
狮头鹅MAP2K1基因SNP位点筛查及生物信息学分析
2022-06-24 14:30陈新企周光现王慧芳李立先贾汝敏赵志辉
江苏农业科学 2022年11期
陈新企 周光现 王慧芳 李立先 贾汝敏 赵志辉

















摘要:MAP2K1基因屬于GnRH信号通路且与哺乳动物的卵泡发育紧密相关。为研究MAP2K1基因在狮头鹅上的基因多态性,以200羽狮头鹅为试验对象,利用DNA池结合PCR产物直接测序法,筛选出SNP位点并进行生物信息学分析。检测结果表明,在狮头鹅群体中发现1个SNP位点,为E3-63 A>G,存在3种基因型。生物信息学预测显示,MAP2K1基因突变前后编码氨基酸并未发生改变,属于不稳定不跨膜酸性亲水性蛋白,存在卷曲螺旋结构,无信号肽存在。亚细胞定位于细胞核、细胞质、细胞支架的概率分别为60.9%、30.4%、8.7%,主要在细胞核发挥生物学作用;有34个磷酸化位点,有1个N-糖基化位点,1个保守结构S_TKc,蛋白二级结构主要由α螺旋和无规则卷曲组成。MAP2K1基因突变前后mRNA的最小自由能,质心发生二级结构,含有假结的mRNA二级结构、折叠结构、MaxExpect结果、点-括号结果、最小自由能、mRNA结构的热力学集合、质心结构所形成的山地图发生改变,且自由能上升,结构稳定性变弱。从狮头鹅MAP2K1基因的11个外显子上检测出1个SNP位点,MAP2K1基因突变前后改变mRNA二级结构,为进一步研究MAP2K1基因对狮头鹅产蛋数量提供参考。
关键词:狮头鹅;MAP2K1基因;SNP;多态性;生物信息学
中图分类号:S835.3 文献标志码:A
文章编号:1002-1302(2022)11-0043-10
收稿日期:2021-07-25
基金项目:广东省现代农业产业技术体系创新团队项目(编号:2019KJ137、2020KJ137);狮头鹅品种选育及健康养殖校企合作项目(编号:B15337)。
作者简介:陈新企(1996—),女,广西贺州人,硕士研究生,主要从事动物遗传育种与繁殖研究。E-mail:2476359974@qq.com。
通信作者:贾汝敏,教授,研究生导师,主要从事家禽遗传育种与繁殖研究,E-mail:jiarm@gdou.edu.cn;赵志辉,教授,研究生导师,主要从事动物遗传育种与繁殖研究,E-mail:zhzhao@jlu.edu.cn。
狮头鹅被选为广东省第三届“十大名牌”农产品,生长速度快、耗料少,其营养价值高,亚麻酸含量超过其他肉类并接近人体所需氨基酸的比例,具有很高的经济价值。但狮头鹅年产蛋不足32枚,不足高产鹅的1/4,远不能满足市场需求。丝裂原活化蛋白激酶Ⅰ(mitogen activated protein kinase 1,简称MAP2K1,又称MEK1或MAPKK1),属于GnRH信号通路,是细胞进入DNA合成期(即S期)的关键,激活ERK1/2,ERK1/2进入细胞核后,又激活转录因子形成转录复合物,导致细胞的增殖分化。MAP2K1基因属于MEK家族成员,该家族包括MEK1、MEK2、MEK3、MEK4、MEK5、MEK6、MEK7基因,是MAPK信号通路中的重要成员。MAPK信号通路包括:p38MAPK通路(p38 mitogen activated protein,p38MAPK),c-Jun氨基末端激酶通路/应激激活蛋白(c-Jun-N-terminal kinase,JNK/stress activated protein kinase,SAPK),细胞外信号调节蛋白激酶通路(extra cellular signal-regulated protein kinase,ERK)及细胞外信号调节激酶5(ERK5)通路。MAP2K1基因主要参与ERK途径,被认为是ERK1/2最主要调节因子之一。ERK1/2参与分化细胞的减数分裂,有丝分裂后功能的调控],活化原始卵泡和卵泡的生长,促进卵泡颗粒细胞增殖及类固醇激素合成。