
柴达木盆地荒漠植物功能性状及其对环境因子的响应
2022-06-28 08:20:08霍佳璇潘莹萍孟德惠王源源鲁瑞洁黄永梅
生态学报 2022年11期
霍佳璇,任 梁,潘莹萍,赵 瑾,向 响,余 程,孟德惠,王源源,鲁瑞洁,黄永梅
北京师范大学地理科学学部,地表过程与资源生态国家重点实验室,北京 100875
植物功能性状是表征植物生长、发育、繁殖等生命活动的一系列特征,是植物在漫长的进化过程中,由气候、干扰作用和生物条件共同筛选的结果[1],反映了植物对资源的利用和对环境的适应[2]。植物功能性状受到植物本身和环境因子的共同影响,同种植物在不同的环境条件下功能性状值存在差异,不同植物在相同的生境条件下,植物功能性状可能会存在趋同[3]。荒漠广泛分布于南北半球的中低纬度地区,中国的荒漠类型主要为温带荒漠,约占国土面积的1/5[4]。荒漠地区气候干旱、土壤贫瘠,环境条件决定着植物群落的类型、构建和分布,影响植物功能性状,探究荒漠植物功能性状及其与环境因子的关系能够揭示荒漠植物适应干旱环境的不同策略。近年来,关于植物功能性状与环境因子关系的研究大多集中在森林和草地[5—8],针对荒漠植被的研究相对较少,其中大部分只对植物的氮磷元素进行研究和分析,且中国的相关研究多集中在内蒙古[9-10]和新疆部分地区[11—13]。与全球、全国以及区域尺度的研究结果相比,中国北方荒漠地区植物具有相对较高的叶片N和P含量[14],但单独对某一区域进行研究时,植物功能性状的特征和规律又不尽相同,在新疆,植物叶片的碳氮磷含量显著低于中国和其他干旱荒漠地区[12],同样的结果也出现在阿拉善荒漠植物中[10],而对于柴达木盆地荒漠植物功能性状的特征尚不明确。
柴达木盆地位于青藏高原东北部,气候高寒干旱,土壤贫瘠,分布着不同类型的荒漠植被,是研究荒漠植物功能性状的理想场所。20世纪90年代,孙世洲对柴达木盆地及其周边山地的植被类型进行了详细调查和描述[15],之后潘晓玲等对柴达木盆地植物区系进行了区分和探讨[16],牛慧慧等计算并分析了柴达木盆地东部荒漠植物的生态位特征,认为该区域生态位重叠较低,物种竞争小,群落处于一个相对稳定的状态[17]。总体上,针对柴达木盆地植被和群落动态的研究较多,但对柴达木盆地这一特殊地理区域植物功能性状的研究目前还比较缺乏。因此,本研究以柴达木盆地为研究区域,探究荒漠植物功能性状及其对环境因子的响应,拟回答以下问题:1)柴达木盆地荒漠植物功能性状的基本特征;2)影响温带荒漠植物功能性状的主要环境因子有哪些?旨在为该地区荒漠生态系统的保护提供理论依据。
1 材料与方法
1.1 研究区概况
