
不同贮藏温度对西州密25号哈密瓜果实软化的影响
2022-06-15 02:29阿塔吾拉铁木尔张连文谢晓定
新疆农业科学 2022年5期
徐 斌,阿塔吾拉·铁木尔,张 婷,潘 俨,张连文,谢晓定
(1.新疆农业科学院农产品贮藏加工研究所,乌鲁木齐 830091;2. 天津商业大学,天津 300134;3. 新疆工程学院,乌鲁木齐 830023)
0 引 言
【研究意义】西州密25号哈密瓜口感优良、风味独特[1]。西州密25号哈密瓜属于典型的呼吸跃变型果实,采后在不适宜的温度条件下贮藏极易引起果实组织软化,口感变差,抗病性降低,容易腐烂变质,影响商品价值[2],研究贮藏温度对西州密25号哈密瓜软化的影响,对新疆脆肉型厚皮甜瓜贮藏保鲜有重要意义。【前人研究进展】软化是果实在成熟及衰老过程中的典型特征,伴随一系列的生理变化,包括呼吸强度增加,细胞壁降解酶活性升高,细胞壁结构物质的降解等[3,4]。研究表明,果实在采后贮藏期间硬度的变化与细胞壁多糖的溶解度和解聚密切相关,是由多种细胞壁降解酶促使细胞壁多糖发生降解而引起细胞壁结构发生变化所致,已在梨、杏子等果实中得到验证[5-8]。目前,关于哈密瓜果实采后软化的研究多集中在86-1、新密3号等品种[9-11]。【本研究切入点】西州密25号哈密瓜作为近年来西部地区外销量较大的品种,鲜见关于贮藏温度对其果实采后软化影响的相关报道。需研究不同贮藏温度环境下西州密25号哈密瓜果实硬度与果肉细胞壁降解酶活性变化规律。【拟解决的关键问题】以哈密瓜西州密25号为试材,采后贮藏在不同温度(0、5、10和15℃)环境条件下,定期测定果实硬度、呼吸强度及细胞壁降解酶(β-Gal、PG、PE、Cx)的活性,分析温度对哈密瓜果实采后软化的影响,以及果实呼吸强度与软化之间的关系,为西州密25号哈密瓜贮藏温度的选择及果实软化调控提供理论参考。
1 材料与方法
1.1 材 料
1.1.1 哈密瓜
用西州密25号哈密瓜采自吐鲁番鄯善地区,露地直播。于雌花开后38~42 d(商品成熟期)采摘,单果平均质量2.0 kg,中心可溶性固形物含量16%~18%。采摘时留T型果柄,剔除病果、伤果、畸形果,挑选果形大小、色泽一致的新鲜果实,在瓜地阴凉处放置2 h去除田间热后运往实验室备用。
1.1.2 主要试验仪器
TMS-Pro物性测试仪(美国FTC公司),分析软件为Texture Exponent 32。酶标分析仪(Rayto RT-6100),检测试剂盒由乌鲁木齐瑞康达医疗器械有限公司提供。
1.2 方 法
1.2.1 样品取样
在瓜体赤道部位横向切取厚度约50 mm环形瓜块,采用直径15 mm打孔器在瓜块上取圆柱形样品,平行截取20 mm长度用于TPA硬度测定,其余部分随机称取50 g果肉切成约10 mm正方体小块,用液氮速冻后置于-80℃冰箱保存用于检测酶的活性。取样时间分别是采样当天(0 d,取6个样本量的平均值作为初始值),每隔5 d取样测定1次指标,取样7次。图1
1.2.2 指标测定1.2.2.1 硬度
采用TMS-Pro物性测定仪,测定为TPA模式,用P/75探头下压测试。方法参照刘翔等[12]:预压速度2 mm/s,压缩速度1 mm/s,压后回升速度2 mm/s,回升距离8 cm,触发力0.1 N,试样受压变形量为30%。每个处理每次测定3个果实,每个果实重复3次。
1.2.2.2 呼吸强度
方法参照LI等[13],采用JFQ-315OH果蔬呼吸测定仪。气体流速为0.9 L/min。呼吸强度采用以下公式计算,每处理用果实6个,重复3次,为mg CO2/(kg·h)。