ERK1/2差异性调节FSH诱导的卵泡颗粒细胞中孕激素和雌二醇合成。MAP2K1是ERK1/2的上游激酶,为少有的双重特异性蛋白激酶,既为酪氨酸蛋白激酶,又为丝氨酸/苏氨酸蛋白激酶。MAP2K1基因突变导致人各种疾病的发生,例如肝癌、结直肠癌等;导致鼠隐性表型致死,影响胎盘的发育,滋养层细胞不能侵入胎盘迷路;影响猪精子生成,影响公牛精液品质。目前,MAP2K1基因在狮头鹅的研究还未见相关报道。有关狮头鹅最新的研究为基于全基因组重测序技术的狮头鹅Indel标记分析和狮头鹅个体产蛋智能记录系统研制。而关于MAP2K1基因的多态性研究主要与人疾病相关,迄今为止,还没有关于狮头鹅MAP2K1基因的多态性及生物信息学分析,及是否可以作为影响产蛋性能候选基因的报道。因此,研究狮头鹅MAP2K1基因外显子多态性可快速、准确、不受环境条件干扰地揭示MAP2K1基因是否影响狮头鹅的产蛋量。
1 材料与方法
1.1 试验材料
狮头鹅由国家级狮头鹅保种场(广东省饶平县)提供,从1 400羽500日龄繁殖期母鹅中随机挑选200羽健康成年母鹅。在翅下静脉肝素钠抗凝采集血样。Green Taq Mix购自南京诺唯赞生物科技有限公司,HiPure Blood DNA Mini Kit试剂盒购自美基生物公司,DL-2000 Marker购自广州东盛生物科技有限公司。
1.2 基因组DNA提取和DNA池的构建
使用试剂盒进行血液样品DNA提取。经1%凝胶电泳检测DNA完整性,使用Nanodrop检测纯度。每个混池100个DNA样品,每个样品各取 1 μL,然后混合均匀,构成狮头鹅基因组DNA池。
1.3 引物设计与合成
目前,GenBank上未公布鹅MAP2K1基因序列,根据文献发现鹅的基因序列与鸭的基因序列一致性较高,因此采用鸭的基因序列设计相应引物。由表1可知,根据GenBank上已公布的綠头鸭MAP2K1基因序列(登录号XM_027465642.2),采用Primer Premier 5.0对狮头鹅MAP2K1基因的11个外显子(Exon1~Exon11)设计引物。
1.4 PCR扩增及SNP位点鉴定
PCR反应总体系为25.0 μL:Green Taq Mix 12.5 μL,上下游引物各1.0 μL,DNA样品1.0 μL,ddHO 9.5 μL。PCR反应程序:94℃预变性5 min;94℃变性30 s,55℃退火30 s,72℃延伸30 s,35循环;72℃延伸10 min,4℃保存。利用DNAstar中seq man应用查看,找出狮头鹅MAP2K1基因SNP位点;计算SNP位点等位基因频率、基因型频率、纯合度、杂合度、有效等位基因数。使用PIC-CALC和PopGen32分别计算多态信息量(PIC)和哈代-温伯格平衡(Hardy Weinberg's law)。
1.5 狮头鹅MAP2K1蛋白生物信息学分析
GenBank上目前未有鹅MAP2K1蛋白序列,所使用序列为DNA测序结果预测的蛋白序列。利用ExPASy分析理化性质、疏水性、卷曲螺旋结构,利用SignalP4.1 Server对信号肽进行预测,利用TMHMM预测蛋白跨膜结构,利用PSORT Ⅱ预测亚细胞定位,利用SMART预测蛋白结构域,利用NetPhos2.0预测磷酸化,利用NetNGlyc1.0 Server预测糖基化位点,利用SOPMA软件预测蛋白二级结构,利用SWISS-MODEL软件预测三级结构。
1.6 狮头鹅MAP2K1基因mRNA二级结构生物信息学分析
利用RNAfold web server和RNAstructure进行mRNA二级结构分析。
2 结果与分析
2.1 狮头鹅MAP2K1基因PCR扩增
由图1可知,对狮头鹅MAP2K1基因Exon1~Exon11进行扩增,PCR扩增后分别获得472、633、649、252、365、377、384、393、415、419、453 bp的产物,琼脂糖凝胶电泳结果显示条带干净清晰无污染。引物序列大小均与预期一致。