柴达木盆地位于青藏高原东北部(35°30′N—39°30′N、90°00′E—98°31′E),海拔约2600—3000m。本区属于高原大陆性荒漠气候,气候干旱,风力强盛。盆地内年均温在5℃以下,年均降水量由东南(300mm左右)至西北递减(<20mm)[18],年均蒸发量2000—3000mm[17]。本区土壤类型主要为棕钙土、灰棕漠土和盐土,植被类型从东部的荒漠化草原、荒漠,到西部变为极端干旱裸露、基本上无植被的荒漠地带。荒漠植被常见建群种有蒿叶猪毛菜(Salsolaabrotanoides)、合头草(Sympegmaregelii)、梭梭(Haloxylonammodendron)、驼绒藜(Ceratoideslatens)、沙拐枣(Calligonummongolicum)、红砂(Reaumuriasongarica)、多枝柽柳(Tamarixramosissima)等。
1.2 野外调查
于2019年8月对柴达木盆地植物群落和植物功能性状进行调查,样点均选在较为平坦的冲洪积平原或冲洪积扇上,避免坡度和坡向等微地形对植物功能性状的影响。共调查25个样点(图1),包括小乔木荒漠、灌木荒漠、(小)半灌木荒漠3种荒漠植被亚型[4],涵盖多枝柽柳、蒿叶猪毛菜、合头草、红砂、膜果麻黄(Ephedraprzewalskii)、沙拐枣、小果白刺(Nitrariasibirica)、梭梭、驼绒藜等群系。在每个样点根据实际植被覆盖度设置灌木群落样方,面积为5m×5m或者10m×10m,记录样方内所有植物物种名称、每株植物的高度、冠幅,灌木层盖度根据每丛冠幅计算。依据《中国植被》[4]和《中国植物志》[19]将柴达木盆地荒漠植物生活型划分为半灌木小半灌木、灌木、多汁盐柴类灌木和小乔木。具体物种的生活型划分如表1所示。
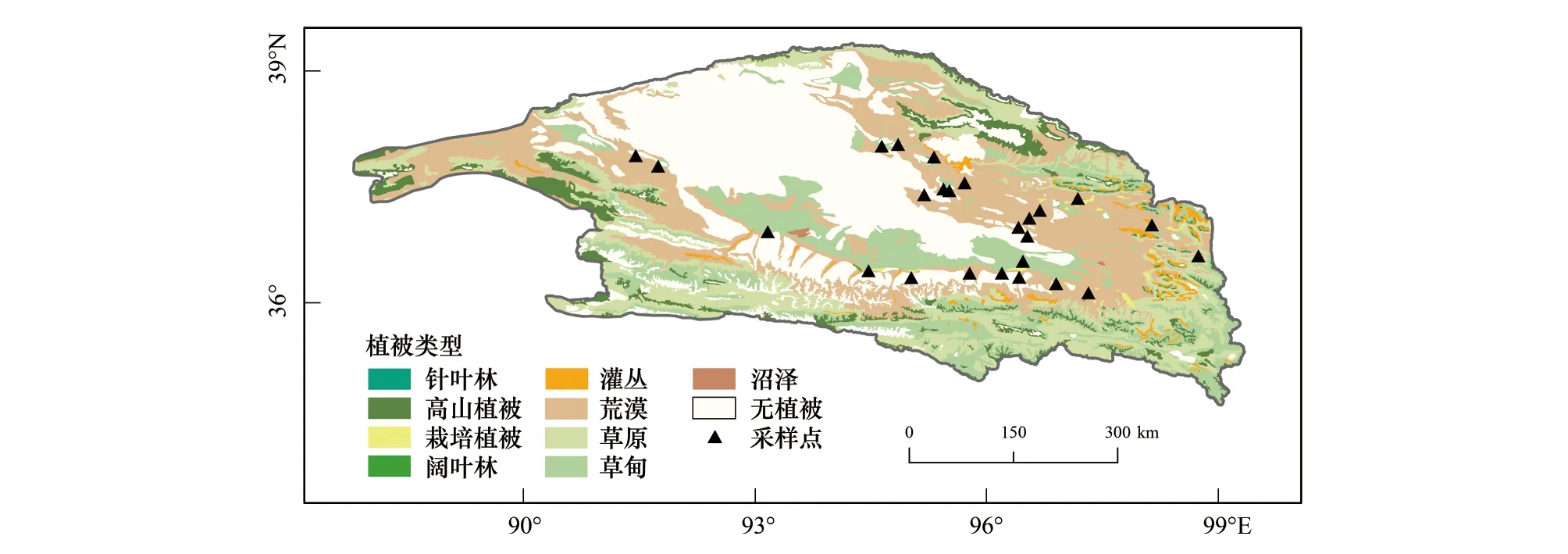
图1 柴达木盆地植被类型及采样点分布图Fig.1 Vegetation and the sampling sites in Qaidam Basin

表1 柴达木盆地荒漠植物生活型划分Table 1 Division of desert plant life forms in Qaidam Basin
1.3 样品采集与测定
选取样方中相对分盖度累积达80%的物种,每个物种随机选取9个植株,测量植株高度,选取10—15株以上植物个体采集完全伸展、没有病虫害的一定数量的叶片(梭梭和沙拐枣为绿色同化枝),带回实验室进行分析测定[20]。新鲜叶片遮光浸泡约12h,取出后迅速吸去表面水分,称重得到叶片饱和鲜重(精度为0.01g),之后在105 ℃下杀青30min,在65℃继续烘干至恒重(约48h),称量得到叶片干重(精度为0.01g),用于叶片干物质含量(leaf dry matter content, LDMC)的计算。