
针叶林补阔对土壤有机碳、氮含量的影响*
2022-06-16 14:49:44彭素琴刘郁林刘苑秋樊乙萱周玉姗邹文文
赣南师范大学学报 2022年3期
彭素琴,刘郁林,刘苑秋,樊乙萱,周玉姗,杨 琪,邹文文
(1.赣南师范大学 地理与环境工程学院,江西 赣州 341000;2.江西农业大学 林学院,南昌 330045,3.江西环境工程职业学院,江西 赣州 341000)
亚热带红壤区是我国土壤侵蚀最严重的区域之一.为控制水土流失、提高土壤肥力,马尾松(Pinusmassoniana)等针叶树种作为主要造林树种在该地区植被恢复过程中得到大规模的推广应用.近30多年,马尾松面积迅速扩大,仅江西省马尾松面积就达到了239.4万hm2.针叶纯林树种结构单一、林分结构不合理[1-2],导致地力衰退[3-5]、生态效益差[6]、生物多样性丧失、林分生产力下降和土壤养分元素亏缺[7]等问题,极大地影响了森林生态功能的发挥.乡土阔叶树种相比针叶树种根系系统更复杂、凋落物分解速率更快、土壤肥力和生态系统服务功能更高[8],乡土阔叶树种可以在生物多样性保护、生态系统稳定性改善和生态功能等方面发挥重要作用[9],营造针阔混交林正逐渐成为替代世界上大面积分布的人工针叶纯林最有前景的森林经营模式之一[10].
针叶林补阔成为优化林分结构、提高森林质量的有效措施,研究表明针叶林中混交阔叶树后形成针阔混交林是改善土壤肥力、增强森林土壤养分循环和生态系统稳定性的重要措施[11],混交林林地生产力明显高于林分结构单一的人工纯林[12],因此混交林通常比纯林有更高水平的生态系统功能[13].人工针阔混交林的土壤有机碳和氮储量显著高于针叶和阔叶纯林[14-15],石漠化地区针阔混交林相比针叶纯林土壤SOC、TN显著提升[16],但不同阔叶树种其套种效果不同[17].
为此,本文以亚热带退化红壤区江西泰和为研究代表区域,利用乡土树种木荷(Schimasuperb)进行补阔调整的马尾松木荷混交林分土壤为研究对象,马尾松纯林和木荷纯林做对照,探讨马尾松纯林补阔后不同林分土壤全碳、氮含量及其季节分布差异,以揭示针叶林补阔后土壤碳、氮的变化规律,进一步阐明退化红壤植被恢复机理,为人工林科学经营提供科学依据.
1 研究区概况与研究方法
1.1 研究区概况
试验地位于江西省泰和县石山乡(115°04′E,26°44′N).该试验区属亚热带季风湿润性气候,年平均气温18.6 ℃,绝对最低气温-6 ℃,绝对最高气温41.5 ℃.20世纪90年代以前,试验区内植被覆盖度仅0.3.最高海拔为131.3 m,最低海拔为74.7 m,有机质含量为6.5 g/kg,pH为4.9,土壤成土母质是第四纪红粘土,属于典型的红壤低丘岗地,为中亚热带区典型退化红壤区.1991年选择马尾松(Pinusmassoniana)、湿地松(Pinuselliottii)、枫香(Liquidambarfomosana)、木荷(Schimasuperb)等12个针阔叶树种进行植被恢复荒山绿化,开展不同树种、不同混交组合等植被恢复与重建试验,建立了面积为133 hm2试验林.
1.2 样地设置
2009 年利用乡土树种木荷进行补阔恢复的马尾松木荷混交林分为研究对象,马尾松纯林、木荷纯林作为对照.在每种林分类型中选择具有相近坡向、坡位和坡度的代表性林分各4个小班,每个小班设置1个20 m×20 m 样地,即每种林分4个重复,共计12个样地(见表1).
