
马尾松PmHMGS基因的克隆与表达1)
2022-06-24 08:13:38吴泽英
东北林业大学学报 2022年5期
吴泽英
(山地植物资源保护与种质创新教育部重点实验室(贵州大学),贵阳,550025)
李正春 文晓鹏
(贵州省森林资源与环境研究中心(贵州大学))(山地植物资源保护与种质创新教育部重点实验室(贵州大学))
马尾松(PinusmassonianaLamb)为松科松属高大乔木,是我国特有、植被面积最大的产脂树种[1]。松脂是一类萜类混合物,由松节油和松香组成,可作为生物燃料、医药、化妆品和食品工业的原料[2]。萜类化合物组成的基本单位是异戊二烯(C5),根据C5数目可将萜类化合物分为单萜(C10)、倍半萜(C15)、二萜(C20)等,松节油的主要成分是单萜和倍半萜化合物,松香的主要成分是树脂酸,是一类二萜化合物的异构体混合物[3-4]。松脂富含的萜类化合物主要由丙酮酸途径(MEP)和甲羟戊酸途径(MVA)合成[5]。
3-羟基-3甲基戊二酰辅酶A合酶(HMGS)是MVA途径的关键酶,能催化1分子乙酰辅酶A与1分子乙酰乙酰辅酶A缩合成3-羟基-3甲基戊二酰CoA,随后在羟甲基戊二酰辅酶A还原酶(HMGR)的催化下生成甲羟戊酸(MVA),MVA经一系列酶的磷酸化和脱羧作用生成异戊烯基焦磷酸(IPP),从而为单萜、半萜和二萜等化合物的合成提供通用前体[5]。如今HMGS基因已在许多植物被分离和鉴定,如思茅松[3]、竹叶花椒[6]、银杏[7]和檀香[8]等。关于马尾松HMGS基因,有报道其表达量与松脂产量有相关性[2],但马尾松参与萜类生物合成的HMGS基因未被鉴定。基于马尾松HMGS基因未被克隆鉴定,本研究首次克隆马尾松PmHMGS的cDNA全长序列,对序列进行生物信息学分析,并采用实时荧光定量检测PmHMGS在幼年树不同组织、机械损伤和茉莉酸甲酯处理不同时间下的表达情况,以期为进一步探讨马尾松PmHMGS在萜类代谢途径中的分子调控机制提供基础理论参考。
1 材料与方法
1.1 试验材料及处理
