
淫羊藿苷通过激活转录共激活因子预防大鼠激素性股骨头坏死进展的实验研究
2022-06-10 08:16:54何嘉怡迟瑞敏贺毅蔡卓许涛郭风劲叶亚平
骨科 2022年3期
何嘉怡 迟瑞敏 贺毅 蔡卓 许涛 郭风劲 叶亚平
糖皮质激素相关性股骨头坏死是大剂量或长期使用糖皮质激素的严重副作用[1]。随着骨坏死进展,股骨头塌陷并导致髋关节骨性关节炎和疼痛,其致残率高并严重影响病人生活质量[2]。晚期股骨头坏死病人最有效的治疗是全髋关节置换,但其费用高昂且存在晚期假体松动和磨损等并发症。如何在股骨头坏死早期有效预防骨坏死进展,加速骨坏死的康复治疗已成为临床上亟待解决的关键问题[3-4]。
近期一项多中心的随机双盲对照临床试验表明,复方仙灵骨葆胶囊(淫羊藿含量占70%[5])可有效降低接受糖皮质激素治疗病人的股骨头坏死发生率,但其分子机制不清[6]。有研究[7]显示,淫羊藿苷可通过抑制血管内血栓形成及血管外脂质沉积来降低股骨头坏死的发生率。有学者将淫羊藿苷与聚乳酸羟基乙酸/磷酸三钙纳米纤维支架结合,促进股骨头坏死病人的骨缺损修复[8]。据报道转录共激活因子(TAZ)能促进间充质干细胞(BMSCs)的成骨分化,TAZ 能激活Runt 相关转录因子2(runt-related transcriptional factor 2,Runx2)依赖的成骨分化基因表达并抑制抗过氧化物酶体增殖物激活受体(peroxisome proliferator-activated receptor γ,PPARγ)依赖的成脂分化基因表达,但是TAZ在淫羊藿苷调控激素性股骨头坏死中的作用尚无研究报道[9]。本研究拟通过体内实验和体外实验,从分子水平去研究淫羊藿苷对激素性股骨头坏死的保护作用及其相关机制,为股骨头坏死的治疗提供新的作用靶点,为有效降低激素性股骨头坏死的发生率并加速早期股骨头坏死病人的康复提供有价值的参考。
材料与方法
一、实验材料
淫羊藿苷(Sigma,美国),地塞米松(碧云天生物,中国),甲强龙(辉瑞,美国),siRNA-TAZ 和空白siRNA(广州瑞博生物科技有限公司,中国),引物合成(擎科生物,中国),PPARγ抗体(Abcam,美国),结缔组织生长因子(connective tissue growth factor,CTGF)抗体(Abcam,美国),TAZ 抗体(CST,美国),Runx2 抗体(博士德,中国),GAPDH 抗体(博士德,中国),辣根过氧化物酶偶联二抗(博士德,中国),骨钙蛋白(osteocalcin,OCN)抗体(博士德,中国),血小板-内皮细胞粘附分子(Platelet endothelial cell adhesion molecule-1,PECAM-1/CD31)抗体(Santa Cruz,美国),血管内皮生长因子(vascular endothelial growth factor,VEGF)抗体(Santa Cruz,美国),生物素化羊抗兔/小鼠抗体(博士德,中国)。蛋白定量检测试剂盒(博士德,中国),ECL化学发光试剂盒(Thermo 公司,美国),Trizol 抽提液(Invitrogen,美国),总RNA 提取试剂盒(Toyobo,日本),cDNA 合成试剂盒(Toyobo,日本),SYBR Green 荧光定量PCR 试剂盒(Toyobo,日本),CCK-8 试剂盒(博士德,中国),Annexin V-FITC/PI 试剂盒(碧云天生物,中国)。成年雄性SD 大鼠由华中科技大学同济医学院实验动物中心提供。
