201306
物种多样性是群落多样性的关键,既体现了物种与物种之间以及物种与环境之间相关关系的复杂度,又体现了生态系统中生物资源的丰富度。生物多样性是生态系统发挥功能的前提,是人类赖以生存和持续发展的基础[1]。淡水鱼类作为水生生态系统中重要的组成成分,在维持生态系统的稳定性、为人类提供食物来源以及社会经济文化服务方面发挥了重要作用。然而,随着经济水平的不断提升,社会发展的不断进步以及人类干扰强度的不断增大,淡水生态环境恶化、鱼类栖息地丧失以及生物入侵现象不断发生,给内陆淡水鱼类带来严重威胁,改变了鱼类群落结构,降低了鱼类群落生物多样性水平[2]。
近年来,生物多样性与生态系统功能关系成为了生态学研究领域的一个重大科学问题[3],已有的研究大多使用基于分类的物种多样性指数(Taxonomical diversity)表征生态系统中生物的丰富度。然而,简单的物种多样性并不能完全体现出生态系统的复杂度[4]。随着研究的逐渐深入,研究者发现物种功能性状与群落构建和生态系统服务功能具有紧密的关系,通过对功能性状多样性的研究,可以更加清楚的理解物种多样性和生态系统功能、多样性对环境干扰的响应规律,有利于深入了解生态系统功能变化机制[5]。对鱼类群落多样性的研究工作发现,更多物种和功能性状的出现预示着群落在面对环境干扰时具有更高的抵抗力[6]。因此,鱼类群落动能性状多样性通常被用来表征水生生态系统的稳定性、复杂性以及受损后的恢复力[7]。对鱼类群落多样性的研究更应该考虑加入功能多样性维度到生物多样性研究工作中,通过分析物种多样性与功能性状多样性之间的关系,对于监测生态系统的健康状况,分析不同功能类群之间生态位重叠的程度,解决水生生物的保护和生物多样性的管理决策问题提供了重要途径[8]。
尽管功能多样性的重要性被越来越多的研究者所认同,对功能多样性的研究也逐渐增多,但是,目前淡水鱼类功能生态学的理论和实践研究仍然非常薄弱[9]。江西柘林水库是长江中游鄱阳湖流域的大型峡谷型水库,库区水环境、水文及水生生物群落组成差异性显著,鱼类物种组成结构复杂。本研究基于柘林水库的鱼类季度调查数据,分析该水域鱼类群落多样性的时空格局变化特征,探讨鱼类群落物种多样性与功能多样性之间的相互关系,以期为该水域鱼类资源的合理利用和鱼类物种的多样性保护提供科学参考。
1 材料与方法
1.1 研究区域概况
柘林水库位于修河流域中上游,处于幕阜山脉和九岭山脉之间,西接修水、地跨永修和武宁两县,地理坐标为东经115°04′—115°40′,北纬29°03′—29°27′(图1)。水库汇水区域9340 km2,水域面积308 km2,正常水位65 m,平均水深16.3 m,最大水深45 m,总容量79.2亿m3,流域内大小支流有603条[10]。柘林水库目前规划建设有庐山西海鳡国家级水产种质资源保护区,调整后的保护区面积达到218 km2,对保护库区内鳡及其它保护物种包括桃花水母、鳜、蒙古鲌、似鮈等发挥了重要的作用。