试验植物材料为1.5年生和2年生的马尾松实生苗。种子采自贵州都匀马尾松国家林木良种基地,在贵州大学农业生物工程研究院实验室培育而得。
茉莉酸甲酯(MeJA)处理:选健康长势一致的1.5年生马尾松苗,参照文献[9]的方法配置MeJA溶液和处理苗,用10 mmol/L的MeJA溶液分别喷施苗0、6、24、48 h;机械损伤处理为2年生长势一致的苗,在地上部10 cm处茎段切割长2 cm,宽为1/2茎环的伤口,分别处理0、2、6、12、24、48 h。每个处理3个生物学重复,在相应的时间点取全木质化的茎,液氮速冻,冰箱中-80 ℃保存。组织特异性分析取材:选3株生长良好一致的1.5年生马尾松苗,取成熟叶、全木质化茎和根,液氮速冻,冰箱中-80 ℃保存。
1.2 马尾松RNA提取与cDNA合成
参照艾德莱RN38总RNA提取试剂盒说明书,提取根、茎、叶和不同处理下茎的总RNA。用酶标仪(Thermo Scientific,USA)检测RNA的浓度。通过1%琼脂糖凝胶电泳检验RNA的纯度和完整性。选择完整且浓度达标的RNA,用TakaRa公司反转录试剂进行反转录,合成cDNA第一链。用内参基因SKI质检cDNA,合格的cDNA稀释5倍后存放于-20 ℃备用。
1.3 基因的cDNA全长克隆
基于马尾松高、低产脂转录组数据的HMGS序列,用Primer Primer5.0设计特异性克隆引物PmHMGS-F和PmHMGS-R(表1),以茎的cDNA为模板,用PrimeSTAR高保真聚合酶进行扩增得到PmHMGS基因的cDNA片段。PCR反应体系为:5 μL的PrimeSTAR,3.5 μL的ddH2O,1 μL的cDNA,0.5 μL的正反引物。反应程序为:98 ℃变性10 s,60 ℃退火10 s,72 ℃延伸30 s,35个循环,4 ℃保存。经1%琼脂糖凝胶电泳检测和胶回收PCR产物。将目的片段链接到pEASY-Blunt克隆载体上,经菌落PCR检测,挑取阳性单菌落送生物工程(上海)股份有限公司测序。测序获得的序列经拼接、比对分析,得到PmHMGS基因的cDNA全长序列。