另取植物新鲜叶片直接烘干后研磨至过100目筛,叶片碳含量 (Leaf carbon content, LCC)和叶片氮含量 (Leaf nitrogen content, LNC)通过全自动元素分析仪(vario EL cube, Elementar Analysensysteme GmbH, Hanau, Germany) 测定;叶片磷含量 (Leaf phosphorus content, LPC)用电感耦合等离子体原子发射光谱仪(Spector Arcos Eop, SPECTRO Analytical Instruments GmbH, Clive, Germany)测定,叶片稳定碳同位素(δ13C)用稳定同位素比质谱仪 (IRMS, Delta V Advantage; Thermo Fisher Scientific, Inc. USA)测定。
在每个样点采集土壤样品,土壤取样深度为0—20cm。土壤样品自然风干后过筛,砾石和土壤分别称重,计算砾石含量。土壤质地通过Mastersize 3000激光粒度仪测定。土壤容重通过环刀法测定。土壤全碳、全氮含量通过全自动元素分析仪测定;土壤全磷用电感耦合等离子体原子发射光谱仪测定;土壤pH用pH计(pHS- 3C+酸度计)测定;土壤有机碳(TOC, Total organic carbon)含量去除无机碳后通过碳氮元素分析仪(CN 802 Carbon Nitrogen Analyzer, VELP, Italy)测定,土壤无机碳通过测定碳酸钙获得。
本研究采用了全球[21]1280个物种和中国[22]753个物种的叶片氮、磷含量,以及青藏高原[23]74个物种的叶片氮含量数据与柴达木盆地植物功能性状进行比较分析。本研究中采用的气象数据来自中国区域高时空分辨率地面气象要素驱动数据集[24]。
1.4 数据分析
对所有植物功能性状数据进行log10转换使数据符合正态分布。采用单因素方差分析(one-way ANOVA)对不同区域和不同生活型植物性状的差异性进行分析,通过最小显著差异法(LSD)方法进行多重比较。采用冗余分析(RDA,Redundancy analysis)对植物功能性状与环境因子的关系进行分析,与其他植被类型研究结果的比较采用独立样本t检验。数据分析和作图在R 4.0.3中完成。
2 结果
2.1 柴达木盆地植物功能性状基本特征
柴达木盆地荒漠植物功能性状的变化特征如图2所示。柴达木盆地植物LCC、LNC、LPC、叶片C∶N、叶片N∶P、叶片δ13C、LDMC和Height的变化范围分别为272.07—466.00mg/g、 12.40—44.50mg/g、0.51—2.07mg/g、8.62—29.74、11.37—39.29、-27.38‰—-14.80‰、100—480mg/g、3.00—264.22cm。柴达木盆地植物LNC高于中国和全球植物的LNC,低于青藏高原植物的LNC;LPC低于全球和中国的LPC;植物叶片N∶P高于全球和中国的叶片N∶P,但均不存在显著差异。