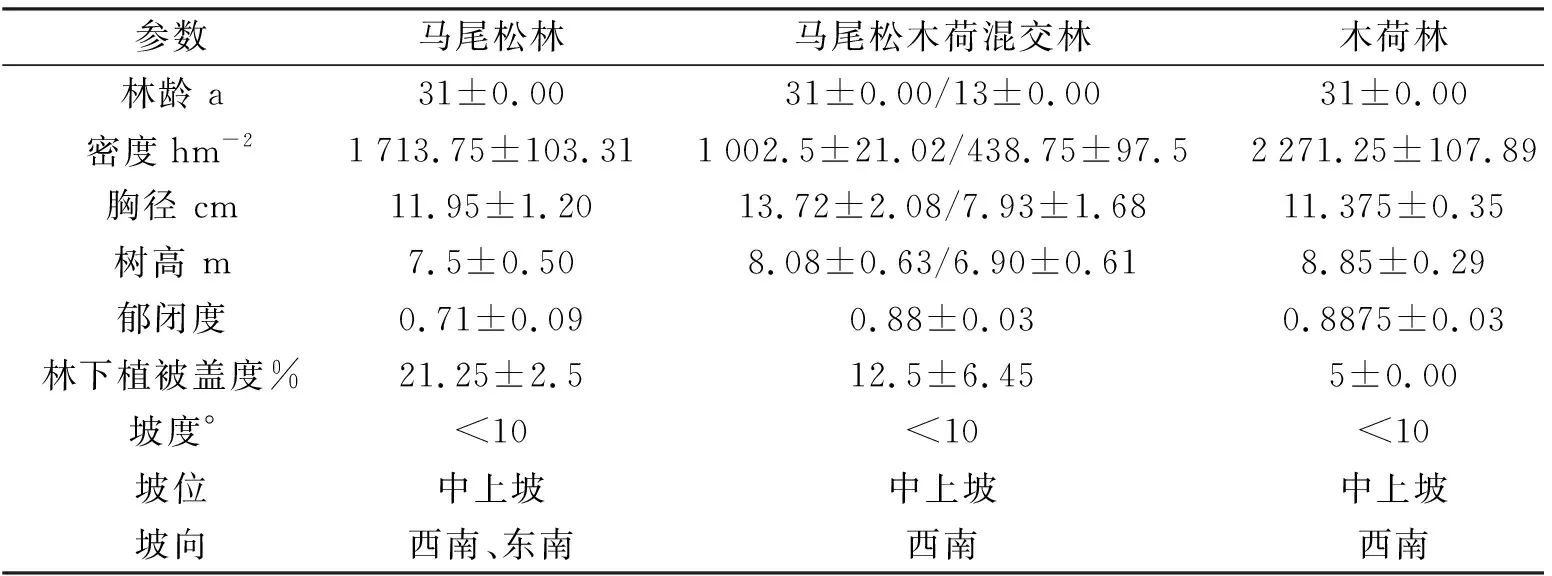
表1 样地基本情况
1.3 样品的采集与测定
按照对角线五点取样法在各样地中采集土壤样本,先去除枯枝落叶等地表凋落物,然后采集0 cm~10 cm土壤样品,保持每个采样点的取土深度和采量均匀一致,同时用土壤三参数速测仪测定10 cm处的土壤湿度和温度.最后将每个小区中采集的土样过 2 mm钢筛后混匀为一个混合样,自然风干,测定土壤理化性质.
测定方法参考《土壤农化分析》[18],方法如下:
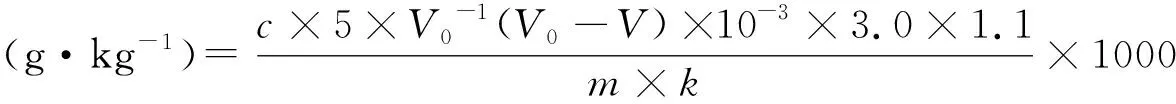
1.4 数据处理
用Excel2010对数据进行初步的整理和计算,用SPSS 26.0统计软件对所有数据进行单因素、多因素方差分析及相关性分析,比较不同林分类型、土层、季节之间的差异,显著性水平设定为α=0.05,多重比较采用 LSD 法(P=0.05),论文所有数据的图形处理采用Orgin2018软件制作.