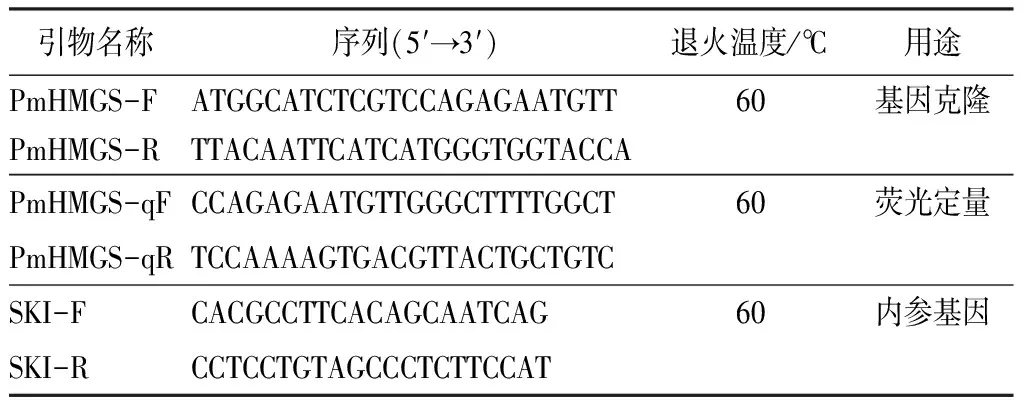
表1 PmHMGS基因的RT-PCR和qRT-PCR引物
1.4 基因的生物信息学分析
将克隆得到的PmHMGS基因序列提交到NCBI的ORF Finder预测开放阅读框,获得蛋白序列;通过ProtParam、TMHMM 2.0、SignalP4.1 Server、PSORT、SOPMA、SWISS-MODEL、CLUSTLWZA、ENDscript/ESPript在线网站和MAGA 7.0本地软件分析PmHMGS蛋白质的理化性质、跨膜结构域、信号肽、亚细胞定位、二级结构、三级结构、同源性比较和构建系统发育树。
1.5 基因的表达分析
根据PmHMGS基因序列,用Primer5.0设计荧光引物PmHMGS-qF和PmHMGS-qR,见表1。以SKI为内参基因,通过qRT-PCR方法检测PmHMGS基因在不同组织、不同处理下的表达情况。反应程序:95 ℃预变性30 s,95 ℃变性15 s,60 ℃退火15 s,72 ℃延伸20 s,共35个循环。3个技术重复和3个生物学重复。
1.6 数据处理
用算法2-ΔCt计算基因的相对表达量,Excel2016处理数据,SPSS 22.0软件分析数据的显著性。
2 结果与分析
2.1 PmHMGS基因的cDNA全长克隆
通过PCR扩增PmHMGS基因,产物如图1所示,扩增条带在1 500 bp左右,条带位置、大小符合预期,可分离纯化产物送公司进行测序。
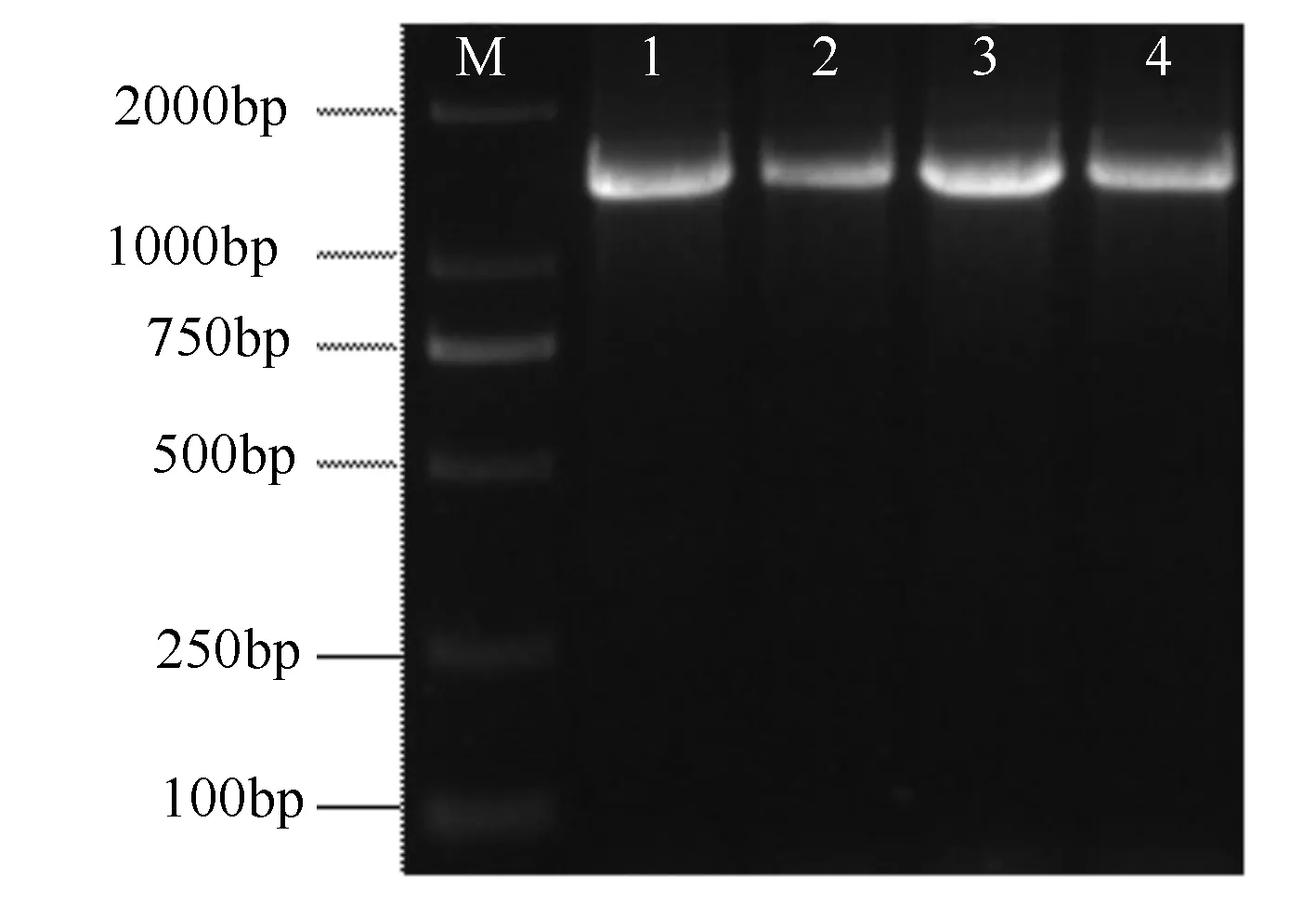
M为Marker;1、2、3、4为PmHMGS的cDNA全长。
2.2 PmHMGS基因的生物信息学分析
通过ORF Finder网站分析显示,PmHMGS基因的ORF为1 425 bp,编码474个氨基酸,与转录组HMGS基因序列的ORF一致,命名为马尾松3-羟基-3甲基戊二酰辅酶A合酶基因(PmHMGS)。