方差分析结果表明,LNC、LPC、叶片N∶P和LDMC在不同生活型间差异不显著,LCC、叶片C∶N、叶片δ13C和植株高度在不同生活型间存在显著差异(图2)。灌木LCC(426.77mg/g)显著高于小乔木(356.62mg/g)和半灌木小半灌木(343.86mg/g),叶片C∶N也呈现出同样的结果。小乔木和灌木生活型植物的叶片δ13C显著高于半灌木小半灌木(-25.25‰)与多汁盐柴类灌木(-25.90‰)。四种植物生活型中,半灌木小半灌木生活型的植株高度最低(23.20cm),其他三类差别不大。

图2 柴达木盆地荒漠植物功能性状基本特征Fig.2 Characteristics of functional traits of desert plants in Qaidam Basin不同小写字母表示同一植物功能性状在不同研究区域生活型之间存在显著差异(P<0.05), ns表示无差异
2.2 环境因子对植物功能性状的影响
RDA分析结果如图3所示,第Ⅰ轴和第Ⅱ轴的解释率分别为44.72%和35.54%,两轴的解释率相加达到80.26%。第Ⅰ轴主要体现了年均温、年均降水量、砂粒含量和土壤pH值对植物功能性状的影响,第Ⅱ轴主要体现了土壤砾石含量、土壤容重、土壤有机碳、土壤无机碳和土壤全氮含量对植物功能性状的影响。沿第Ⅰ轴年均温与LCC、LDMC、叶片δ13C和植株高度的夹角小于90°,而年均降水量与这些性状的夹角大于90°,表明柴达木盆地植株高度、LDMC、LCC和叶片δ13C随着年均温的增加而增加,随年均降雨量的增加则减小。沿第Ⅱ轴影响较大的土壤砾石含量和无机碳含量与叶片N∶P正相关,与LNC和LPC呈负相关关系。沿第Ⅰ轴可以区分出半灌木小半灌木、小乔木和灌木3种生活型,排序图表明半灌木小半灌木生长于降水量相对较高的地方,随着温度的增加和降水量的减少,植物生活型逐渐向小乔木、灌木等更加耐干旱的植物生活型转变。
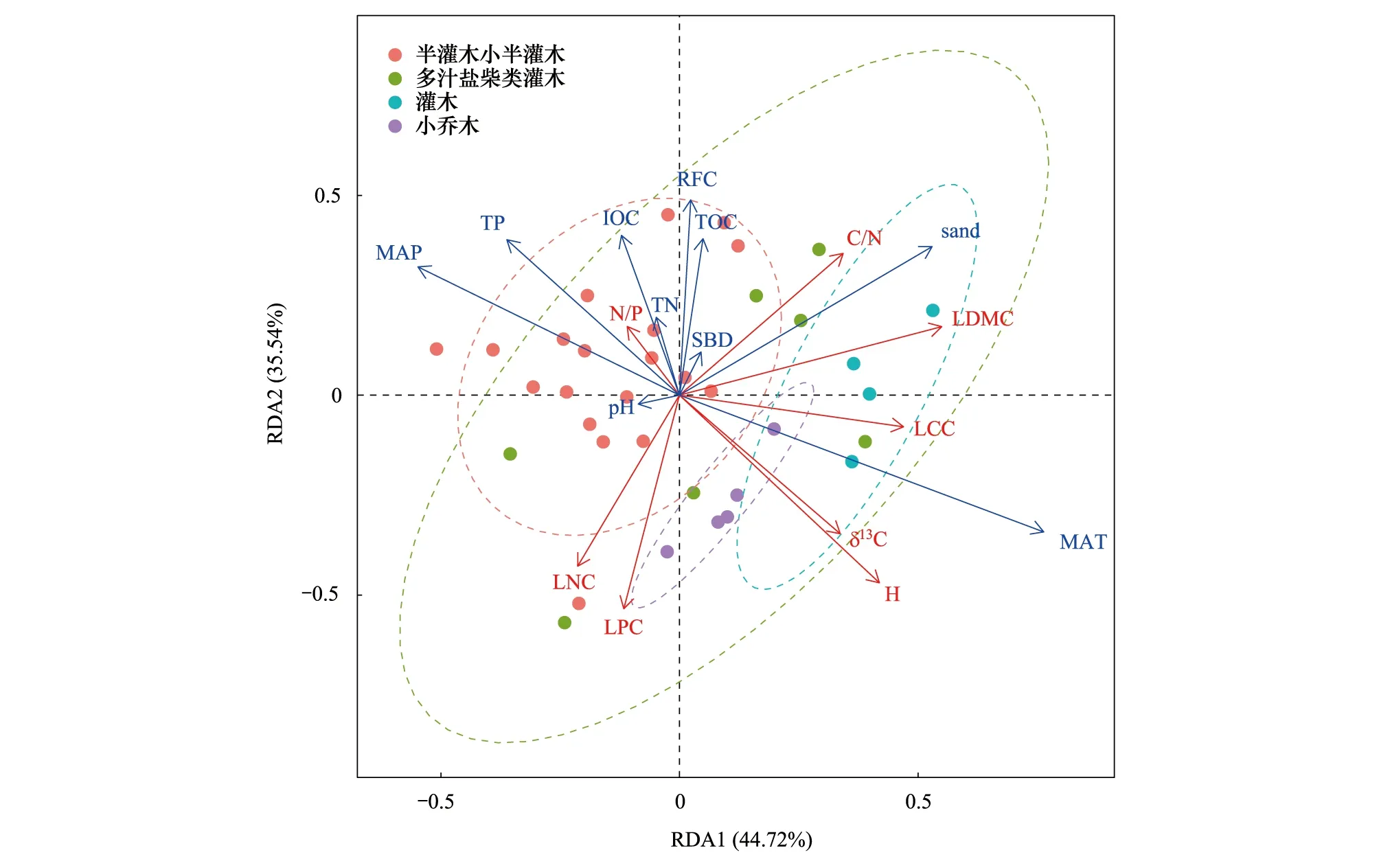
图3 柴达木荒漠植物功能性状与环境因子的RDA分析Fig.3 RDA of plant functional traits and environmental factors in Qaidam BasinMAT:年均温;MAP:年均降水量;RFC∶砾石含量;SBD:土壤容重;sand:砂粒含量;TOC∶有机碳;IOC∶无机碳;TN∶土壤总氮;TP:土壤总磷;pH:土壤pH值;LDMC∶叶片干物质含量;H:植株高度;LCC∶叶片碳含量;LNC∶叶片氮含量;LPC∶叶片磷含量;δ13C:叶片稳定碳同位素;C/N∶叶片碳/氮;N/P:叶片氮/磷
3 讨论
3.1 柴达木盆地荒漠植物功能性状的基本特征