2 结果与分析
2.1 不同林分特征及凋落物状况
由表2可知,3种林分年凋落物量马尾松木荷混交林>木荷纯林>马尾松林,但林分间差异不显著.3种林分间马尾松林凋落物碳含量显著高于马尾松木荷混交林和木荷纯林,林分间差异显著,马尾松木荷混交林凋落物氮含量显著高于纯林,林分间差异显著,马尾松林凋落物碳氮比高于马尾松木荷混交林和木荷纯林.
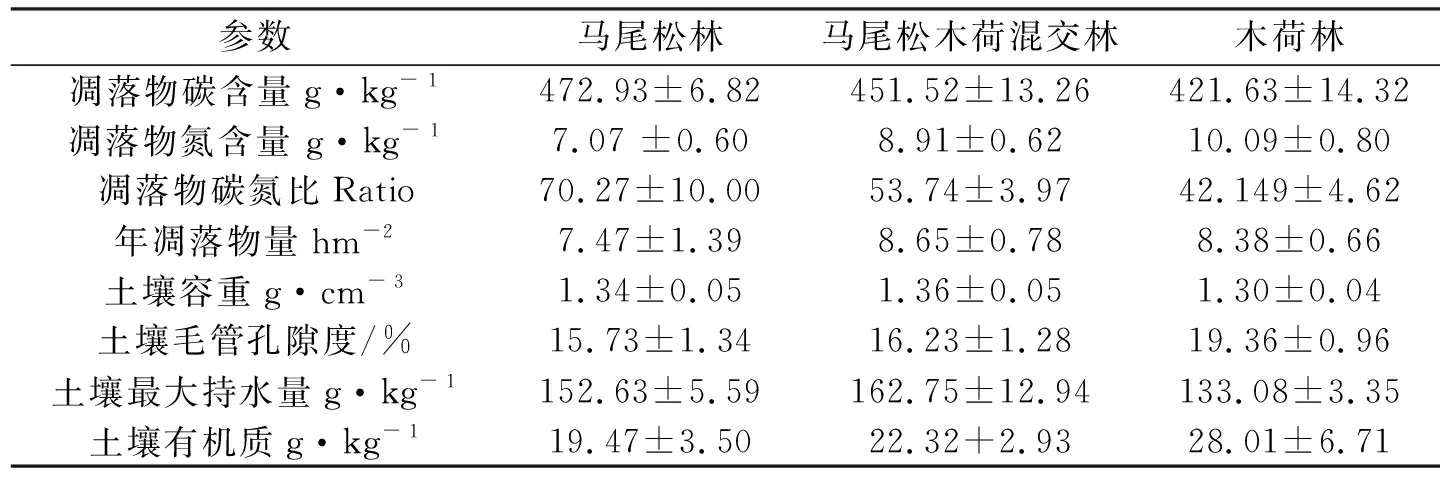
表2 不同林分土壤、凋落物基本理化性质
2.2 不同林分土壤pH、温度、湿度和含水率的季节性变化特征
由表3可知,马尾松纯林、马尾松木荷混交林和木荷纯林土壤pH均呈酸性,3种林分土壤pH值均在春季最大,秋季最低,秋季与春夏冬差异显著,同一林分内春夏冬差异不显著.3种林分土壤含水率和土壤湿度在季节间变化存在明显差异,3种林分均在夏季最高,秋季最低,其中马尾松纯林春季和夏季含水率几乎相同.3种林分土壤温度存在明显的季节性变化差异,均在秋季温度最高,冬季最低.同一季节,马尾松木荷混交林土壤温度大多高于马尾松纯林和木荷纯林.