ProtParam在线软件预测PmHMGS蛋白的理化性质显示,PmHMGS蛋白的分子式为C2357H3656N618O724S23;相对分子质量为52 972.04,理论等电点为5.61。带负电氨基酸残基总数为59,带正电氨基酸残基总数为50。不稳定系数为44.34,表明PmHMGS为不稳定蛋白质。其脂肪族氨基酸指数为76.98,总的亲疏水性平均系数为-0.28,表明该蛋白质为亲水性蛋白。
SignalP4.1 Server分析显示PmHMGS蛋白不含信号肽;THMM2.0在线工具对蛋白序列跨膜结构进行分析,发现PmHMGS未形成跨膜结构域;利用Cell-PLoc 2.0在线软件进行亚细胞定位预测,结果表明该蛋白定位于细胞质。萜类代谢MVA途径存在于细胞质中,这3种预测结果可以推断马尾松PmHMGS酶在细胞质中合成后,不进行蛋白转运,而是留在细胞质中直接与代谢底物作用,参与萜类化合物的合成。
利用Pfam和NCBI的CD Search网站分析PmHMGS蛋白的保守结构域,CD Search结果显示该蛋白含典型的HMGS活性结构域(图2),Pfamh网站预测到该蛋白保守结构域的N末端位于第4~176位点,C末端为第178~452位点。
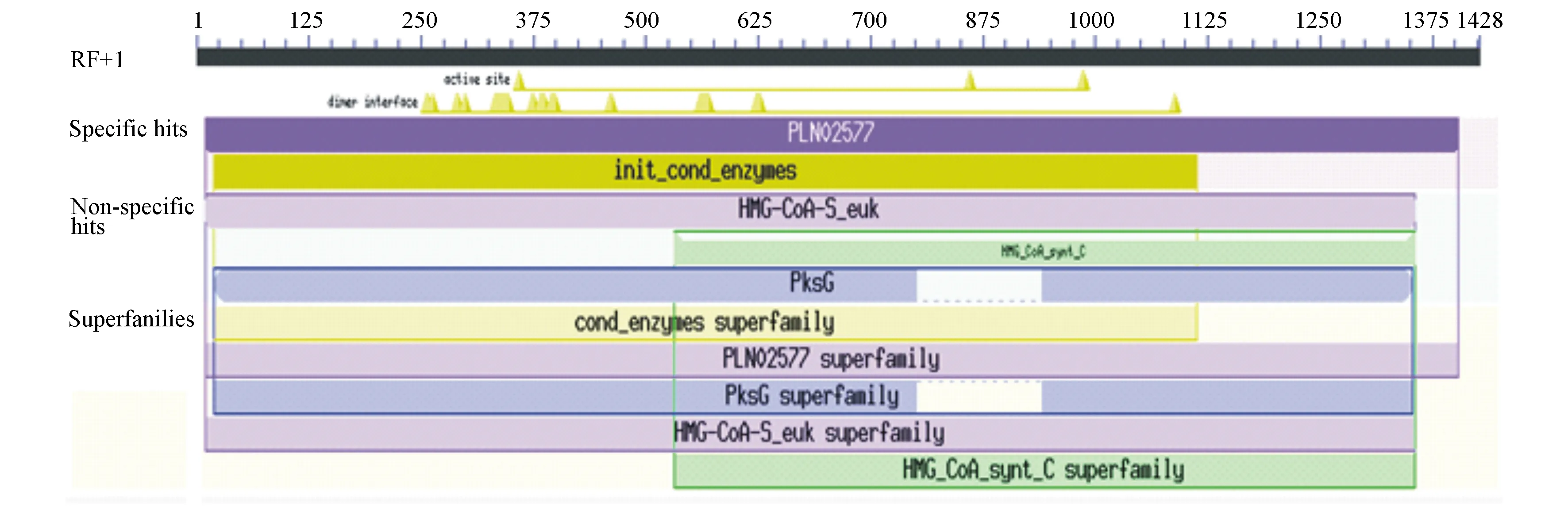
图2 马尾松PmHMGS蛋白的保守结构域预测
应用SOPM在线网站预测马尾松PmHMGS蛋白的二级结构,结果表明该蛋白二级结构中有46.20%的氨基酸残基为α-螺旋(H),13.08%的氨基酸残基为β-螺旋(E),4.64%的氨基酸残基为β-转角(T),36.08%的氨基酸残基为无规则卷曲(C)。用SWISS-MODEL预测该蛋白的三维结构(图3),以拟南芥3-羟基-3-甲基戊二酰CoA合酶(PDB NO:2fa3.1.A)的三维结构为模板来建模,序列相似性为74.33%。
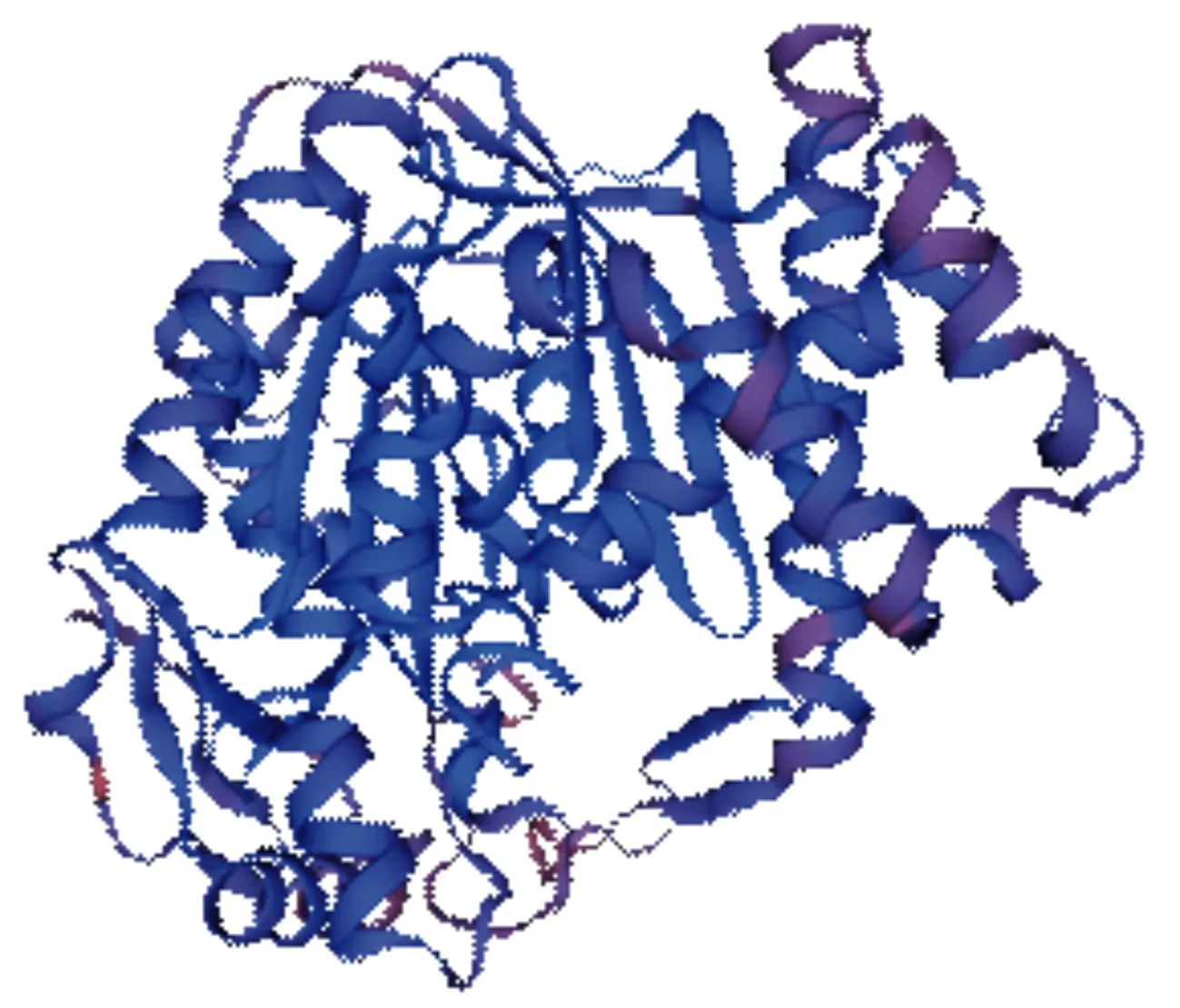
图3 马尾松PmHMGS蛋白三级结构的预测
