
采伐干扰对吉林蛟河针阔混交林生产力稳定性的影响*
2022-06-15 08:46张春雨赵秀海
林业科学 2022年3期
刘 晨 张春雨 赵秀海
(北京林业大学森林资源和环境管理国家林业和草原局重点实验室 北京 100083)
近年来生物多样性正以人类历史上前所未有的速度下降,导致重要生态系统功能不可避免地降低(Isbelletal., 2011; Cardinaleetal., 2012; Liangetal., 2016; Huangetal., 2018)。生物多样性与生态系统功能及其稳定性之间的关系已成为生态学的研究热点(Davidetal., 2000; Ruijvenetal., 2005; Tilmanetal., 2014; Fons, 2019)。多样性对生态系统过程的稳定效应最好的例子来自草地试验,如许多研究表明多样性较高的草本群落的初级生产力的年际波动与物种贫乏的群落相比一直较低(Hautieretal.2014),这些结果经常被引用为多样性能够稳定包括森林在内的植物群落生产力的证据(Loreauetal., 2013)。但是,森林与草地生态系统在很多关键过程上存在差异,如草本群落物种组成年际变化较大,而森林群落变化相对缓慢,尤其是多样性-稳定性背后的关联机制会因生态系统而异(Loreauetal., 2013),所以,探究森林生态系统中多样性与稳定性的关系显得尤为重要。
群落生产力的稳定性衡量的是生产力在多年之间围绕它的长周期均值的波动情况(Tilman,1999)。因此,生产力稳定性可表示为平均生产力(μ)与其时间变化(σ,表示生产力的标准偏差)的比率,即生产力变异系数的倒数,任何影响μ或σ的过程都会改变生产力稳定性(Lehmanetal., 2000; Isbelletal., 2010)。采伐是影响森林生产力稳定性的重要因素,可通过增加光的可利用性来补充树木和提高树木生长速率(Clarosetal., 2008)。合理的抚育方式和强度有利于林分大小结构的调整,同时导致物种多样性的变化(Dagleyetal., 2018; 常伟等, 2015; 罗菊春等, 2006)。物种多样性基于生态位互补机制来维持植物群落的生产力稳定性(Loreauetal., 2013),而林分结构对生态系统生产力稳定性的影响至今仍知之甚少(Mazzochinietal., 2019)。非生物因素决定了植物生长和生存的资源可用性(Sandeetal., 2017),它们是群落生产力稳定性的强大驱动力,如土壤条件是整个亚马逊热带森林生长的决定因素(Quesadaetal., 2012; Paolietal., 2005)。因此,明确采伐干扰背景下的林分密度、土壤养分、林分大小分化和物种多样性与生产力稳定性的关系对提高森林质量促进森林可持续经营具有重要意义(李建等, 2017; 郝珉辉等, 2018)。