1.2.2.3 细胞壁降解酶活性
多聚半乳糖醛酸酶(简称PG)和果胶酶(简称PE)活力测定参照张鹏龙等[14]方法,采用3,5-二硝基水杨酸法在540 nm波长处测定反应混合液释放还原糖的量。PG、PE分别以每克组织1 min催化多聚半乳糖醛酸和果胶产生1 μg底物为1个酶活力单位(U)。纤维素酶(简称Cx)活性测定参照何俊瑜等[15]方法测定。β-半乳糖苷酶(简称β-Gal)活性测定参照HOU Yuan-yuan等[16]方法。
1.3 数据处理
利用Excel2007软件统计数据,计算平均值及标准差;用Sigmaplot12.0软件绘制折线图;数据之间的差异性通过SPSS21.0软件进行单因素方差分析,利用Duncan两两比较进行差异显著性分析;各项指标数据之间的相关性通过SPSS21.0软件进行双变量相关,Pearson相关系数、双侧显著性检验法检验。
2 结果与分析
2.1 不同贮藏温度对果肉硬度的影响
研究表明,西州密25号哈密瓜果肉硬度随着贮藏时间延长呈逐渐下降趋势。贮藏前5 d,各温度条件下果肉硬度下降不明显,且相互间差异不显著。贮藏第10 d,各温度条件下果肉硬度差异极显著(P<0.01),0℃条件下果肉硬度下降至35.1 N,是初始值的88% ,15℃条件下果肉硬度下降至22.0 N,是初始值的55%。各取样点测得果肉硬度均存在极显著差(P<0.01)。贮藏第25 d,0℃条件下果肉硬度表现出较低水平,显著低于5和10℃条件下果肉硬度。贮藏第30 d,0℃条件下果肉硬度下降至7.4 N,是初始值的19%,5、10℃条件下果肉硬度分别下降至12.6、9.6 N,是初始值的31%、24%,15℃条件下果肉硬度下降至4.7 N,是初始值的12%。不同温度对果肉硬度影响具有显著性差异,温度越低果肉硬度下降速率越慢。贮藏20 d后,0℃条件下果肉硬度下降速率明显高于5和10℃,这与果实出现冷害有关,贮藏第25 d,果实表面出现褐色凹陷斑点,移至室温后果肉出现水浸状,是哈密瓜冷害症状。图2

注:不同大写字母表示实验数据间具有显著性差异(P<0.05),不同小写字母表示实验数据间具有极显著性差异(P<0.01),下同
2.2 不同贮藏温度对果实呼吸强度的影响
研究表明,在30 d贮藏期内,果实呼吸强度总体呈现先升高后降低的趋势,即出现了明显的呼吸高峰。0、5和10℃条件下果实呼吸高峰均出现在贮藏第20 d,15℃条件下果实呼吸高峰出现在贮藏第15 d,相对低温可以延缓果实呼吸强度峰值的出现。不同温度对西州密25号哈密瓜果实采后呼吸强度影响具有显著性差异,0℃条件下果实呼吸强度峰值为30.53 mg CO2/(kg·h),显著低于其他3个温度,并且在整个贮藏期内呼吸强度平均值最低。15℃条件下果实呼吸强度峰值为36.45 mg CO2/(kg·h),是0℃条件下的1.2倍,在整个贮藏期内呼吸强度平均值最高。5和10℃条件下果实呼吸强度峰值分别为35.64 mg、35.44 mg CO2/(kg·h),两者差异不显著,且在整个贮藏期内变化趋势相似,平均值相近。图3

图3 不同温度下果实呼吸强度变化Fig.3 The effect of different temperature on fruit respiration intensity
2.3 不同贮藏温度对果肉β- Gal活性的影响