柴达木盆地灌木的LNC和LPC小于小乔木,而LCC大于小乔木,这与浙江天童的研究结果一致[25],但LNC和LPC的差异不显著,仅LCC差异显著,此外,LNC和LPC在所有生活型间的差异也不显著,特别是磷的含量和变化范围都较小。一方面,柴达木盆地气候土壤条件较一致,土壤中氮磷元素含量的范围变化并不大,尽管降水量最小值与最大值相差较多,但总体上仍属于干旱区,植物也以适应干旱生境的旱生、超旱生植物为主[26-27];另一方面氮元素和磷元素在植物叶片中的含量较稳定[28],因此不同的植物生活型之间LNC和LPC没有明显的差异。叶片δ13C通常用来表征植物的水分利用效率[29],本研究中小乔木和灌木的叶片δ13C显著大于多汁盐柴类灌木和半灌木小半灌木,原因是小乔木中的梭梭和灌木中的沙拐枣为C4植物,通常C4植物叶片δ13C高于C3植物并且具有较高的水分利用效率[29-30],更加适应干旱的生境[31]。四种生活型中,半灌木小半灌木的植株高度最低,其他三类没有显著差别,这主要是由植物自身的遗传特性决定。总体来看,尽管个别性状在不同生活型之间存在差异,但植物性状的差异并不大,这可能也是环境对植物进行长期自然选择的结果,植物在这一过程中逐渐形成了稳定可遗传的适应干旱生境的功能性状[11,32]。
柴达木盆地荒漠植物LNC、LPC和叶片N∶P与全球和中国相比不存在显著差异,研究表明叶片中的氮、磷元素变异较小,对于非生物环境的变化不敏感,叶片氮磷浓度相对稳定,即叶片养分浓度稳定假说[28, 33]。与中国其他植被类型相比柴达木盆地荒漠植物功能性状存在显著差异,LDMC显著低于中国东部森林[34];LCC显著低于中国东部森林[35]、中国灌木[36]和中国草地[37]。与中国草地植物LNC相比[37],柴达木盆地荒漠植物的LNC显著较低,但显著高于中国东部森林[35];LPC显著低于中国东部森林[35]和中国草地[38],叶片N∶P显著高于中国东部森林[35](表2)。

表2 柴达木盆地荒漠植物功能性状与其他植被类型的比较Table 2 Comparison of functional traits between desert plants and other vegetation types
LDMC能够反映植物的资源获取能力和叶片的组织构建,高LDMC的叶片通常相对坚硬,更能抵御物理伤害,而肉质植物的LDMC含量一般较低[20, 39],这可能也是本研究中植物LDMC显著低于中国东部森林的原因。尽管普遍认为LCC在不同生活型中为乔木>灌木>草本,但柴达木盆地荒漠植物的LCC显著小于其他植被类型,在阿拉善荒漠植物的研究中LCC也相对较低[10]。干旱条件下,植物会通过减少地上生物量的分配、叶片退化等形式减少水分的蒸发,并把更多光合作用的产物投资到地下部分和用于抵御外界恶劣环境的器官或组织上。另外荒漠植物LCC含量较低,也可能是因为植物灰分所占比例较大[40-41]。