本研究以吉林蛟河长期监测采伐样地为对象,分析采伐干扰对生产力稳定性的影响,阐明物种多样性和林木大小分化的直接和间接效应。重点解决如下问题: 1)生物多样性和林木大小分化属性对森林生产力稳定性的相对贡献大小如何?2)采伐干扰如何影响生产力稳定性的变化?旨在通过揭示针阔混交林生产力稳定性的决定机制,为东北森林的可持续经营提供理论支持。
1 研究区概况
研究区位于吉林省蛟河林业试验区,属长白山系张广才岭山脉。该地属温带大陆季风性气候,年均气温3.8 ℃,最热月7月的平均气温21.7 ℃,最冷月1月的平均气温-18.6 ℃,年均降水量700~800 mm。研究区土壤为山地暗棕色森林土,土层厚20~100 cm。研究区植被为长白山植物区系,森林类型为天然次生林,处于由中龄林向近熟林的过渡阶段,主要乔木树种有红松(Pinuskoraiensis)、白牛槭(Acermandshuricum)、胡桃楸(Juglansmandshurica)、春榆(Ulmusdavidianavar.japonica)和水曲柳(Fraxinusmandshurica)等。主要灌木种有毛榛子(Corylusmandshurica)、簇毛槭(Acerbarbinerve)和东北山梅花(Philadelphusschrenkii)等。主要草本有猴腿蹄盖蕨(Athyriummultidendatum)、毛缘苔草(Carexpilos)和小叶芹(Aegopodumalpestre)等。
2 研究方法
2.1 样地设置与调查
2011年7月在吉林蛟河林业实验区建立4块100 m×100 m针阔混交林采伐样地。4块样地呈田字形排列,彼此间距100 m,具有相似的地形状况、均一的林分条件、相似的物种组成。利用全站仪将4块样地划成100个20 m×20 m的连续样方,调查样地内所有胸径≥1 cm树木的物种名称、胸径、树高、冠幅和位置坐标等,并挂牌标记用于长期观测,样地概况见表1。采伐后观察森林状态,后于2013、2015和2018年每年7月份进行了复测调查。

表1 样地概况Tab.1 Survey of sample plots
2011 年12 月对4 块样地进行了不同强度的抚育采伐。采伐强度通过胸高断面积来控制,确定采伐木时考虑保留木的林分结构和物种多样性,根据保留木的相邻距离、树种及林木大小确定采伐木,去除距离太近的相邻木1株或多株,尽量去除先锋树种或个体数量较多的树种,优先去除生长状况不良林木和霸王树等。间密留匀、留优去劣,以达到林木分布均匀、林分结构更合理、维持物种多样性的目的(周建云等, 2012) 。记录采伐木的号牌,结合采伐前观测数据,经计算后得到100个连续样方的采伐强度,实际采伐强度为2.7%~77.6%,平均值为39.2%。
2.2 生产力稳定性计算
每棵树木的地上生物量是基于树木胸径使用物种特定的生长异速方程计算的,这些方程基本包含了研究区内的树种(Heetal., 2018)。对部分缺失公式的树种,使用通用的异速生长方程来计算(Wang, 2006),确保计算出样方内所有树木的地上生物量。