研究表明,在整个贮藏期内,不同温度条件下β- Gal活性总体呈升高趋势,其中15℃条件下果肉β- Gal活性出现先升高后降低的变化,峰值出现在第15 d,活性值为0.96 U,是初始值的1.5倍。与初始值相比,5和10℃条件下果肉β- Gal活性在贮藏前5 d变化不明显,贮藏第10 d开始逐渐升高,贮藏至30 d,分别升高至0.87、0.80 U,是初始值的1.36倍和1.25倍。0℃条件下果肉β- Gal活性在贮藏第10 d下降至0.60 U,贮藏第15 d迅速升高至0.75 U,之后呈缓慢上升趋势,贮藏第30 d升高至0.86 U,是初始值的1.3倍。贮藏前5 d,0℃条件下果肉β- Gal活性与其他3个温度条件下果肉β-Gal活性存在显著性差异。贮藏第5至25 d,15℃条件下果肉β- Gal活性与其他3个温度条件下果肉β-Gal活性存在显著性差异。贮藏第10~30 d,0℃条件下果肉β- Gal活性与5℃条件下果肉β- Gal活性差异不显著。贮藏第30 d,0和5℃条件下果肉β- Gal活性值略高于10和15℃条件下果肉β- Gal活性值。图4

图4 不同温度下果肉β- Gal活性变化Fig.4 The effect of different temperature on pulp β-Gal activity
2.4 不同贮藏温度对果肉PG活性的影响
研究表明,在整个贮藏期内,0℃条件下果肉PG活性呈先降低后逐渐升高趋势,在贮藏第5 d出现最低值0.315 U,且始终与其他3个温度条件下果肉PG活性存在极显著差异,表现为贮藏25 d时间内,果肉PG活性极显著低于其他3个温度条件下果肉PG活性。5、10和15℃条件下果肉PG活性在贮藏第5 d变化不大,基本保持初始值的水平,在整个贮藏期内均呈先升高后降低趋势,5℃条件下果肉PG活性峰值出现在第15 d,活性值为0.564 U,10℃条件下果肉PG活性峰值出现在第25 d,活性值为0.653 U,15℃条件下果肉PG活性峰值出现在第15 d,活性值为0.736 U,极显著高于其他3个温度条件下果肉PG活性。在贮藏10 d内,5、10和15℃条件下果肉PG活性之间不存在显著性差异,贮藏第10 d至30 d,5℃条件下果肉PG活性极显著低于10和15℃条件下果肉PG活性。贮藏25 d时间内,温度越高果肉PG活性越强,且0℃条件下果肉PG活性始终低于初始值,贮藏25 d后,0℃条件下果肉PG活性出现升高趋势,贮藏第30 d升高至0.509 U,显著高于5℃条件下果肉PG活性,果实贮藏前期0℃可有效抑制果肉PG活性升高。图5

图5 不同温度下果肉PG活性变化Fig.5 The effect of different temperature on pulp PG activity
2.5 不同贮藏温度对果肉PE活性的影响
研究表明,在整个贮藏期内,4个温度条件下果肉PE活性随时间变化比较复杂,有升有降,但总体呈上升趋势。0和5℃条件果肉PE活性在贮藏第5 d分别下降至306.85 、300.58 U,比初始值分别下降了23.93、30.20个活力单位,之后开始逐渐升高,0℃条件下果肉PE活性在贮藏第25 d出现峰值为567.33 U。贮藏第30 d,0℃条件果肉PE活性值为490.64 U,是初始值的1.5倍,5℃条件果肉PE活性值为553.99 U,是初始值的1.7倍。10℃条件下果肉PE活性呈先升高后下降再逐渐升高的变化趋势,贮藏第15 d至25 d,10℃条件下果肉PE活性高于其他3个温度条件下果肉PE活性,并且在贮藏第25 d达到峰值648.26 U,是初始值的1.9倍。15℃条件下果肉PE活性在贮藏第5 d为576.08 U,显著高于相同贮藏时间其他3个温度条件下果肉PE活性,之后呈逐渐下降趋势,在贮藏第25~30 d再次出现升高,第30 d升高至572.86 U,是初始值的1.7倍。温度越高果肉PE活性平均值越大。图6