由表2可知,将扩增产物测序后的序列进行同源比对,发现该扩增序列与鸭MAP2K1基因序列的一致性达99%。
2.2 狮头鹅MAP2K1基因混池SNP位点鉴定结果
利用DNASTAR对狮头鹅MAP2K1基因测序结果与GenBank上已公布的绿头鸭MAP2K1基因序列进行对比分析。由图2可知,MAP2K1-3引物上有明显套峰。
2.3 狮头鹅MAP2K1基因个体验证
由图3可知,对引物MAP2K1-3进行200羽狮头鹅单个DNA样品PCR扩增后测序,发现1个SNP位点,有3种基因型。由图4可知,在Exon3的第63碱基处A突变成G,命名为E3-63 A>G。
2.4 基因型频率、等位基因频率及哈代温伯格定律数据分析
对筛选出的1个SNP位点,进行等位基因频率、基因型频率、纯合子、杂合子、有效等位基因数、PIC和Hardy Weinberg's law的计算。由表3可知,E3-63 A>G等位基因A和G的频率分别为19.93%、80.07%,G为优势等位基因,中度多态(PIC=0.268 2),在群体遗传过程中符合Hardy Weinberg's law(P>0.05)平衡状态。
2.5 MAP2K1蛋白生物信息学分析
2.5.1 MAP2K1蛋白的理化性质分析
利用ExPASy(http://web.expasy. org/protparam/)对狮头鹅MAP2K1蛋白突变前后的氨基酸组成、等电点和不稳定系数等理化性质进行预测分析。结果显示,MAP2K1蛋白突变前后编码氨基酸并未发生改变。MAP2K1蛋白分子式为CHNOS,由395个氨基酸组成,其中,亮氨酸占比最高,为9.1%,相对分子量为43 576.09,理论等电点PI为6.19,说明该蛋白为酸性,正电荷的残基总数为46,负电荷的残基总数为50,半衰期为30 h,不稳定系数为48.87,说明该蛋白不稳定,脂肪系数为86.63,亲水性为-0.323,说明该蛋白为亲水性蛋白。
2.5.2 MAP2K1蛋白疏水性分析
利用ProtScale(http://web.expasy.org/protscale/)对编码氨基酸疏水性进行预测。以0分为界,数值越大,疏水性越强,数值越小,亲水性越强。结果显示,在第45位和第47位氨基酸亲水性最强,为-2.956;在第170位和第173位氨基酸疏水性最强,为1.989,平均亲水性为 -0.323,说明该蛋白具有亲水性。
2.5.3 MAP2K1蛋白Coil区分析
利用在线网站(https://embnet.vital-it.ch/software/COILS_form.html),预测Coil区。当不同窗口值大于0.5时,则该位点存在Coil区。由图5可知,在14号窗口 30~48位氨基酸有卷曲结构,第30~43位氨基酸的值为1.000,第44~48位氨基酸的值为0.622。在21号窗口第23~50位氨基酸有卷曲结构,第 23~43位氨基酸的值为0.999,第43~50位氨基酸的值为0.989。28号窗口第23~60位氨基酸有卷曲结构,第23~57位氨基酸的值为0.987,第58~60位氨基酸的值为0.626,说明该蛋白具有卷曲螺旋结构。
2.5.4 MAP2K1蛋白信号肽分析
信号肽是决定新生肽链在细胞中的定位或决定某些氨基酸残基修饰的一些肽段。利用蛋白质序列信号肽分析工具SignalP4.1(http://www.cbs.dtu.dk/services/SignalP-4.1/)對MAP2K1突变前后蛋白进行分析。当3种C、Y、S-score计算结果>0.5时,则该位点存在信号肽。结果显示, 在第6位氨基酸处S最大值为0.108,在第26位氨基酸处C、Y最大值分别为0.119、010,均未超过0.5,说明该蛋白无信号肽存在。
2.5.5 MAP2K1蛋白跨膜结构分析