以100个20 m×20 m样方为基本单位,计算每个样方内所有胸径≥5 cm林木的单株地上生物量,将单株生物量求和得到2011、2013、2015和2018年的现存生物量。生物量生产力为每个调查周期内样方生物量的增量,用每个样方3个调查周期的生产力均值除以其标准差来计算生产力稳定性,计算公式(Wangetal.,2014)为:
式中: SPi为第i个样方的生产力稳定性;σi为第i个样方的生物量生产力标准差;μi为第i个样方生物量生产力的平均值。
2.3 物种多样性与林木大小分化指标
物种多样性用物种丰富度、Simpson指数和Shannon-Wiener指数来量化(Shannon,1949;Simpson,1949)。林木大小分化用胸径Shannon-Wiener指数、胸径Simpson指数、胸径基尼指数和胸径变异系数来量化(娄明华等, 2019)。计算林木大小分化属性时,以20 m×20 m的样方为单位,以胸径5 cm为间隔划分径级,分别计算每个样方中的径级数和每个径级的株数。用径级数代替物种数代入Shannon-Wiener指数和Simpson指数计算公式,计算出胸径Shannon-Wiener指数及胸径Simpson指数。各指标计算公式见表2。

表2 物种多样性和林木大小分化指标的计算公式①Tab.2 Formulas of species diversity and size differentiation in the studied forest
2.4 土壤指标测定
在每个20 m×20 m样方内进行土壤取样。以每个样方西北角为中心,在半径为2 m的圆形区域内避开植物根系及岩石,收集0~20 cm的表层土500 g,并用铝盒收集约20 g新鲜土样。铝盒内土壤立即称鲜质量后放入105 ℃烘箱烘干,后冷却称干质量求出含水量,剩余土壤带回实验室阴干和研磨以测定土壤化学指标,酸碱度采用电极电位法测定(Allisonetal., 1968),全碳含量用硫酸消煮-重铬酸钾外加热法测定(Walkleyetal., 1934),全氮含量用凯氏定氮法测定(吴晓荣等, 2009),全钙含量用气量法测定(鲍士旦, 2000),全磷含量用硫酸-高氯酸消-铜磷抗比色法测定。因土壤养分含量之间有强相关性,对土壤养分含量进行主成分分析,第一主成分的贡献率达到73%,故选用土壤养分含量第一主成分作为土壤养分的综合指标,各主成分载荷系数及其贡献率见表3。
2.5 数据处理
采用多元线性回归分析对物种多样性和林木大小分化指标进行筛选。在筛选之前,用Z-score方法对数据标准化处理(Foxetal., 1992)。利用方差膨胀因子(VIF)对转换后的数据进行多重共线检验,VIF<10 时认为变量之间没有多重共线性(Fox, 2008)。对保留的变量进行方差分解计算,分析不同类型变量对生产力稳定性的解释能力。方差分解运算用R软件vegan包的“varpart”函数进行。基于多重共线的检验结果,在参照以往模型形式的基础上(Cardinaleetal., 2009; Graceetal., 2012),将结构方程模型的基本形式定义为:
SP=a1× LI+a2× NT+a3×BAD+