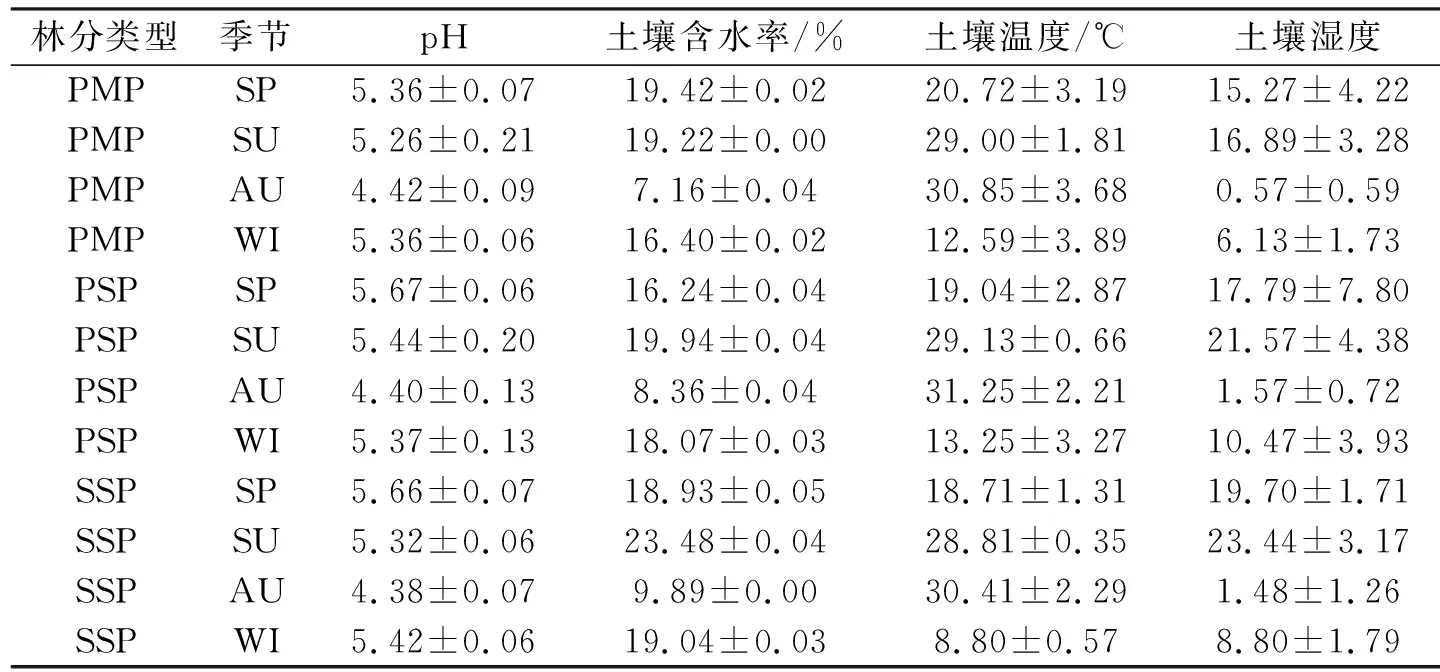
表3 不同林分0 cm~10 cm土壤pH、温度、湿度和含水率
2.3 不同林分土壤有机碳、全氮、C/N的季节性动态
从表4可以看出,林分、季节对土壤有机碳、氮及土壤碳氮比影响极显著,但季节和林分的交互作用对土壤有机碳、氮及土壤碳氮比均无显著影响.
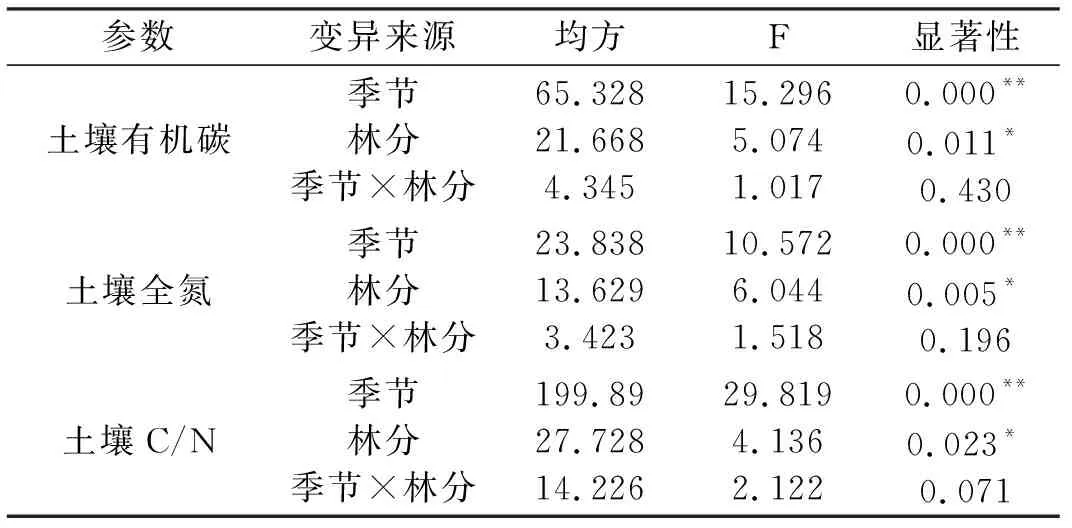
表4 土壤碳、氮的多因素方差分析
2.3.1 不同林分土壤有机碳含量的季节性动态
从图1(注:不同小写字母表示同一季节各林分间存在显著性差异;不同大写字母表示同一林分不同季节间存在显著性差异(P<0.05),下同)可以看出,与马尾松林和马尾松木荷混交林相比,木荷林具有较高的土壤有机碳含量,马尾松木荷混交林次之,马尾松林相对较低,表明马尾松纯林补植木荷阔叶树后显著提升了土壤有机碳含量.春季和秋季3种林分间土壤有机碳含量无显著差异(P>0.05),夏季马尾松木荷混交林、木荷林显著高于马尾松林(P<0.05),但马尾松木荷混交林与木荷林之间差异不显著(P>0.05).除春季外,马尾松木荷混交林各季节土壤有机碳含量均高于马尾松纯林,夏、秋、冬季比马尾松林显著提高45%、4%、10%.在同一林分中,土壤有机碳含量在4个季节中的变化趋势相同,表现出先升后降趋势,春季含量均最低.