把得到的氨基酸序列输入TMHMM(http://www.cbs.dtu.dk/services/TMHMM/)在线跨膜运输软件进行预测。结果显示,MAP2K1蛋白无跨膜区域,说明MAP2K1不是跨膜蛋白。
2.5.6 MAP2K1蛋白亚细胞定位分析
亚细胞定位是指某种蛋白或表达产物在细胞内的具体存在部位。通过工具PSORTⅡ(https://psort.hgc.jp/form2.html)预测MAP2K1突变前后编码蛋白的亚细胞定位。结果显示,该蛋白位于细胞核的概率为60.9%、位于细胞质的概率为30.4%、位于细胞支架的概率为8.7%,主要在细胞核发挥生物学作用。
2.5.7 MAP2K1蛋白磷酸化位点分析
利用NetPhos软件(http://www.cbs.dtu.dk/services/NetPhos/)预测,当磷酸化电势大于阈值0.5时,则该位点有可能被磷酸化。结果显示,MAP2K1蛋白有34个磷酸化位点,其中,丝氨酸磷酸化位点有23个,位于第18、72、86、90、135、140、150、200、218、222、228、231、241、244、252、268、283、289、300、301、306、366、387位氨基酸处;苏氨酸磷酸化位点有8个,位于第28、55、178、226、238、294、380、388位氨基酸处;酪氨酸磷酸化位点3个,位于第125、229、318位氨基酸处,说明MAP2K1蛋白主要是丝氨酸(67.65%)磷酸化。
2.5.8 MAP2K1蛋白糖基化分析
利用NetNGlyc1.0 Server(http://www.cbs.dtu.dk/services/NetNGlyc/)对MAP2K1突变前后蛋白糖基化位点预测,发现结果相同。当N-糖基化电势大于阈值0.5时,则该位点有可能被N-糖基化。结果显示,第21位对应天冬氨酰氨残基的酰胺氮原子存在1个N-糖基化位点,其概率为0.780 8。
2.5.9 MAP2K1蛋白结构域分析
用NCBI和SMART(http://smart.embl-heidelberg.de/)对突变前后蛋白结构域分析。由图6可知,MAP2K1有1个特殊结构S_TKc,位于68~363位氨基酸处、E值为2.04×10-79;一个卷曲螺旋结构位于19~59位氨基酸处。
2.5.10 MAP2K1蛋白二级结构预测
利用Hopfield(https://npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl?page=npsa_sopm.html)预测突变前后编码蛋白二级结构,发现结果相同。由图7可知,MAP2K1蛋白由α螺旋(44.56%)、无规则卷曲(40.76%)、延伸链(11.65%)、和β-转角(3.04%)组成。说明该蛋白主要由α螺旋和无规则卷曲组成,β-转角和延伸链散布于整个蛋白中。
2.5.11 MAP2K1蛋白三级结构预测
通过SWISS-MODEL(https://www.swiss-model.expasy.org/interactive)预测突变前后编码蛋白三级结构,发现结果相同。由图8可知,MAP2K1蛋白质三级结构与SMTL ID:6u2g.1.模板序列相似性95.92%,说明预测的蛋白三级结构与二级结构预测结果基本一致。
2.6 MAP2K1基因mRNA二级结构分析
利用软件RNAfold web server(http://nibiru.tbi.univie.ac.at/cgi-bin/RNAWebSuite/RNAfold.cgi)和RNAstructure(http://rna.urmc.rochester.edu/RNAstructure Web/Servers/Predict1/Predict1.html)分析突变前后MAP2K1基因mRNA二级结构。由图9可知,MAP2K1基因突变前后mRNA二级结序列不同,序列由A变成G,下面进一步分析突变前后mRNA二级结构的平面图、点-括号图及山地图是否存在差异。