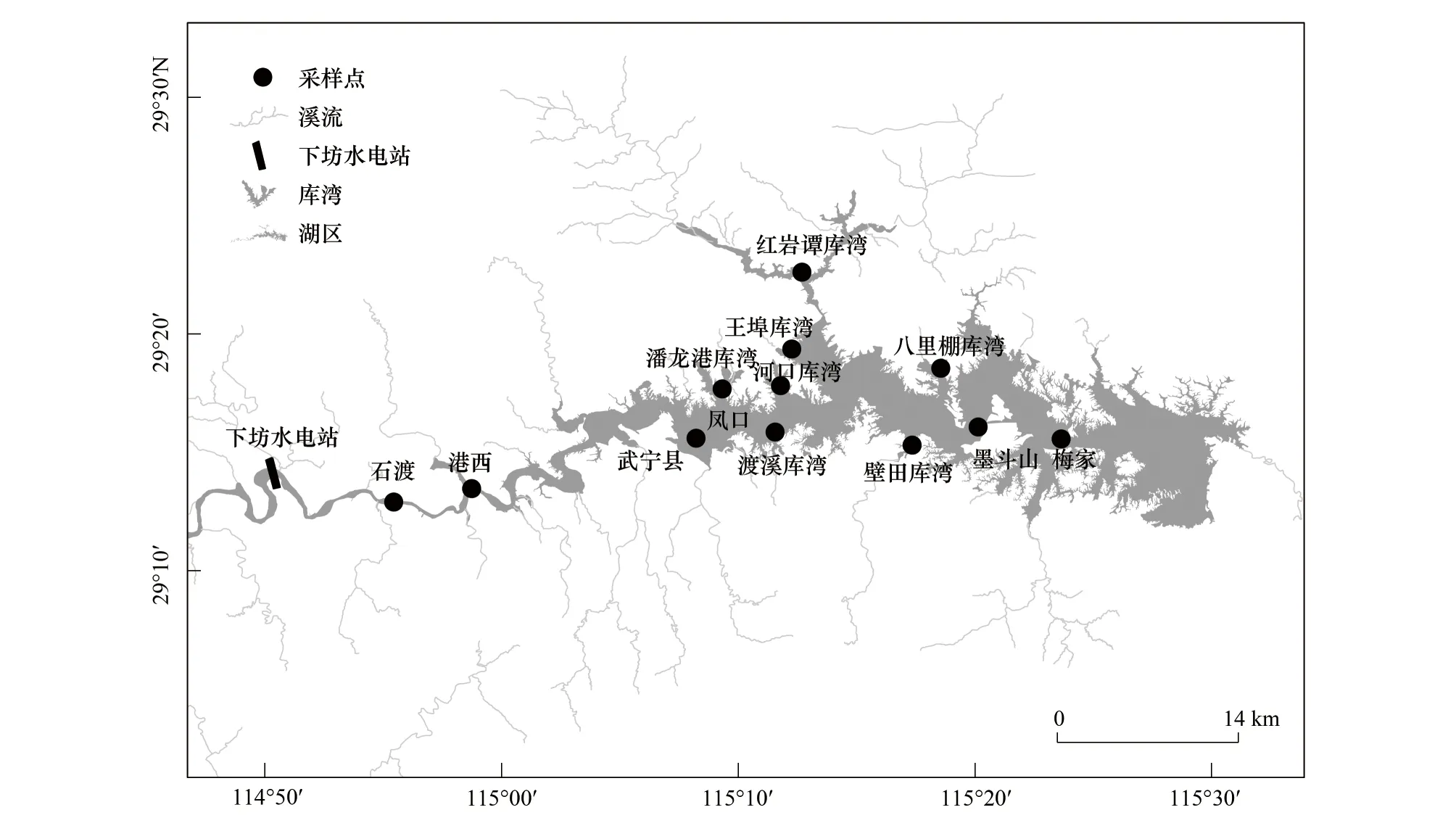
图1 柘林水库水系及采样点图Fig.1 Drainage in Zhelin Reservoir and sampling sites in this study
1.2 样点设置及调查方法
本研究样点分布于柘林水库的上游修河水域(石渡、港西)、中游库湾水域(潘龙港、河口、王埠、红岩潭、八里棚、壁田、渡溪)、及下游库心水域(墨斗山、梅家),于2020年9月、12月与2021年4月对库区分三个季度进行渔获物调查;三次调查共计28个采样点。为保证充分收集不同栖息水层环境中鱼类标本,使用定制三层流刺网(中层网目3 cm、8 cm、15 cm各50 m,网高分别为2 m、3 m和5 m)和地笼(截面30 cm×30 cm,网目1.2 cm,长度15 m)开展渔获物调查。网具放置过夜(12 h),对收集到的鱼类标本参照陈宜瑜[11]和朱松泉[12]进行分类鉴定,统计渔获物的全长(精确到0.1 cm)、体长(精确到0.1 cm)、体重(精确到0.1 g)等生物学信息。
1.3 数据分析
1.3.1群落结构组成分析
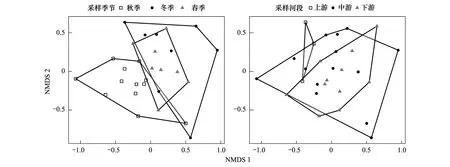
使用非度量多维标尺(Nonmetric Multidimensional Scaling, NMDS)排序方法评估鱼类群落结构在时间和空间格局下的差异性。对收集的样点鱼类物种数据矩阵进行log转换以降低极值对统计的干扰,采用Bray-Curtis相异距离构建物种在样点之间的距离矩阵。使用置换多元方差分析(Permutational Multivariate Analysis of Variance,PERMANOVA)检验鱼类群落结构在时空格局上的差异性显著水平,计算多元模型中的Fisher′s F-ratio值,并在999次随机置换过程中计算显著性P值[13]。
1.3.2物种多样性
为指示鱼类群落多样性的变化规律,使用以下物种多样性指数进行表示:
(1)Shannon-Wiener多样性指数(H′)
式中,Pi为第i种鱼类个体数占总个体数的比例,S为物种数;
(2)Simpson多样性指数(D)
式中,Pi为第i种鱼类个体数占总个体数的比例,S为物种数;
(3)Margalef丰富度指数(d)
d=(S-1)/lnN
式中,N为总个体数,S为样点总鱼类种数
(4)Pielou均匀度指数(J)
J=H′/lnS
式中,H′为Shannon-Wiener多样性指数,S为样点总鱼类种数;