将PmHMGS蛋白序列放在NCBI的Blast中检索,下载同源性较高的不同物种序列,用在线软件CLUSTLWZA比对分析,ENDscript/ESPript对分析结果作图。结果显示(图4),马尾松PmHMGS蛋白序列与樟子松的PsHMGS蛋白序列同源性为99.58%,与思茅松的PkHMGS蛋白序列同源性为99.37%,与曼地亚红豆杉的TmHMGS蛋白序列同源性为84.18%,与银杏GbHMGS蛋白序列同源性为81.95%。从图中可知,马尾松PmHMGS蛋白与其他已知功能植物的HMGS蛋白功能结构域几乎一致,有活性中心“NXD/NE/vEGI/vDx(2)NACF/yxG”保守基序和5个参与底物催化的保守活性氨基酸位点,分别为Glu86、Cys120、Ser251、Gly328和Ser362[7]。这一结果表明HMGS蛋白的功能结构域在分子进化中具有较高的稳定性和保守性。
用MEGA7.0软件对马尾松、樟子松、思茅松等15种植物的HMGS蛋白序列构建系统发育树,研究HMGS蛋白之间的序列多样性和进化关系。结果如图5所示,马尾松的PmHMGS蛋白与樟子松、思茅松、曼地亚红豆杉和银杏的HMGS蛋白序列聚为一支,它们都属于裸子植物。其中马尾松HMGS与樟子松HMGS亲缘关系最近,其次是思茅松,这与植物的物种分类一致,反映了植物HMGS的进化保守性和进化多样性。
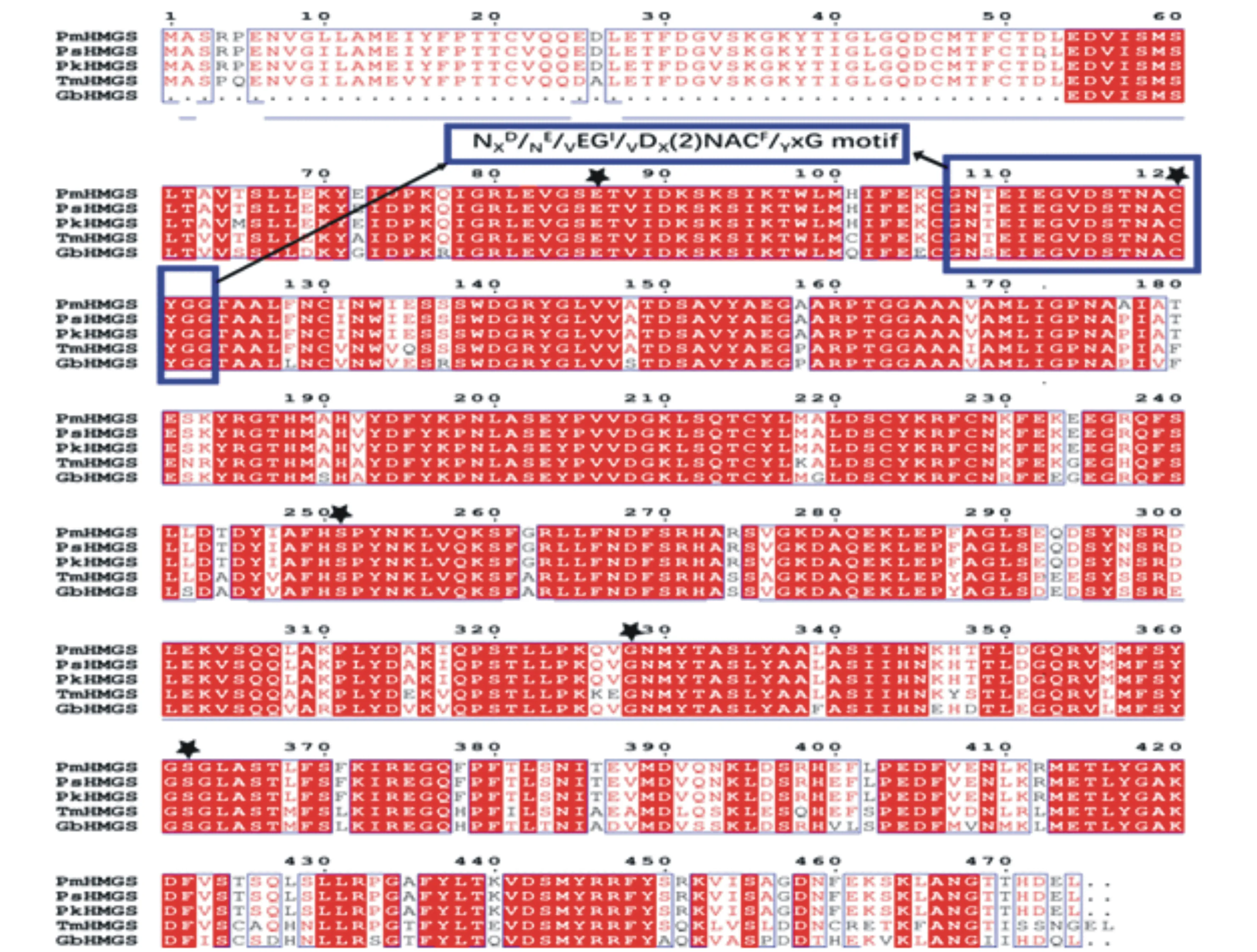
蓝色方框为PmHMGS蛋白的活性中心;黑色五角星为HMGS蛋白活性位点;PsHMGS为樟子松蛋白序列(CAA65250.1);PkHMGS为思茅松蛋白序列(QLL99514.1);TmHMGS为曼地亚红豆杉蛋白序列(AAT73206.1);GbHMGS为银杏蛋白序列(AWG97032.1)。
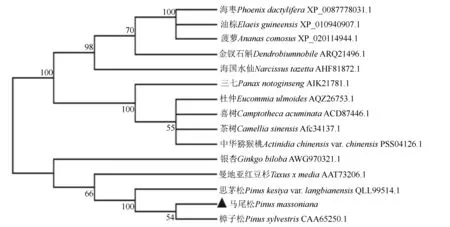
图5 马尾松PmHMGS蛋白的系统进化树
2.3 PmHMGS基因的表达分析