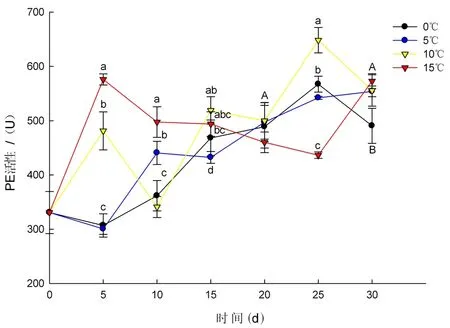
图6 不同温度下果肉PE活性变化Fig.6 The effect of different temperature on pulp PE activity
2.6 不同贮藏温度对果肉Cx活性的影响
研究表明,在整个贮藏期内,4个温度条件下果肉Cx活性均呈先升高后下降的变化趋势,出现明显的活性峰值。其中,0℃条件下果肉Cx活性峰值出现在第20 d,为2 899.49 U,是初始值的1.27倍;5℃条件下果肉Cx活性峰值出现在第25 d,为2 946.96 U,是初始值的1.30倍;10℃条件下果肉Cx活性峰值出现在第25 d,为3 126.01 U,是初始值的1.37倍;15℃条件下果肉Cx活性峰值出现在第15 d,为3 182.65 U,是初始值的1.40倍。表明温度越高果肉Cx活性峰值越大。贮藏5 d,4个温度条件下果肉Cx活性没有显著差异。贮藏10 d,15℃条件下果肉Cx活性显著高于其他3个温度条件下果肉Cx活性。贮藏第20 d,0和5℃条件下果肉Cx活性没有显著差异,10和15℃条件下果肉Cx活性没有显著差异。贮藏第25 d,10℃条件下果肉Cx活性显著高于其他3个温度条件下果肉Cx活性。贮藏第30 d,5和10℃、15℃条件下果肉Cx活性之间没有显著差异,0℃条件下果肉Cx活性极显著低于其他3个温度条件下果肉Cx活性,且与初始值相近。相对低温可抑制果肉Cx的活性。图7
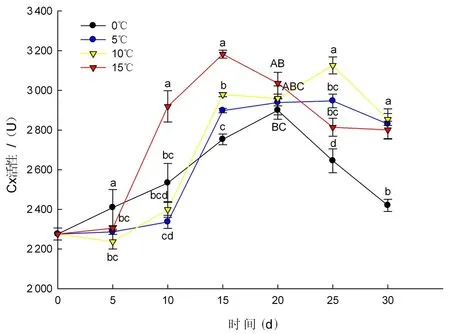
图7 不同温度下果肉Cx活性变化Fig.7 The effect of different temperature on pulp Cx activity
2.7 果实硬度与呼吸强度、细胞壁降解酶活性间相关性
研究表明,果肉硬度与呼吸强度呈负相关,与果胶酶、纤维素酶、β-半乳糖苷酶、多聚半乳糖醛酸酶活性呈极显著负相关(P<0.01);呼吸强度与果胶酶活性呈正相关,与β-半乳糖苷酶活性呈显著正相关(P<0.05),与纤维素酶、多聚半乳糖醛酸酶活性呈极显著正相关(P<0.01);果肉果胶酶与纤维素酶、β-半乳糖苷酶活性呈极显著正相关(P<0.01),与多聚半乳糖醛酸酶活性呈显著正相关(P<0.05);果肉纤维素酶与β-半乳糖苷酶、多聚半乳糖醛酸酶活性呈极显著正相关(P<0.01);果肉β-半乳糖苷酶与多聚半乳糖醛酸酶活性呈极显著正相关(P<0.01)。西州密25号哈密瓜采后果实硬度的下降与果胶酶、纤维素酶、β-半乳糖苷酶、多聚半乳糖醛酸酶活性的升高具有极显著正相关关系。呼吸强度的变化与果肉4种细胞壁降解酶活性变化存在显著或极显著正相关关系。果肉果胶酶、纤维素酶、β-半乳糖苷酶、多聚半乳糖醛酸酶活性相互间具有显著正相关关系。表1
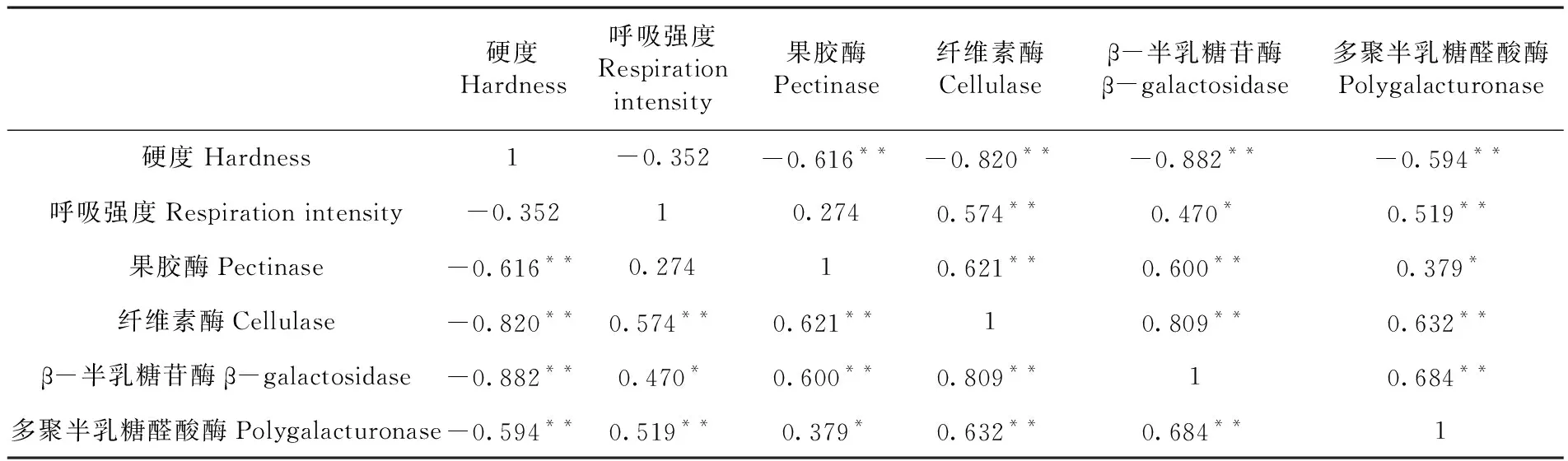
表1 果肉硬度与呼吸强度、细胞壁降解酶活性相关性(R)矩阵Table 1 Correlation(R) matrix between fruit firmness and respiration intensity, enzymes activities of cell wall