(5)重要性指数(IRI)
IRI=(N%+W%)F%
式中,N%为某一物种鱼类尾数占总尾数的比例,W%为某一物种鱼类生物量占总渔获物生物量比例,F%为某一物种出现样点数占总样点数比例。IRI>1000为优势种,IRI在100—1000之间为重要种,IRI在10—100之间为常见种,IRI在1—10之间为一般种,IRI<1为少见种[14]。
1.3.3功能多样性分析
参照已有的鱼类功能多样性研究中所使用的功能性状,本研究通过查阅文献资料和Fishbase网站对采集到的鱼类物种识别其功能性状,主要包含营养等级、栖息水层(底层、中层、上层)、食性(底栖动物食性、草食性、浮游生物食性、肉食性、腐屑食性、杂食性)、运动方式(静止或运动)、身体形状(鳗鱼形、细长形、梭形、长方形、椭圆形)、感觉触须(有或无)、最大体长尺寸(等级1:0 参照Cornwell等[16]和Villeger等[7]计算功能多样性(Functional diversity, FD)的方法,使用凸壳体积法(convex hull volume)计算鱼类群落的功能多样性。凸包体是一个群落所填充的功能空间体积,凸包体积越大,则表明组合中功能丰富度越高(Functional richness, FRic)。利用功能均匀度指数(Functional evenness, FEve)量化物种在功能空间中的分布均匀度,物种拥有的功能性状越相似则该值越低。使用FDis方程计算功能离散度(Functional disperse, FDis)描述群落功能性状是如何分布,功能性状越集中则该值越大。功能冗余度(Functional redundancy, FRed)指示了群落中具有相同功能性状物种数的高低[17],代表了生态位的重叠程度,通常用Rao′s多样性指数与Simpsion的比值进行计算。 使用Kruskall-Wallis检验方法分析物种多样性及功能多样性在不同时空格局下的差异性,Dunn test两两比较方法用来检验不同分组之间生物多样性指数的差异性。使用一般加性模型(General Additive Model)检验物种多样性指数与功能多样性两者之间的关系,显著性水平设置为0.05。以上数据分析均在R语言中实现,其中,NMDS和PERMANOVA使用vegan包进行分析;功能多样性选择Gower距离矩阵在FD包中进行计算[18—19]。 本次调查在柘林水库共采集到鱼类53种,隶属5目12科36属(表1)。其中,鲤形目(38种)种类最多,占调查物种总数的71.69%;其次是鲈形目(7种)和鲇形目(6种),其他分类目种类较少。在科等级上,鲤科(37种)种类最多,占总数的69.81%;鲿科鱼类4种,鮨科鱼类3种,鳗鲡科、鳅科、鱵科、鲇科、鮰科、沙塘鱧科、鳢科、刺鳅科、虾虎鱼科各1种。