利用qRT-PCR技术分析PmHMGS基因在马尾松不同组织、茉莉酸甲酯和机械损伤处理不同时间下茎的表达情况,结果显示(表2~表4),PmHMGS基因在根、茎、叶中有差异表达,在茎中表达量最高,其次为根和叶,两者表达量大致相同。茉莉酸甲酯处理后,马尾松茎中PmHMGS的表达量随着MeJA处理时间增加而增加,在处理48 h时,PmHMGS仍然响应的MeJA的诱导表达,是未处理的5.6倍。机械损伤处理后,PmHMGS基因在茎中的相对表达先升后降,在12 h时达到最大值,是未处理的10.17倍。
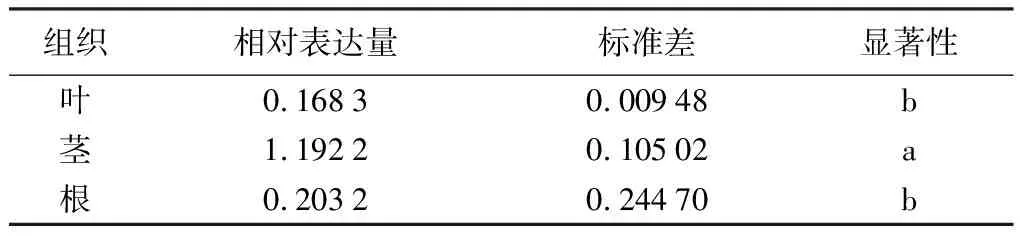
表2 PmHMGS基因在叶、茎、根中的相对表达量
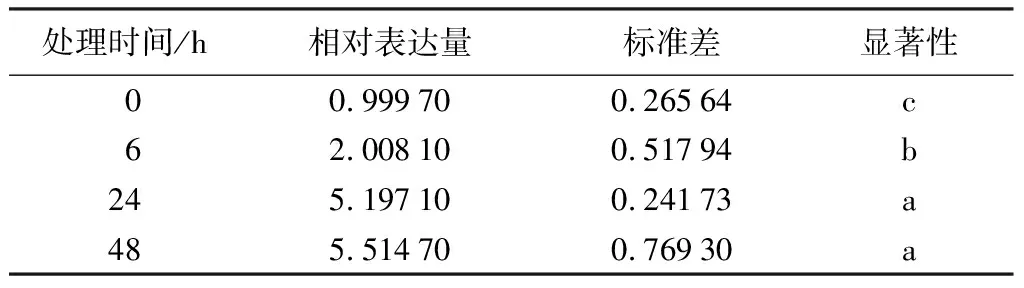
表3 茉莉酸甲酯处理不同时间下PmHMGS基因在茎中的相对表达量
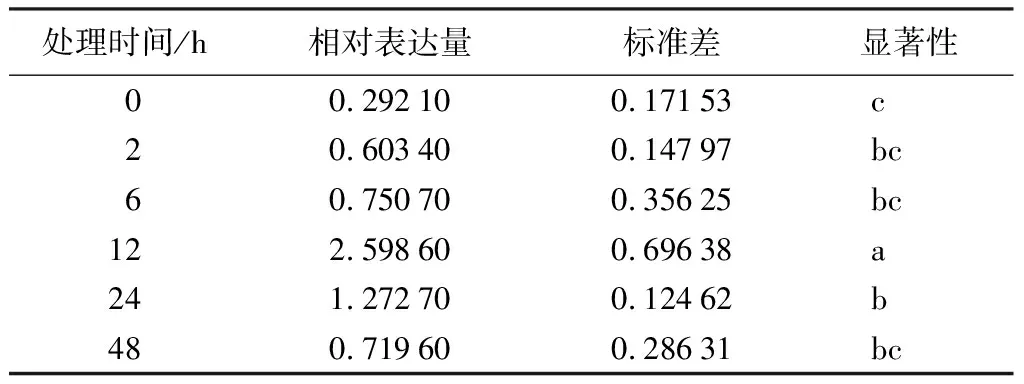
表4 机械损伤处理不同时间下PmHMGS基因在茎中的相对表达量
3 讨论与结论
松脂产量是一个高度遗传的性状,通过传统的育种方法可以获得大量的遗传收益,但传统方法过程十分缓慢[10]。另外,马尾松产脂性状还受生物、非生物胁迫和气候等因素的影响,让传统育种工序变得更繁琐[11]。目前随着转录组测序技术的发展,挖掘和研究高产脂相关的基因,从基因工程的角度来获得高产脂品种或创造高产脂种质马尾松的研究也越来越成为研究热点[12]。
HMGS是MVA途径中萜类生物合成类异戊二烯的限速酶,类异戊二烯是所有萜的C5前体[13]。目前HMGS已在许多植物被克隆和鉴定,而马尾松HMGS的研究相对空缺。本研究首次从马尾松克隆并鉴定了MVA通路中编码HMGS的PmHMGS基因,且分析了它的组织表达水平。克隆的马尾松HMGS基因的cDNA全长为1 425 bp,命名为PmHMGS。推导的PmHMGS蛋白含有474个氨基酸,蛋白相对分子质量为52 972.04,这与报道的植物HMGS蛋白一般由460到500个氨基酸残基组成,相对分子质量为50 000到60 000一致[14]。