3 讨 论
采后呼吸强度及细胞壁结构的改变是引起果实硬度变化的主要原因[17-19]。研究发现,西州密25号哈密瓜果实存在明显的呼吸高峰,与15℃相比,0、5、10℃能延迟呼吸高峰的出现,并且0℃条件下果实呼吸强度平均值及峰值显著低于其他3个温度,说明低温能有效抑制果实的呼吸强度。然而,0℃条件下贮藏20 d后,果实硬度迅速下降,并且出现冷害症状,因此,0℃并不适宜西州密25号哈密瓜贮藏。果实细胞壁的主要成分由果胶物质、纤维素和半纤维素组成,在贮藏过程中,果胶物质及纤维素的降解导致胞间层结构发生变化,细胞壁结构解体,从而引起果实的软化[20]。苹果在贮藏过程中果实硬度下降、质地变软,伴随着细胞壁降解酶活性升高,原果胶含量下降及可溶性果胶含量上升[21]。细胞壁降解酶主要有果胶酶、β-半乳糖苷酶、多聚半乳糖醛酸酶和纤维素酶,这些降解酶在果实软化过程中具有不同的作用,例如果胶酶可去除果胶分子链上半乳糖醛酸羧基上的酯化基团,增加果胶在水中的溶解度;β-半乳糖苷酶可通过降解具有支链的多聚醛酸使细胞壁组分变得不稳定,促使果胶降解或溶解;多聚半乳糖醛酸酶能够水解细胞壁果胶中多聚半乳糖醛酸多糖主链的α-1,4-糖苷键,生成半乳糖醛酸和低聚半乳糖醛酸,导致细胞壁成分和结构的变化;纤维素酶能促进纤维素的降解,导致细胞壁“经纬结构”松散[22-30]。研究发现,随着贮藏时间的延长,不同温度条件下果肉细胞壁降解酶的活性存在显著差异,表现为15℃条件下,果肉β-半乳糖苷酶、多聚半乳糖醛酸酶和纤维素酶活性峰值及平均值均高于其他3个温度条件下的活性峰值和平均值,且温度越高,酶的活性越强,温度能够调控这3种酶的活性。在贮藏前期,0℃能够有效抑制β-半乳糖苷酶、多聚半乳糖醛酸酶活性,贮藏后期,0℃条件下2种酶活性出现升高趋势,可能与果实出现冷害有关,有待进一步研究。不同温度对果胶酶的影响比较复杂,数据分析显示,随着贮藏时间的延长,果胶酶总体呈逐渐上升的变化,但这种变化受温度影响没有表现出规律性,果胶酶是由多种酶组成的复合酶,其中每一种酶的结构及受环境因素的影响均不相同,温度对果胶酶的调控机理也有待进一步研究。相关性分析可知,果肉硬度与4种酶的活性呈极显著负相关(P<0.01),4种酶活性的升高与果肉硬度的下降具有极显著的正相关关系。温度可通过调控西州密25号哈密瓜果实呼吸强度及细胞壁降解酶的活性来影响果实的后熟及软化,但果实软化是一个复杂的过程,仍需结合细胞壁成分变化、细胞超微结构等来进一步阐明。
4 结 论
5℃适合西州密25号哈密瓜果实贮藏。西州密25号哈密瓜果实硬度与呼吸强度呈负相关、与β-Gal、PG、PE和Cx活性呈极显著负相关。呼吸强度与纤维素酶、多聚半乳糖醛酸酶呈极显著负相关,与β-半乳糖苷酶呈显著负相关。
猜你喜欢
中草药(2022年20期)2022-11-15
食品科学(2022年20期)2022-10-31
农产品市场周刊(2022年2期)2022-03-31
农产品市场周刊(2022年2期)2022-03-09
江苏农业科学(2021年8期)2021-05-26
三农资讯半月报(2020年18期)2020-10-14
炎黄地理(2019年5期)2019-10-20
丝路视野(2018年15期)2018-05-14
创新作文(1-2年级)(2017年5期)2017-12-07
饮食科学(2016年9期)2016-11-18