二、实验方法
(一)大鼠BMSCs的分离、培养与分组
所有动物操作均经华中科技大学同济医学院实验动物伦理委员会批准。大鼠骨髓间充质干细胞(rBMSCs)取自3 月龄SD 大鼠的股骨。取出股骨冲出骨髓,贴壁培养得到rBMSCs。本实验使用第3代rBMSCs。
分组如下:①对照组;②地塞米松组(10-6mol/L);③联合干预组[地塞米松(10-6mol/L)+不同浓度淫羊藿苷(1×10-8mol/L,1×10-7mol/L,1×10-6mol/L,1×10-5mol/L)]。
(二)细胞增殖检测
使用CCK-8 试剂盒检测细胞增殖情况。将rBMSCs 接种到96 孔板中(5×103个/孔)。24 h 后根据上述分组给药。培养72 h 后,每孔加入10 μL CCK-8 溶液,37 ℃黑暗孵育1 h。最后,使用IMAX190 光度计读取450 nm 处的吸光度。通过比较光密度值(OD)来评价各组细胞增殖情况。
(三)细胞凋亡检测
使用Annexin V-FITC/PI 试剂盒检测细胞凋亡情况。按上述分组给药72 h后收集细胞。加入Annexin V-FITC/PI 染色,再使用流式细胞仪进行分析。Annexin V+/PI-和Annexin V+/PI+的细胞分别被认为是早期和晚期的凋亡细胞。
(四)成骨和成脂分化试验
成骨培养基由DMEM/F12、10%FBS、10 mmol/L β-甘油磷酸酯、10-8mol/L 地塞米松和50 μg/mL 抗坏血酸组成;成脂培养基由DMEM/F12、10%FBS、0.5 mmol/L 异丁基甲基黄嘌呤和10 mg/mL 胰岛素组成。接种细胞后每两天更换培养基,21 d 后进行茜素红-S 染色(成骨分化)和油红-O 染色(成脂分化)[10-11]。
(五)小干扰RNA转染试验
将第3 代rBMSCs 以5×104个/孔的密度种植在6 孔板中,当细胞融合达到50%时,经脂质体将siRNA 转染到rBMSCs 中以沉默TAZ 表达[10]。48 h 后,采用地塞米松(1×10-6mol/L)+淫羊藿苷(1×10-7mol/L)处理上述细胞72 h,以了解rBMSCs在地塞米松和淫羊藿苷联合作用下,TAZ 蛋白的表达是否可影响其下游靶基因(CTGF)表达及细胞成骨分化(Runx2)。
(六)蛋白免疫印迹
使用Western Blot 法进行蛋白表达定量分析。提取蛋白,BCA 法测定蛋白浓度,SDS-PAGE 电泳,转膜,孵育一抗(PPARγ、TAZ、CTGF、Runx2、GAPDH),孵育二抗,使用ECL化学发光试剂盒观察蛋白条带。
(七)实时荧光定量PCR
使用实时荧光定量PCR 进行基因表达定量分析。Trizol提取总RNA,按cDNA合成试剂盒操作步骤逆转录得到cDNA,然后扩增处理,循环条件如下:95 ℃预变性30 s,然后95 ℃变性5 s和60 ℃下退火30 s,共循环40 次。Runx2 引物序列:上游CTACTCTGCCGAGCTACGAAAT,下 游 TCTGTCTGTGCCTTCTTGGTTC;ALP 引 物 序 列 :上 游AGGGAAGGGTCAGTCAGGTT,下 游CCTGGACCTCATCA-GCATTT;OPN 引物序列:上游CAAGGACCAACTACAACCA,下 游 GGAG - ACAGGAGGCAAGG;GAPDH 引 物 序 列:上 游 AACGACCCCTTCATTGACC - TC,下 游CCTTGACTGTGCCGTTGAACT。