柴达木盆地土壤含盐量较高导致植物体内积累较多的矿物质,从而导致灰分含量较高,LCC较低。N∶P用来指示植物受氮限制还是磷限制[42],本研究中4种生活型植物叶片的N∶P均大于16,表明柴达木盆地的荒漠植物主要受P的限制,这与新疆67种荒漠植物的研究结果一致[12]。李玉霖等的研究也表明干旱区荒漠生态系统的植物具有较低的叶片P含量和较高的叶片N/P[14]。LNC显著高于中国东部森林,但显著低于中国草地,豆科植物在草地植物中所占比例较大,豆科植物的根系与土壤中的根瘤菌共同作用可以将大气中的N2转化为自身可利用的氮元素,这可能是草地植物总体LNC较高的原因。然而,柴达木盆地土壤中氮含量较低,LNC却仅次于草地植物,温度-植物生理假说认为植物较高的叶氮含量可以维持低温下正常的生理过程[21],柴达木盆地属于高寒荒漠类型,年均温仅3.7℃左右,这可能是植物为了适应高寒干旱的气候条件所采取的生存策略。LPC主要来源于土壤,而土壤磷主要来源于岩石风化[43],荒漠地区降水稀少、气候干旱不利于岩石的风化,由此造成的土壤磷含量较低,从来源上减少了植物可获取的磷元素。此外有研究发现石灰性土壤中的碳酸钙存在固磷作用,而柴达木盆地土壤碳酸钙含量较高,这阻碍了植物对磷的吸收[44-45]。有研究表明当土壤pH值在6.5—7.5之间时土壤中磷的有效性最高[46],而柴达木盆地土壤pH均大于7.5,这也可能降低了土壤中磷的可利用性。以上因素可能共同导致了柴达木盆地植物叶片较低的磷含量,并且可能存在磷限制。
3.2 环境因子对植物功能性状的影响及其生存策略
本研究结果表明影响柴达木盆地荒漠植物功能性状的环境因子主要是MAT、土壤砂粒含量和土壤砾石含量,代表了气候的干旱程度、土壤的保水能力和养分状况(图3)。温度和降水是决定区域气候类型的重要条件,而土壤砾石含量和土壤质地决定了土壤的水分和养分条件。随着温度的升高,LDMC、LCC和植株高度增加,同时更加适应干旱条件的C4植物增多[47],引起了叶片δ13C值的升高。Niinemets Ü发现比叶重随降水的减少和太阳辐射的增加而增加[48],这意味着LDMC与气候的干旱程度可能也存在一定的正相关关系。在干旱的环境中,植物LDMC较高而LNC低,这可以维持植物在严酷环境下的光合作用,减少水分损失[49]。一般认为缓慢投资-收益型物种叶片寿命长、比叶重大,光合能力弱、叶片氮磷含量低;相反的,快速投资-收益的物种叶氮含量高、光合速率和呼吸速率大,但叶寿命短、比叶重小[50-51]。高LDMC、LCC和植株高度可能是荒漠植物在资源获取时采取的慢速投资-收益策略,通过RDA分析我们可以得出灌木和小乔木采取的策略为缓慢投资-收益型,其大部分光合产物用于构建自身组织结构,具有较高的LDMC和LCC。这类植物生长缓慢,但它们的性状组合有利于储存养分, 以适应资源限制和不利环境,采取的生态策略通常为保守型[52]。