保守结构域及多序列分析表明,PmHMGS基因与其他植物HMGS基因具有高度的相似性,含有典型的HMGS蛋白保守结构域,即保守基序“NXD/NE/vEGI/vDx(2)NACF/yxG”和5个活性氨基酸位点“Glu86、Cys120、Ser251、Gly328、Ser362”,暗示PmHMGS可能具有其他植物HMGS的功能[7]。系统进化树分析表明,PmHMGS蛋白与属裸子植物的樟子松、思茅松、曼地亚红豆杉和银杏的HMGS蛋白序列聚为一支,其中PmHMGS与樟子松的PsHMGS亲缘关系最近,表明HMGS在不同植物中具有进化起源的保守性,并表现出氨基酸序列和功能结构域的保守性。
HMGS基因在不同植物中的表达模式存在差异,且萜类化合物的产量与HMGS基因表达的组织特异性相关。如檀香的SaHMGS在根中表达量最高,其次是心材,根和心材是提取精油(倍半萜化合物)的主要部位[8];银杏GbHMGS1在叶和果实高表达,叶和果实是地上器官中生产萜类化合物和保护免受太阳辐射的主要部位[7];洋甘菊McHMGS在花中表达量最高,且花中主要倍半萜的含量显著高于其他组织[13]。本研究对PmHMGS基因组织特异性表达分析发现,在1.5年生马尾松的茎中表达量最高,叶和根的表达量相近,与已报道的马尾松HMGS基因在树干木质部中与松脂产量成正相关,在成熟叶中与松脂产量呈负相关相符[2],推测PmHMGS主要在茎中调控萜类化合物的合成。但与思茅松的PkHMGS的表达有出入,其在针叶中高表达[3]。茉莉酸甲酯是一种小信号分子,在外源作用下可以调控植物的次生代谢[14]。MeJA与萜烯代谢密切相关,如MeJA处理提高了檀香SaHMGS、洋甘菊McHMGS、果香菊CnHMGS的表达量,相应的萜类化合物檀香醇、洋甘菊倍半萜类、果香菊倍半萜类的含量也增加[8,15-16]。本实验发现MeJA处理马尾松后,茎中的PmHMGS基因表达水平显著升高,与檀香SaHMGS在茎中响应MeJA的表达模式相似,说明PmHMGS应答MeJA的诱导表达,从而可能增加马尾松萜类化合物的合成。