马尾松林春、夏、秋季之间呈上升趋势,冬季较秋季略下降,秋季最高,秋季相比春、夏、冬季分别高71%、25%、8%,季节之间无显著差异(P>0.05).马尾松木荷混交林和木荷林土壤有机碳含量季节变化一致,均在夏季达到最高后下降,其中马尾松木荷混交林春、秋、冬相比夏季分别减少了50.2%、9.7%、11.6%,木荷林春、秋、冬相比夏季分别减少了41.0%、15.8%、28.1% ,马尾松木荷混交林和木荷林夏、秋、冬3季显著高于春季(P<0.05),但夏、秋、冬3季之间不显著(P>0.05).
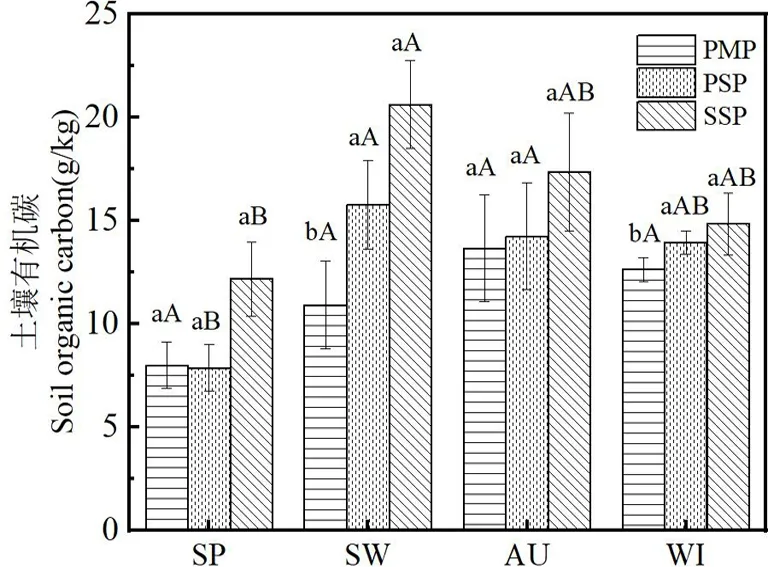
图1 不同林分土壤有机碳含量

图2 不同林分土壤全氮含量
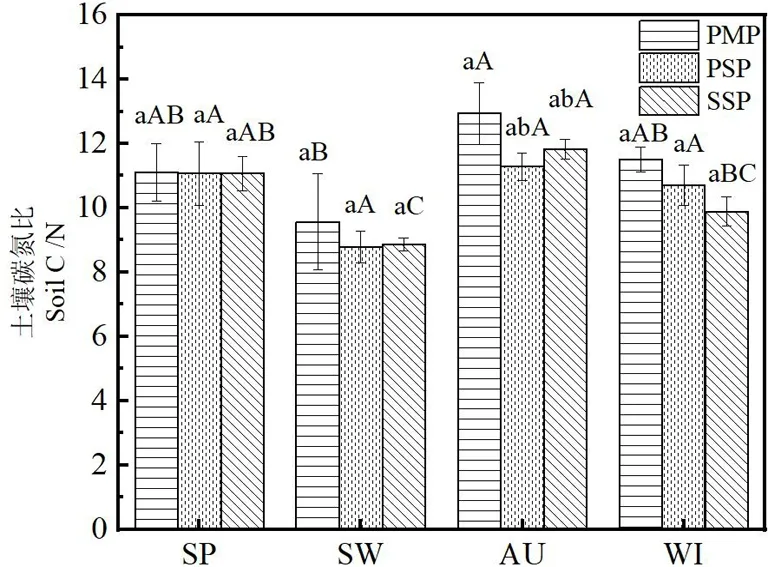
图3 不同林分土壤C/N
2.3.2 不同林分土壤全氮的季节性动态
3种林分土壤全氮含量如图2所示,土壤全氮含量均表现为木荷林>马尾松木荷混交林>马尾松林,表明马尾松林补植木荷阔叶树后提升了土壤N储量,其中春、秋、冬季3种林分全氮含量差异不显著,夏季木荷林显著高于马尾松林和马尾松木荷混交林(P<0.05),且马尾松林和马尾松木荷混交林分间无显著差异(P>0.05),夏季木荷林和马尾松木荷混交林分别比马尾松林分别高102%、53%.同一林分中,3种林分全氮含量表现出显著季节差异(P<0.05),夏季显著高于春、秋、冬季,且马尾松林冬春季显著差异,马尾松木荷混交林和木荷林春、秋、冬季间无显著差异.