柘林水库上中下游库区分别采集到鱼类36、40和32种,各库区均以鲤形目鱼类为主,分别占采集物种数的72.97%、85.00和75.00%,其他分类目物种数较少。 表1 柘林水库鱼类名录及功能性状Table 1 List of fishes and its functional traits in Zhelin Reservoir 续表 物种Species食性Food type营养级Trophic guild最大体长等级Maximum size栖息水层Location运动性Motility身体形态Body shape感觉触须Sensorial barbel种群恢复力Life span黄颡鱼 Tachysurus fulvidraco肉食性3.5等级3底层静止型长方形有中瓦氏黄颡鱼 Pseudobagrus vachellii肉食性3.5等级4底层静止型长方形有中细体拟鲿 Pseudobagrus pratti肉食性3.5等级2底层静止型细长形有中圆尾拟鲿 Pseudobagrus tenuis肉食性3.5等级3底层静止型细长形有中斑点叉尾鮰 Leporinus punctatus肉食性2.3等级5底层静止型细长形有中鲈形目 Perciformes鳜 Siniperca chuatsi肉食性4.5等级4中层游动型椭圆形无中大眼鳜 Siniperca knerii肉食性3.9等级3中层游动型梭形无中长体鳜 Siniperca roulei肉食性3.7等级2中层游动型梭形无中黄黝鱼 Micropercops swinhonis浮游生物食性3.2等级1底层静止型长方形无中乌鳢 Channa argus肉食性4.4等级5底层静止型细长形无低中华棘鳅 Sinobdella sinensis底栖动物食性3.3等级3底层静止型鳗鱼形无高子陵吻虾虎鱼 Rhinogobius giurinus底栖动物食性3.2等级2底层静止型长方形无高 表2 柘林水库不同季节鱼类优势种组成Table 2 Index of relative importance of fish species among seasons in Zhelin Reservoir 基于物种组成的非度量多维排序结果分析发现,空间格局下鱼类物种组成差异性不显著,上游、中游与下游区域物种组成结构相似,分离现象不明显(图2),置换方差检验结果未达到显著水平(F=0.724,P=0.792);季节梯度下,物种组成结构存在明显的分离现象,差异性达到极显著水平(F=2.565,P=0.002),两两比较结果显示鱼类群落结构的差异主要表现为秋季和冬季之间,达到显著水平(F=3.270,P=0.018)。