树脂道是生产、储存和运输松脂的复杂网络,遍布于针叶树的根、茎、针叶和球果中[17]。当松脂管道受机械损伤、昆虫攻击和病原体入侵等非生物或生物刺激破坏,树脂道被切断并释放树脂防御伤害,同时新生分化创伤性树脂道,也正调控或负调控基因的表达,从而激活或抑制酶的活性,影响代谢物的积累[18]。如以下损伤处理后:紫茎泽兰的倍半萜烯合酶基因EaTPS1的表达被抑制,卡迪宁(倍半萜化合物)得以大量积累[19];兴安落叶松防御系统的多酚氧化酶(PAL)和内苯丙氨酸解氨酶(PPO)的活性增强[20];思茅松PkHMGS上调表达,在高、低产脂思茅松高表达分别是损伤处理12 h时和6 h[3]。试验发现PmHMGS也响应机械割伤胁迫,在损伤处理12 h时,PmHMGS的表达量达到最大值,与PkHMGS在高产脂思茅松表达模式相似,但与低产脂的PkHMGS表达不同,说明不同种质、不同产脂力的针叶树,HMGS基因对胁迫刺激信号的感知能力不同。综合表明,马尾松PmHMGS基因可能同上述响应MeJA和机械损伤处理的基因一样,调控萜类代谢物的合成,本研究为后续深入探讨PmHMGS基因在马尾松松脂合成中的功能提供基础理论依据。
猜你喜欢
东北林业大学学报(2021年1期)2021-12-03 19:17:48
东北林业大学学报(2021年1期)2021-01-14 04:26:10
绿色科技(2019年5期)2019-11-29 13:17:37
中成药(2018年11期)2018-11-24 02:57:04
天然产物研究与开发(2018年7期)2018-08-21 02:04:02
石油炼制与化工(2018年6期)2018-03-27 03:18:40
山东林业科技(2017年1期)2017-06-29 07:53:58
中文信息(2017年2期)2017-04-13 18:02:02
广西林业科学(2016年1期)2016-03-20 05:33:00
广西林业科学(2016年1期)2016-03-20 05:32:59