2.3.3 不同林分土壤C/N的季节性动态
从同一林分不同季节土壤来看(图3),碳氮比变化规律相对较为复杂,介于 8.78~12.94之间,马尾松木荷混交林和木荷林各季节C/N比均低于马尾松林,表明马尾松林补植木荷阔叶树后降低了土壤C/N.3种林分碳氮比均在秋季最高,夏季最低.马尾松林夏季显著低于秋季,春冬季间无显著差异,马尾松木荷混交林季节间无显著差异,木荷林秋季显著高于夏、冬季,且与春季无显著差异.从同一季节不同林分来看,各季节马尾林C/N比均高于马尾松木荷混交林和木荷纯林,春、夏、冬季3种林分间C/N差异不显著,春季3种林分碳氮比几乎无变化,秋季马尾松显著高于马尾松木荷混交林和木荷纯林(P<0.05),分别高出1.66、1.12,马尾松木荷混交林和木荷纯林间无显著差异,可以看出马尾松纯林补植木荷阔叶树后土壤C/N较马尾松纯林明显降低,林地土壤的C、N含量明显提高,反映马尾松木荷针阔混交林具有良好的土壤养分循环累积机制.
2.4 土壤碳、氮及C/N与环境因子的关系
表5为土壤碳、氮因子和环境因子的相关性分析.土壤有机碳与土壤温度、土壤全氮、凋落物氮含量显著或极显著正相关,与土壤pH极显著负相关(P<0.01);土壤全氮与土壤含水率、凋落物氮含量显著正相关(P<0.05);土壤碳氮比与湿度、pH、土壤含水率、凋落物碳含量显著或极显著负相关.