2.6.1 mRNA最小自由能(MFE)与质心二级结构平面图分析
利用RNAfold对mRNA二级结构进行分析,由图10可知,突变前后MFE结构存在明显差异,总自由能由-1 419.11 kJ/mol上升到 -1 415.77 kJ/mol;突变前后质心二级结构发生改变,自由能由 -712.19 kJ/mol 上升到 -707.60 kJ/mol;且突变后的内环结构多于突变前,说明环区的形成是破坏结构的稳定性,突变后自由能均呈上升趋势,结构的稳定性变弱。
2.6.2 含有假结的mRNA二级结构平面图分析
利用RNAstructure分析含有假结的mRNA二级结。平面圈图将mRNA的碱基均匀地分布在圆形四周,弯成一个圆圈,在圈内用弧线或直线把配对碱基连接起来。假结结构是发卡结构环区的碱基与茎环结构外侧单链区的一些碱基形成氢键,结果形成类似于两茎连接结构,但两茎外侧形成2个环区。由图11可知,突变前后的结果不同,说明突变前后拥有的生物活性可能不同。
2.6.3 折叠结构平面图分析
利用RNAstructure分析折叠结构,预测序列的最低自由能结构和1组低自由能结构。由图12可知,突变前后折叠结构不同,且自由能从-1 447.12 kJ/mol降低到 -1 455.06 kJ/mol,说明突变前后所折叠成特有的空间结构不同,可能会对生物活性和特定的生物学功能产生影响。
2.6.4 MaxExpect结果平面图分析
利用RNAstructure分析特定结构的基团,这是一种结构预测的替代方法,在结构预测中可能具有更高的保真度。由图13可知,自由能从360.7 kJ/mol降低至 352.4 kJ/mol,突变前后形成的特定结构不同,说明突变前后形成特定结构的基团不同会影响生物活性,可能使生物功能发生改变,且突变后的结构稳定性更稳定。
2.6.5 mRNA二级结构点-括号分析
(((((....((...............))中,. (点)表示未配对碱基,()(括号)则代表表示匹配的碱基。利用RNAfold对mRNA二级结构分析,由图14、图15可知,突变前后的MFE和质心二级结构点-括号结果不同,说明变前后碱基配对情况不同,对突变前后的自由能产生影响。
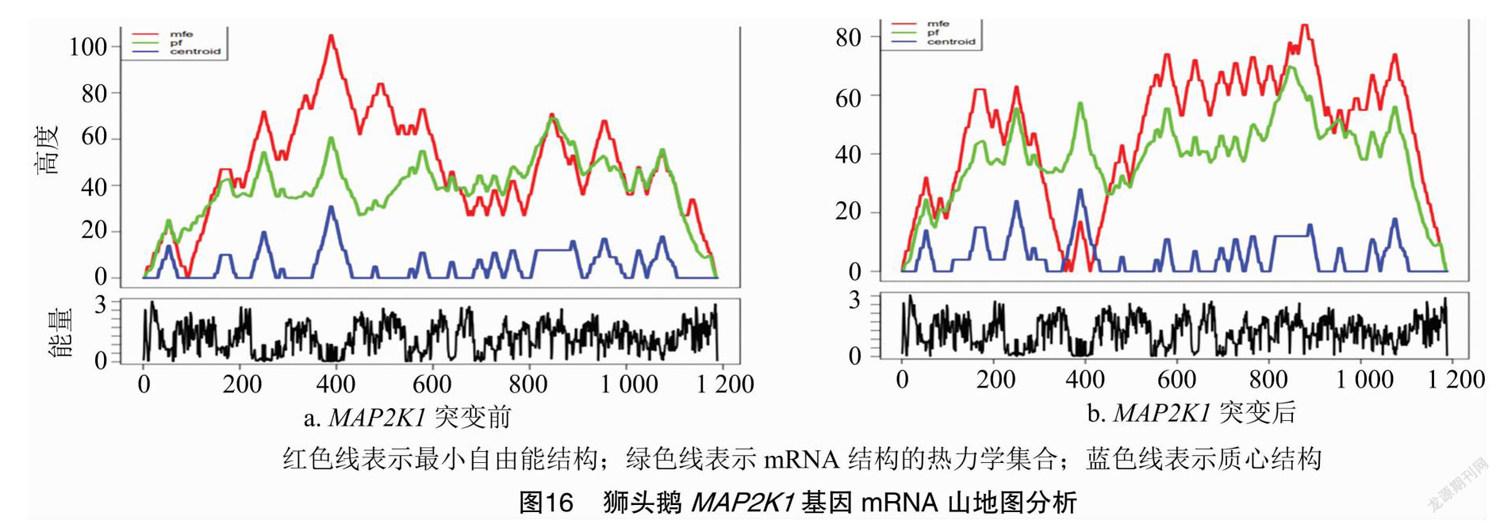
2.6.6 mRNA二级结构山地图分析
利用RNAfold对突变前后mRNA最小自由能结构、热力学集合、质心结构的山地图分析,由图16可知,突变前后山地图明显不一样,说明突变前后碱基配对不同,对自由能产生影响。
3 讨论与结论
3.1 MAP2K1相关基因的多态性分析
目前,还未发现在动物上与MAP2K1基因多态性的相关研究,而MAP2K1相关基因多态性的研究主要与猪的经济效益相关。张佳丽等研究发现,在法系大白猪MAP2K6基因有4个SNPs位点,分别为内含子8、11(Intron8、11,In8、11)和Exon12,均有3种基因型,rs325278117和rs325752048位点A为优势等位基因, rs332107877和rs345304630位点G为优势等位基因,且4个SNPs位点均与经济性状相关,影响法系大白猪的经济效益。蒲蕾等研究发现,在杜洛克猪MAP3K5基因有4个SNPs位点,30769583 A>C位点C为优势等位基因;30781169 A>G位点G为优势等位基因;30940839 A>G位点G为优势等位基因;30962276 G>A位点A为优势等位基因。30769583 A>C、30781169 A>G和 30962276 G>A在杜洛克猪群体中均处于Hardy Weinberg's law平衡状态(P>0.05);30940839 A>G在杜洛克猪群体中偏离Hardy Weinberg's law平衡状态(P<0.01)。