a4× SD +a5× SS + b。
式中: SP为生产力稳定性; NT为土壤养分;LI为采伐强度; BAD为林分密度; SD为物种多样性; SS 为林木大小分化; b 为常数项;a1~a5为对应解释变量的系数。

表3 各主成分载荷系数及其贡献率Tab.3 Loadings and proportion of explained variance of each principal component
为确定最佳模型形式,用拟合优度指数(GFI)和标准化残差均方根(SRMR)评价结构方程模型的拟合效果。GFI 接近1时代表拟合非常好,大于0.95小于1时拟合较好,大于0.90可以接受; 当SRMR 小于0.05时表示模型可接受。通过比较GFI和SRMR值确定最终的变量和模型形式。结构方程模型采用R软件的lavaan包进行运算。
3 结果与分析
3.1 模型变量的筛选和方差分解分析
根据多重共线性检验和逐步回归分析结果,选定胸径Shannon-Wiener指数表征林木大小分化,物种Simpson指数表征物种多样性。以生产力稳定性为响应变量,进行多元线性回归,计算每个解释性变量的系数和标准误差,发现林分密度、胸径Shannon-Wiener指数、物种Simpson指数显著影响生产力稳定性(P<0.05),土壤的全碳、磷、钙、氮含量和酸碱度及含水量对生产力稳定性无显著影响(图1)。采伐强度对生产力稳定性无显著影响,因而检验采伐强度是否通过物种多样性或林木大小分化间接影响生产力稳定性。
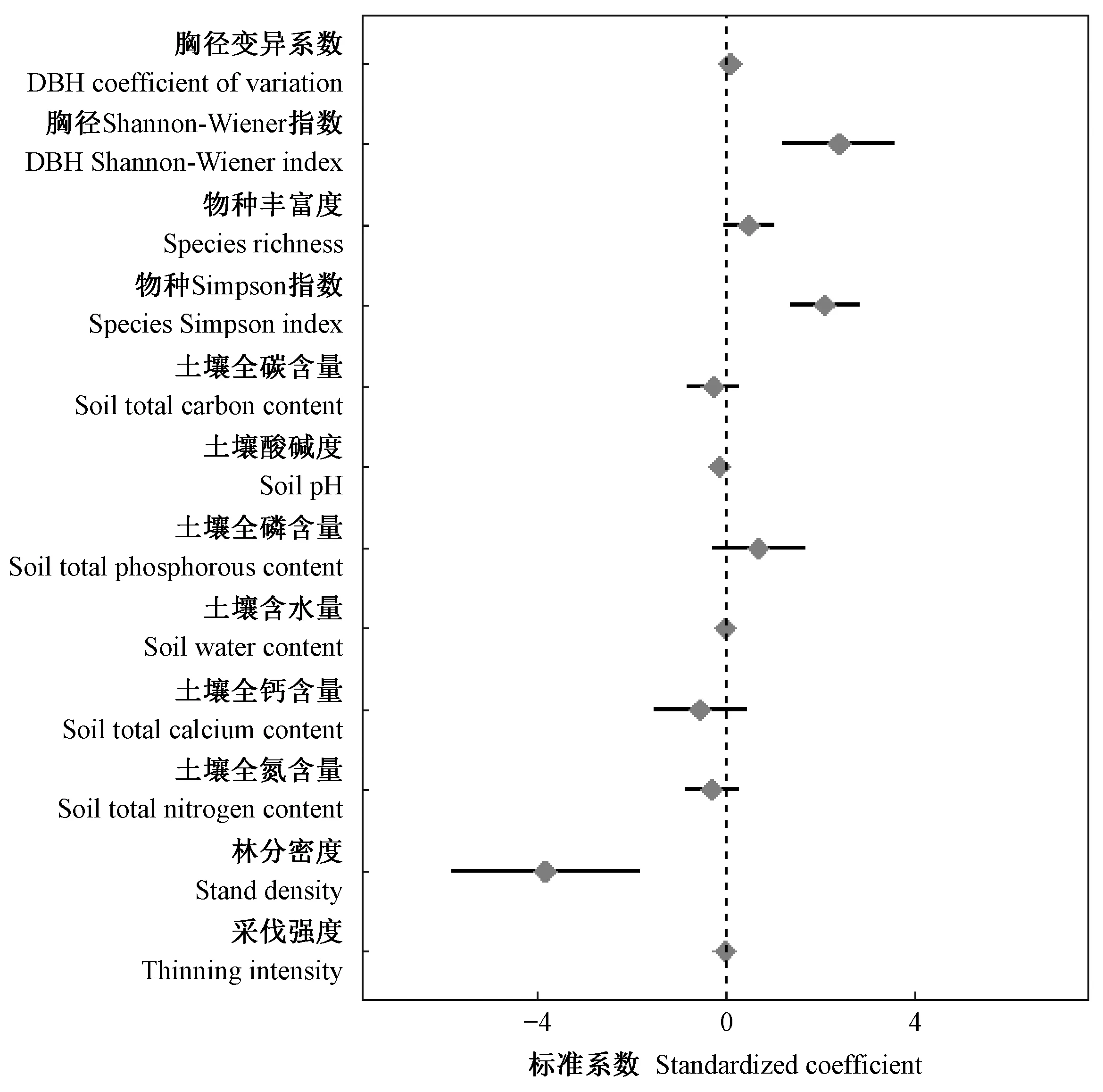
图1 多元线性模型变量的标准系数Fig. 1 Standard coefficient of variables in multivariate linear model
利用方差分解分析量化林分密度、物种多样性和林木大小分化单独及交互作用对生产力稳定性的贡献(图2)。林分密度和林木大小分化的单独解释量均不显著。物种多样性单独解释量在P<0.05水平上显著,解释比例高达19.5%。物种多样性与林分密度的交互作用解释比例较高,达到了12.8%。

图2 各变量影响生产力稳定性的方差分解图Fig. 2 Variance decomposition diagram of different variables affecting productivity stabilityA: 林分密度Stand density; B: 物种多样性Species diversity; C: 林木大小分化Tree size differentiation; A×B: 林分密度和物种多样性的交互作用Interaction between stand density and species diversity; A×B×C: 林分密度、物种多样性和林木大小分化的交互作用Stand density, species diversity,and tree size differentiation interaction.