表5 土壤碳、氮与环境因子的Pearson相关系数
3 讨论
3.1 针叶林补阔对土壤有机碳的影响
森林土壤有机碳主要来源于植被地上的凋落物和其地下根系分泌物以及死亡的根系.因此植被类型不同会影响森林土壤有机碳的数量、质量及周转.在本研究中,马尾松纯林补种阔叶树木荷后变成马尾松木荷针阔混交林,大大提升了土壤有机碳含量,这主要是马尾松纯林补植阔叶树木荷后,由原来的马尾松纯林变为异龄复层的针阔混交林[19],林分结构发生了很大的改变,从而影响了林地结构和碳储量.另外,不同森林类型土壤中有机碳组分稳定性及凋落物输入差异也是导致碳含量差异的原因[20].与针叶树相比,阔叶树种具有较高的年凋落物量和凋落物的浸出物,因此有机碳的输入量较高.本研究中马尾松木荷混交林、木荷林相比马尾松林,具有较高的年凋落物量,不同森林类型外源碳库输入的差异,是导致不同森林类型土壤有机碳含量差异显著的原因之一.林木自土壤中吸收的矿物质养分相当一部分是以凋落物的形式归还土壤,由于不同树种生物学特性不同,使得其凋落物的质和量及分解速率均有较大的差异,从而影响土壤的养分状况[21].马尾松纯林经过套种阔叶树改造后,其分解速率比马尾松纯林有较大的提高,回归土壤的养分量较多,而且土壤有机碳含量与凋落物、土壤氮含量显著正相关(表5),从而提高了林地的有机碳含量[15].
3.2 针叶林补阔对土壤全氮的影响
森林植被氮储量受众多因素影响,如生物量和树种生物学特性等[14].本研究中,马尾松林土壤全氮含量全年均低于马尾松木荷混交林和木荷林,马尾松针叶纯林补阔后会提高土壤氮储量,说明马尾松木荷混交林和木荷林土壤肥力高,有机质较易转化为氮素.不同林型之间氮储量的差异主要与不同植被凋落物生物量、性质及输入差异有关,本研究中马尾松林凋落物叶片分解率相对较低,因此马尾松林的凋落物较难分解,土壤养分较马尾松木荷混交林低,土壤中微生物活性相对较弱,将土壤有机质分解为氮素的能力也就小于马尾松木荷混交林及木荷纯林,此外,马尾松木荷混交林和木荷林年凋落物生物量高于马尾松林,同等时间内积累的生物量更多,氮储量也越高.本研究中3种林分土壤碳和氮的变化趋势相同,且土壤全碳与全氮含量极显著正相关,说明土壤碳素积累与氮储存关系密切.
3.3 针叶林补阔对土壤碳氮比的影响
土壤C/N是土壤质量的敏感指标,在一定程度上能反映有机物质分解程度,是土壤氮素矿化能力的标志,碳氮比低则促进微生物的分解,氮的矿化速率就高.本研究中,与马尾松林相比,木荷、马尾松木荷混交林C/N相对较低,均小于15,木荷林与马尾松木荷混交林之间无显著差异,这说明马尾松木荷混交林与木荷林土壤氮矿化较强[22],马尾松针叶纯林补阔后会降低土壤C/N,从而提高氮矿化速率和氮有效性.
此外,马尾松补阔后形成异龄复层的针阔混交林,相比2种纯林,优化了树种组成,改善了林下凋落物的质量,凋落物生物量显著提升,同时促进了树种多样性,改善了纯林林分结构单一现象,森林空间结构得到进一步优化.
4 结论
针叶林补阔显著改变土壤有机碳、氮浓度.在3种林分中,木荷林的土壤有机碳和全氮最高,其次为马尾松木荷混交林,马尾松林最低,而马尾松木荷混交林和木荷林C/N低于马尾松林.说明马尾松纯林补种阔叶树木荷后,大大提升了土壤有机碳、氮含量,降低了土壤C/N,从而提高氮矿化速率和氮有效性,反映马尾松木荷针阔混交林具有良好的土壤养分循环累积机制.
猜你喜欢
中国林副特产(2022年4期)2022-08-08 05:50:30
散文诗世界(2022年5期)2022-05-23 02:02:55
防护林科技(2022年2期)2022-04-25 23:47:32
防护林科技(2019年12期)2019-04-14 04:01:18
生态学报(2018年21期)2018-12-19 10:47:28
水土保持通报(2018年3期)2018-07-26 00:41:46
湖南林业科技(2017年1期)2017-02-06 05:28:55
河北林业科技(2016年5期)2016-11-08 03:13:01
现代农村科技(2016年13期)2016-08-09 03:33:38
花卉(2016年14期)2016-07-31 18:46:24