30769583 A>C、30781169A>G和30940839 A>G低度多态,30962276 G>A为中度多态,且分别与剩余采食量、平均日采食量、饲料转化率、平均日增质量相关,影响经济效益。本研究在狮头鹅MAP2K1基因中发现1个SNP位点,位于Exon3,碱基A突变为G,并未改变氨基酸编码,G为优势等位基因,中度多态,处于Hardy Weinberg's law平衡状态(P>0.05)。MAP2K6和MAP3K5基因多态性均影响猪的经济效益,虽本研究并未与产蛋量进行相关分析,但E3-63 A>G处于中度多态,具有较大的选择潜力,是否影响狮头鹅的产蛋量还需要进行下一步验证。
3.2 MAP2K1蛋白生物信息学分析
蔡佳等研究发现,在红笛鲷MAPKK5基因cDNA序列全长2 632 bp,分子量为49.42 ku,理论等电点为6.12,编码438个氨基酸,2个保守结构域PB1和S_TKc区,22个磷酸化位点,主要在细胞质发挥生物學功能,α螺旋占36%,β折叠占11%,无规则卷曲占39%;而本研究MAP2K1基因cDNA序列全长1 188 bp,相对分子量为43 576.09,理论等电点PI为6.19,编码395个氨基酸,1个保守结构域S_TKc区,主要在细胞核发挥生物学作用,有34个磷酸化位点,蛋白二级结构由α螺旋(44.56%)、无规则卷曲(40.76%)、延伸链(11.65%)、和β-转角(3.04%)组成。说明MAPKK5与MAP2K1蛋白均属于不稳定不跨膜酸性亲水性蛋白,无信号肽存在,蛋白二级结构主要由α螺旋和无规则卷曲组成;但编码氨基酸数量、保守结构、发挥生物学作用的场所均不同。蒲蕾等研究发现,在大白猪MAP3K5蛋白同样具有保守结构域S_TKc。由此看出,MAPKK5、MAP2K1与MAP3K5蛋白均存在S_TKc结构域,S_TKc结构域具有丝氨酸/苏氨酸激酶催化功能,被上游激酶激活后,能将胞外信号传递至胞内,进而使机体对外界刺激做出应答。
3.3 MAP2K1基因mRNA二级结构生物信息学分析
mRNA二级结构主要通过调节核糖体在序列上的移动速率来调控各种生物过程,mRNA二级结构能帮助mRNA正确定位。突变前后改变mRNA二级序列,序列由A突变成G,突变前后MFE结构与质心二级结构发生改变,且自由能上升,结构稳定性变弱。突变后的内环结构多于突变前,说明各种环区的形成破坏结构的稳定性;突变前后的折叠不同,且mRNA二级结构中环结构含量与蛋白质折叠速率呈显著的正相关性,从而得知突变前后的mRNA二级结构的折叠速率不一样,折叠成不同的空间结构,不同的空间结构具有不同的生物活性,行使不同的生物学功能,突变前后的mRNA二级可能发挥不同的生物功能。假结结构在mRNA中有非常重要的地位,有研究表明在植物病毒中发现了假结的存在,发现一旦假结结构消失,RNA的生物活性也随之消失;核糖体在假结被降解,核糖体就会丧失其生物活性,不会参与到蛋白质的合成过程中,突变前后含假结结构不同,可能拥有不同的生物活性。综上,从mRNA二级结构的多方面分析,可知突变前后的mRNA可能会拥有不同生物活性及其生物功能。
本研究首次筛选了狮头鹅MAP2K1基因的SNP位点,MAP2K1基因E3-63 A>G位点属于中度多态,具有较大的选择潜力。MAP2K1蛋白属于不稳定不跨膜酸性亲水性蛋白,主要在细胞核发挥生物学作用,有34个磷酸化位点,有1个N-糖基化位点,1个保守结构域S_TKc区,蛋白二级主要由α螺旋和无规则卷曲组成。MAP2K1基因突变前后mRNA的最小自由能,质心发生二级结构,含有假结的mRNA二级结构,折叠结构,MaxExpect结果,点-括号结果,最小自由能、mRNA结构的热力学集合、质心结构所形成的山地图发生改变,且自由能上升,结构稳定性变弱。该研究可为后续提高狮头鹅产蛋量候选基因的鉴定提供理论依据。
参考文献:
[1]孙永立. 大鹅:国人餐桌一个不应被忽略的选择[J]. 中国食品工业,2019(12):12-15.