3.2 采伐强度对生产力稳定性的直接、间接和总效应
利用结构方程模型检验解释性变量对生产力稳定性的直接和间接效应,最优模型与数据之间有较高匹配度(GFI = 1.000; SRMR = 0.005)。采伐强度对林分密度、土壤养分和林木大小分化有显著的负向影响(P<0.05),路径系数分别为-0.629、-0.316和-0.255;采伐强度对物种多样性和生产力稳定性没有显著影响,通过林分密度、土壤养分和林木大小分化间接影响物种多样性;林分密度对物种多样性和林木大小分化有显著的正向影响(P<0.05),路径系数分别为1.877和0.721;土壤养分对物种多样性和林木大小分化有显著的正向影响(P<0.05),路径系数分别为0.060和0.086(图3)。林分密度和林木大小分化对生产力稳定性有显著的负向总效应(P<0.05),分别为-2.365和-0.556; 物种多样性对生产力稳定性有显著的正向总效应(P<0.05),路径系数为1.652; 土壤养分对生产力稳定性的总效应不显著;采伐强度对生产力稳定性的直接效应不显著,主要通过林分密度和林木大小分化影响生产力稳定性,采伐强度对生产力稳定性有显著的负向总效应(P<0.05),路径系数为-0.290(表4)。
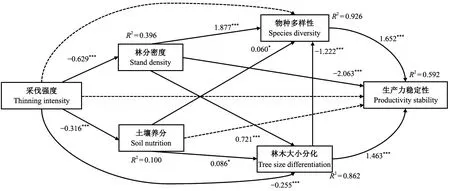
图3 以采伐强度、林分密度、土壤养分、物种多样性和林木大小分化为自变量、以生产力稳定性 为响应变量的结构方程模型Fig. 3 Structural equation models with thinning intensity, stand density, soil nutrition, species diversity, and tree size differentiation as the independent variables and productivity stability as the respons variables物种多样性为物种Simpson指数,林木大小分化为胸径Shannon-Wiener指数,林分密度为胸高断面积密度,土壤养分为土壤变量的第一主成分。实线表示作用路径显著,虚线表示作用路径不显著。 *表示P<0.05,**表示P<0.01,***表示P<0.001。Species diversity is the species Simpson index, size differentiation is the diameter at breast height Shannon-Wiener index, stand density is the breast height section density, and soil nutrients are the first principal component of soil variables. Solid line indicates significant path, while dashed line indicates insignificant path. * indicates significant at P< 0.05 level; ** indicates significant at P < 0.01 level; *** indicates significant at P < 0.001 level.

表4 结构方程模型中各变量的直接、间接和总效应Tab.4 The direct, indirect, and total effects of variables in structural equation models
4 讨论
4.1 林木大小分化和物种多样性对生产力稳定性的影响
森林生态系统具有复杂的群落结构,林木大小分化是群落生物量或生产力的有力预测指标(Wuetal., 2015; Fotisetal., 2017; Zhangetal., 2015; Ouyangetal., 2019),但林木大小分化如何影响森林生产力稳定性很少被关注。本研究证实了林木大小分化对生产力稳定性有显著的正向直接效应(P<0.05),林木分化程度越高,林分生产力稳定性也越高。林木大小分化反映了树木大小分布的异质性,结构复杂的林分由于个体大小差异明显,降低了彼此的资源竞争,增加了资源利用率,有利于树木个体稳定生长(谭凌照等, 2017)。树木大小分布的异质性越大,林分冠层结构越复杂,树木的光捕获能力也越强。林内光利用效率的增加(van der Plas, 2019; Yachietal., 2007; Davidetal., 2009),促进了树木平均生物量生产进而提高森林生产力。