综上,柘林水库鱼类物种组成结构在时间尺度上的差异性大于空间格局下的差异性。 图2 基于鱼类物种组成结构的非度量多维排序图Fig.2 Nonmetric Multidimensional Scaling based on species abundance data according to temporal-spatial patterns 鱼类物种多样性分析结果显示(图3),上游修河水域Margalef指数显著高于中下游水域(P=0.046),Shannon、Simpson与Peilou多样性指数在不同区域之间差异不明显(P>0.05);生物多样性指数在不同采样季节变化不显著。鱼类功能多样性分析结果显示,功能丰富度指数在空间格局下差异显著,上游修河水域鱼类功能丰富度显著高于中下游库区(P=0.044);功能均匀度、功能离散度在时间和空间格局下差异性不显著(P>0.05);鱼类群落功能冗余度相对较高,时空差异性不明显。总体而言,鱼类物种多样性和功能多样性空间格局差异性大于时间尺度下的差异性。 图3 柘林水库鱼类物种多样性与功能多样性指数的时空格局差异性Fig.3 Temporal-spatial distribution patterns of fish taxonomical and functional diversity in Zhelin Reservoir相同小写字母表示无显著性差异(P>0.05) 柘林水库鱼类群落物种多样性与功能多样性分析结果显示,两者之间存在显著的相关关系(图4);其中,功能丰富度FRic与物种多样性指数之间存在显著的正相关关系(P<0.05),与Margalef指数的相关系数R2最高为0.735;功能均匀度与物种多样性指数之间的相关性不显著;功能离散度FDis与物种多样性指数之间存在显著的正相关关系,与Simpson多样性指数的相关系数R2最高,达到0.803;功能冗余度FRed与各物种多样性之间呈现负相关关系,随物种多样性的升高而下降(表3)。 图4 物种多样性指数与功能多样性指数线性模型Fig.4 Regression models between taxonomical and functional diversity 表3 物种多样性指数与功能多样性指数线性可加模型分析Table 3 Additive models of taxonomical diversity and functional diversity 柘林水库各样点鱼类群落的生物多样性指数Shannon-Wiener(0.08—2.14)、Peilou(0.07—0.91)、Margalef(0.41—4.00)均表现较低,Shannon多样性指数低于Magurran[27]提出的一般范围之间(1.5—3.5),低于长江中下游湖泊洪泽湖(1.72—2.22)[28]、洞庭湖(1.68—2.52)[29]、鄱阳湖(1.54—2.58)[30]鱼类多样性水平,高于太湖(0.21—0.99)[26]鱼类多样性水平。