[2]阳希文,孙爱东,陈 哲,等. 狮头鹅反季节繁殖生产技术[J]. 中国家禽,2015,37(10):61-62.
[3]林树欣,潘育璇,黄 靓,等. 狮头鹅种鹅的饲养管理[J]. 广东畜牧兽医科技,2019,44(5):26-27,35.
[4]潘建秋,江丹莉,黄植霞,等. 反季节生产中马岗鹅与狮头鹅繁殖性能的观察[J]. 仲恺农业工程学院学报,2018,31(2):33-36.
[5]孙永泰. 狮头鹅的饲养技术[J]. 水禽世界,2016(2):59.
[6]岑永秀.12个MicroRNAs及其靶基因在香猪睾丸组织中的表达[D]. 贵阳:贵州大学,2017.
[7]丁 苗,涂 萍. miRNA靶向RAS-MAPK通路的表观遗传性激活与肿瘤[J]. 临床与病理杂志,2016,36(11):1868-1872..
[8]高玉花. 东北虎胎盘microRNA表达谱、转录组及microRNA-424调节MAPK通路分子机制研究[D]. 哈尔滨:东北林业大学,2016.
[9]黄 璐. miRNA424及其靶基因MEK1和FGFR1在胎儿生长受限中的作用机制研究[D]. 苏州:苏州大学,2013.
[10]李 娜. 基于GWAS的猪肉品质性状候选基因研究[D]. 北京:中国农业大学,2016.
[11]Hong F,Wu N,Ge Y,et al. Nanosized titanium dioxide resulted in the activation of TGF-β/Smads/p38MAPK pathway in renal inflammation and fibration of mice[J]. Journal of Biomedical Materials Research Part A,2016,104(6):1452-1461.
[12]Raman M,Chen W,Cobb M H. Differential regulation and properties of MAPKs[J]. Oncogene,2007,26(22):3100-3112.
[13]李 峥. 猪卵泡液外泌体对颗粒细胞增殖及孕酮合成的影响[D]. 长春:吉林大学,2020.
[14]Fan H Y,Liu Z L,Shimada M,et al. ERK1/2 in ovarian granulosa cells are essential for female fertility[J]. Biology of Reproduction,2009,81(Suppl 1):153.
[15]Moore R K,Otsuka F,Shimasaki S. Role of ERK1/2 in the differential synthesis of progesterone and estradiol by granulosa cells[J]. Biochemical and Biophysical Research Communications,2001,289(4):796-800.
[16]胡 姍. BMP15基因对绒山羊mGSCs增殖分化的影响[D]. 杨凌:西北农林科技大学,2017.
[17]王海禄. 雪兔毛色季节性变化皮肤转录组的分析[D]. 呼和浩特:内蒙古农业大学,2019.
[18]冯茵怡,严炯艺,夏 星,等. 荔枝核总黄酮对CCl诱导的大鼠肝纤维化的影响及作用机制和潜在Q-marker的预测[J]. 中国中药杂志,2020,45(23):5722-5731.