尽管在图3中半灌木小半灌木生活型与灌木和小乔木在第一轴上处于相反的位置,但我们并不能认为半灌木小半灌木生活型就是快速投资-收益型植物,因为相对于灌木和小乔木来说,半灌木小半灌木可能会更快速的利用和获取资源,但其仍然属于荒漠植物类型,与其他植被类型如草地、森林相比可能依旧是慢速投资-收益型植物。此外,半灌木小半灌木生活型在图3中的分布也较为分散,之后可能还需增加样本量进行进一步的研究和分析。
生态位能够反映植物在生态系统中的时间、空间位置,同时代表了物种的功能地位,环境的影响使不同的物种占据不同的功能位置,生态位出现重叠和分化[53]。Ordoez等[54]认为气候对植物生长形态的影响要大于对叶片性状的影响,即气候条件更多地决定了植物的生活型,这也是为什么沿着RDA的第一轴出现生态位分化(图3)。沿第一轴水热条件梯度可以很好的区分半灌木小半灌木、灌木和小乔木三种生活型,而本次调查中多汁盐柴类灌木仅有多枝柽柳和小果白刺两种,因其主要分布于绿洲和水盐条件较好的盐碱地或沙地,属于隐域型植被,因此没有很明显的与其他生活型区分开。此外,柴达木盆地不同地貌部位也会导致土壤水分条件存在差异,在冲洪积扇的中上部,地下水埋深较深,主要群落类型以旱生灌木、半灌木荒漠为主,而在冲洪积扇的前缘,由于地下水埋深较浅,土壤含盐量增加,多分布多汁盐柴类灌木,如多枝柽柳和小果白刺[4]。通过环境筛选,荒漠区不同地貌部位会分布不同植物群系,从而造成植物功能性状的差异。
4 结论
通过分析柴达木盆地荒漠植物功能性状的特征及其对环境因子的响应,表明影响柴达木盆地荒漠植物功能性状的主要环境因子为温度,其次是土壤砂粒含量和砾石含量,土壤碳、氮、磷含量与植物叶片碳、氮、磷含量关系不明显。柴达木盆地荒漠植物通过采取慢速投资-收益的资源获取策略,以适应资源限制和抵御恶劣的生存环境。不同生活型的荒漠植物存在较明显的生态位分化,以此来适应不同程度的干旱环境,这是环境选择和植物进化共同作用的结果。
猜你喜欢
凯里学院学报(2024年3期)2024-07-17 00:00:00
小哥白尼(趣味科学)(2022年5期)2022-08-15 08:34:46
中国石化(2021年8期)2021-11-05 07:00:14
矿产勘查(2020年6期)2020-12-25 02:41:08
绿色中国(2019年14期)2019-11-26 07:11:34
沉积与特提斯地质(2019年4期)2019-07-19 06:04:20
成才之路(2017年21期)2017-07-27 20:52:40
现代农业科技(2016年22期)2017-03-24 12:21:57
文史春秋(2016年3期)2016-12-01 05:42:20
学周刊·下旬刊(2015年10期)2015-07-22 12:36:17