时间尺度下物种多样性指数与功能多样性指数均未表现出显著的差异性,相反,空间格局下两者表现出不同程度的空间格局差异,主要为上游修河水域拥有相对较高的Margalef值以及FRic指数分布范围。产生这一结果的主要原因为上游单个样点渔获物种类数均高于其他样点,加之较低的个体数水平,从而导致了上游区域样点的Margalef指数高于中下游样点。此外,上游较高的FRic指数预示着较高的功能性状的丰富度,主要原因为上游修河水文形态和环境因子与库心区及库湾区存在明显差异性,石渡与港西采样点水体具有一定流速,基质类型主要为沙石,从而庇护了更多物种,包含喜好静水游动型鱼类(鲢、鳙、黄尾鲴等)和喜好流水底栖型鱼类(鳗鲡、子陵吻鰕鯱鱼、蛇鮈等)的生存,增加了区域内鱼类功能性状结构复杂度,导致该区域功能多样性的上升。 总体而言,柘林水库部分库区物种多样性水平较低且生态系统抗干扰能力相对较弱,保护生物多样性和维护生态系统的稳定性应该寻求如何提高鱼类群落的冗余度以及资源和生境的可用性。首先,确定脆弱的鱼类功能群及其冗余水平,并突出哪些物种需要维持功能组分是实现这些目标的关键;其次,恢复特定功能类群鱼类的种群数量、调控鱼类种群结构、增加鱼类群落冗余度,使得生态位空间被充分的填充利用,达到提高生态系统的抗干扰能力的目的。2 结果与分析
2.1 鱼类物种组成

2.2 鱼类优势种组成

2.3 物种组成时空格局变化
2.4 物种多样性指数变化
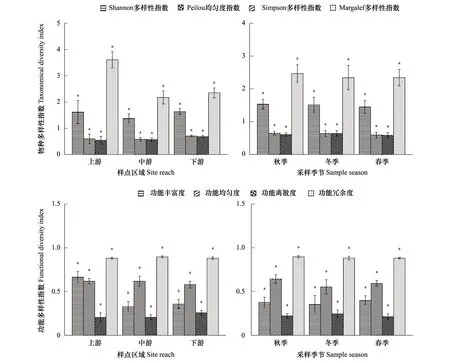
2.5 物种多样性与功能多样性关系
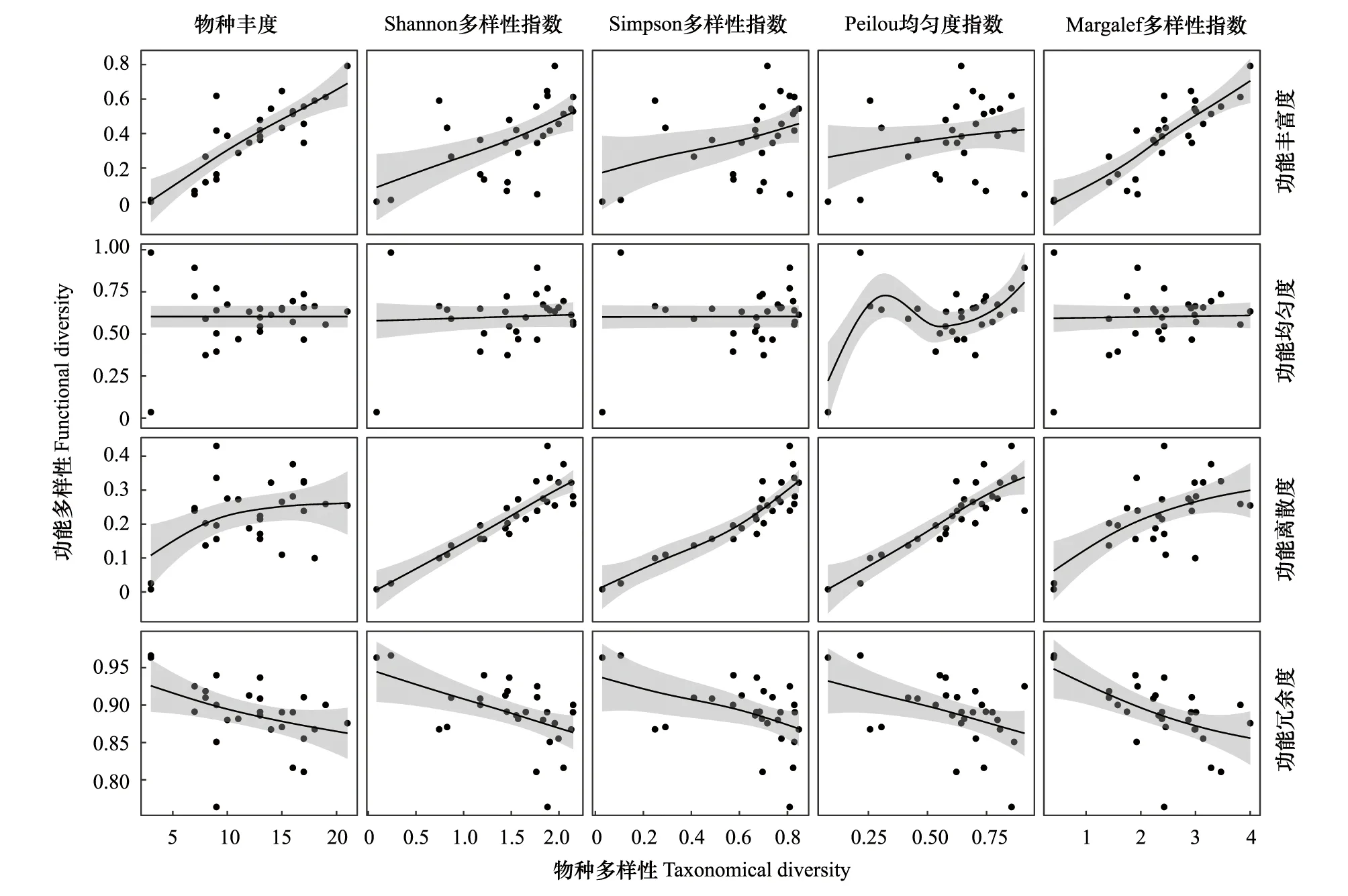
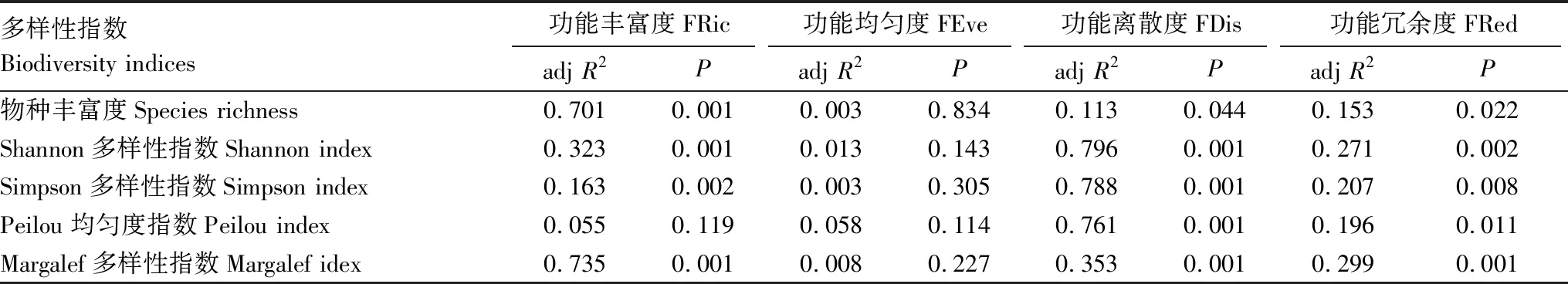
3 讨论
3.1 鱼类群落结构变化
3.2 生物多样性时空变化
3.3 功能多样性与物种多样性指数关系
猜你喜欢
当代水产(2022年8期)2022-09-20
湖北植保(2022年4期)2022-08-23
海洋信息技术与应用(2022年1期)2022-06-05
昆明医科大学学报(2022年2期)2022-03-29
食品安全导刊(2021年20期)2021-08-30
土壤(2021年1期)2021-03-23
儿童时代·幸福宝宝(2020年9期)2020-09-08
河南科学(2020年3期)2020-06-02
福建农业学报(2016年6期)2016-11-01