[19]Cui X D,Zhang A J,Liu J W,et al. Down-regulation of MAP2K1 by miR-539 inhibits hepatocarcinoma progression[J]. Biochemical and Biophysical Research Communications,2018,504(4):784-791.
[20]Jin Z,Jia B X,Tan L D,et al. miR-330-3p suppresses liver cancer cell migration by targeting MAP2K1[J]. Oncology Letters,2019,18(1):314-320.
[21]Zhou C,Wang P,Tu M T,et al. Long non-coding RNA PART1 promotes proliferation,migration and invasion of hepatocellular carcinoma cells via miR-149-5p/MAP2K1 axis[J]. Cancer Management and Research,2020,12:3771-3782.
[22]梁容瑞,周 蕊,李宗芳,等. MEK1表达载体的构建及其对肝癌细胞ERK通路活性的影响[J]. 西部医学,2013,25(10):1447-1450.
[23]Chuang J,Wang C K,Guo Y M,et al. MAP2K1 mutations in advanced colorectal cancer predict poor response to anti-EGFR therapy and to vertical targeting of MAPK pathway[J]. Clinical Colorectal Cancer,2021,20(1):72-78.
[24]Ishii K,Kanatsu-Shinohara M,Toyokuni S,et al. FGF2 mediates mouse spermatogonial stem cell self-renewal via upregulation of Etv5 and Bcl6b through MAP2K1 activation[J]. Development,2012,139(10):1734-1743.
[25]Nadeau V,Charron J.Essential role of the ERK/MAPK pathway in blood-placental barrier formation[J]. Development,2014,141(14):2825-2837.
[26]Bissonauth V,Roy S,Gravel M,et al. Requirement for Map2k1 (Mek1) in extra-embryonic ectoderm during placentogenesis[J]. Development,2006,133(17):3429-3440.
[27]耿红红. 不同精液品质公牛全血mRNA差异表达分析[D]. 南京:南京农业大学,2016.
[28]王慧芳,周光现,孙永峰,等. 基于全基因组重测序技术的狮头鹅Indel标记分析[J]. 畜牧兽医学报,2021,52(3):662-675.
[29]雷栩斌,贾汝敏,张锐毅,等. 狮头鹅个体产蛋智能记录系统的研制[J]. 中国草食动物科学,2021,41(1):13-17,23.
[30]杨 赫. RNA二级结构中假结的预测研究[D]. 长春:吉林大学,2013.
[31]张佳丽,聂光伟,刘 军,等. 猪MAP2K6基因多态性及其与经济性状的关联分析[J]. 中国畜牧杂志,2021,57(3):64-68.
[32]蒲 蕾,岳静伟,张金山,等. 杜洛克猪MAP3K5基因多态位点与生长、饲料利用性状的关联分析[J]. 中国畜牧兽医,2016,43(11):2982-2988.
[33]蔡 佳,黄郁葱,吴灶和,等. 红笛鲷MAPKK5基因的克隆及生物信息学分析[J]. 水产科学,2015,34(5):288-293.
[34]蒲 蕾,岳静伟,张金山,等. 猪MAP3K5基因CDS序列克隆及生物信息学分析[J]. 中国畜牧兽医,2016,43(10):2509-2517.
[35]Kuida K,Boucher D M. Functions of MAP kinases:insights from gene-targeting studies[J]. The Journal of Biochemistry,2004,135(6):653-656.
[36]李瑞芳,于志芬,黄 俏. mRNA的二级结构对蛋白质折叠速率的影响[J]. 生物物理学报,2014,30(7):497-508.
[37]李瑞芳,李 宏,郭春阳,等. mRNA環结构对蛋白质折叠速率的影响[J]. 生物化学与生物物理进展,2018,45(6):672-680.
猜你喜欢
世界科学技术-中医药现代化(2022年3期)2022-08-22
中国校外教育(下旬)(2016年11期)2016-12-27
中国教育信息化·基础教育(2016年10期)2016-12-20
今传媒(2016年11期)2016-12-19
电脑知识与技术(2016年27期)2016-12-15
科技视界(2016年23期)2016-11-04
西南农业学报(2016年6期)2016-04-16
法医学杂志(2015年4期)2016-01-06
中央民族大学学报(自然科学版)(2015年1期)2015-06-11
河南医学研究(2014年7期)2014-02-27