本研究显示物种多样性高促进了生产力稳定性,与以往研究结论一致(Tilman,1999; Hautieretal., 2014; Hectoretal., 2010)。提高物种多样性会增大物种动态不同步性,增强物种之间的时间互补性,减少随时间变化的年际方差,从而提高生产力稳定性。此外,物种多样性增大也可通过加强竞争性作用而破环单个种群的动态,同时减少整个群落生物量生产的可变性,从而增加林分水平生物量生产力的时间稳定性(Tilmanetal., 2006; Proulxetal., 2010)。与Vil等(2013)的研究结论一致。林木大小分化既可直接作用于生产力稳定性,也可通过物种多样性间接作用于生产力稳定性。以往的生物多样性研究更加强调物种多样性的影响(Fons, 2019),而忽视了林木大小分化在森林生态系统功能中的重要作用。
4.2 采伐干扰对生产力稳定性的影响
采伐可显著降低林分密度,林分密度下降会减少物种多样性(Clarosetal., 2008; 李瑞霞等, 2012),降低林木大小分化程度,从而间接降低生产力稳定性。虽然林分密度下降在一定程度上通过减少林木竞争、降低生产力年际波动来提高生产力稳定性,但不足以抵消其通过影响物种多样性和林木大小分化而间接产生的对生产力稳定性的负向效应(Loreauetal., 2013)。
土壤条件作为非生物因素的重要组分,是森林生长的主要驱动力(Quesadaetal., 2012)。林分采伐会扰动土壤,同时提高林内透光度,从而改变土壤水分和温度,加快凋落物分解,改变土壤中生物群落的组成及土壤温度、湿度等理化性质状况,进而使土壤养分变化(Dangetal., 2018; Fayolleetal., 2012)。本研究表明采伐后土壤养分含量下降,而土壤对生产力稳定性没有直接影响,而是通过林木大小分化和物种多样性间接影响生产力稳定性。这意味着土壤养分含量下降会引起森林物种多样性变化,并降低林木大小分化的复杂性(Clarosetal., 2008; Fyllasetal., 2009; Alietal., 2019),同时物种在干扰后重新生长时,生产力会在很大程度上受土壤养分限制(Alietal., 2019; Lebaueretal., 2008),进而对森林生产力稳定性造成影响。Sande等(2017)发现南美半落叶林采伐后的土壤条件对树木生长和更新很重要,但不会影响林木大小分化和物种多样性,这与本研究结果不同,可能与森林类型不同有关。
雷相东等(2005)发现间伐后森林的多样性略有提高,其研究对象处于长白落叶松(Larixolgensis)、 鱼鳞云彬(Piceajazoensis)、臭冷杉(Abiesnephrolepis)混交林的演替前中期。本研究中针阔混交林属近熟林(魏安然等, 2019),处于演替阶段后期,物种多样性已相对稳定,采伐干扰并未直接引起物种多样性变化。林分密度、林木大小分化和物种多样性与生产力稳定性显著相关。采伐强度可通过直接影响林分密度和林木大小分化进而影响生产力稳定性,或通过间接影响物种多样性而影响生产力稳定性。
5 结论
对吉林蛟河地区针阔混交林,利用结构方程模型探讨了采伐干扰对林分生产力稳定性的影响。表明采伐强度对生产力稳定性没有直接影响,与生产力稳定性直接相关的因素是林分密度、林木大小分化和物种多样性,采伐通过调整林分密度和林木大小分化来间接影响生产力稳定性。因此,在制定森林经营管理方案时,要充分考虑到抚育采伐强度对林分密度和林木大小分化的影响,进而提高森林生产力稳定性,促进森林可持续经营管理。
猜你喜欢
今日农业(2022年14期)2022-09-15
清华金融评论(2022年4期)2022-04-13
昆明医科大学学报(2022年1期)2022-02-28
大自然探索(2021年7期)2021-09-26
农民致富之友(2018年12期)2018-06-29
安徽农学通报(2017年6期)2017-04-11
绿色科技(2017年3期)2017-03-14
安徽农学通报(2014年18期)2014-10-23
安徽农学通报(2014年9期)2014-06-23
古代文明(2014年1期)2014-02-23