(八)动物模型制备
选用12 周龄、体重250~300 g 的SD 大鼠52 只,随机分为三组。对照组(12只)不进行特殊处理;甲强龙组(20 只)腹腔注射脂多糖(LPS,0.2 mg·kg-1),然后肌肉注射甲强龙(MP,40 mg·kg-1·d-1)连续3 d;联合干预组(20只)腹腔注射LPS并肌肉注射MP,在首次注射MP后连续灌胃淫羊藿苷(100 mg·kg-1·d-1),直至动物被处死[12]。
(九)免疫组化染色
造模6 周后对大鼠进行安乐死处理并取出股骨,使用10%EDTA 溶液脱钙3 周,包埋于石蜡中。将股骨头沿冠状面以5 μm 厚度切片。使用苏木精和伊红(HE)染色评估小梁结构;应用免疫组化学染色方法检测股骨头OCN、TAZ、CD31、VEGF 蛋白表达情况。使用Image-Pro Plus 软件分析。在镜下至少选择5 个随机区域,根据每张图片的累积光密度值(IOD)定量阳性染色,并测量相应区域面积(area),平均光密度=IOD/area。
(十)TUNEL细胞凋亡检测
使用TUNEL 试剂盒检测骨细胞凋亡情况。以胞核为褐色或胞浆中有褐色颗粒的细胞为阳性细胞。使用Image-Pro Plus软件分析得到平均光密度,用以衡量各组细胞凋亡率。
(十一)股骨头Micro-CT检查
造模6周后对大鼠进行安乐死处理并收集其股骨头,使用Micro-CT 扫描。采用直接三维测量方法计算骨密度(bone mineral density,BMD)、骨体积/组织体积(bone volume per tissue volume,BV/TV)、骨小梁数(trabecular number,Tb.N)、骨小梁间距(trabecular separation,Tb.Sp)和骨小梁厚度(trabecular thickness,Tb.Th)。
(十二)血管造影
造模6 周后麻醉大鼠,分离腹主动脉和静脉。心脏注射肝素化盐水后,通过远端腹主动脉注射Microfil 造影剂,直到观察到其从腹腔静脉持续流出。随后,上述大鼠4 ℃保存过夜,取双侧股骨头,4%多聚甲醛固定,10% EDTA 溶液脱钙。最后,按照上述方法对样本进行Micro-CT 扫描,利用内置软件重建并计算股骨头血管体积。
三、统计学分析
采用SPSS 13.0(IBM 公司,美国)进行统计分析。计量数据以均数±标准差(±s)表示,两组差异用Student-t 检验确定,多组间的差异使用单因素方差分析确定,然后使用Bonferroni检验来具体比较两组之间的差异。P<0.05认为差异有统计学意义。
结 果
一、淫羊藿苷可有效削弱地塞米松对rBMSCs的增殖抑制和凋亡促进作用
研究结果表明地塞米松抑制rBMSCs增殖,促进rBMSCs凋亡(图1 a~c)。而采用不同浓度淫羊藿苷处理后,地塞米松对rBMSCs的上述作用均得到不同程度的削弱,其中10-7mol/L浓度的淫羊藿苷在削弱地塞米松对rBMSCs 的增殖抑制和促进凋亡方面具有最佳的效果(图1 a~c)。
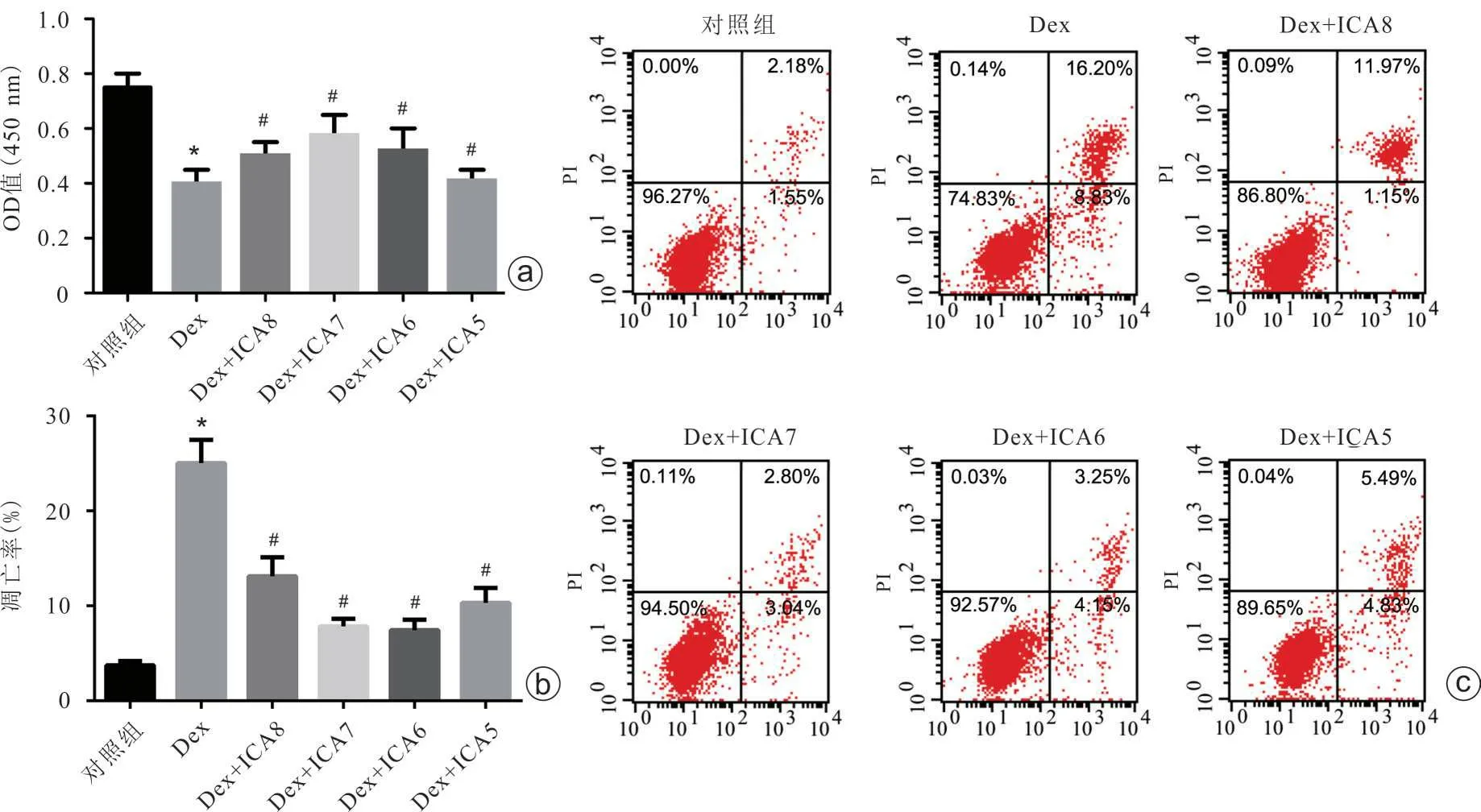
图1 地塞米松和不同浓度淫羊藿苷对rBMSCs增殖和凋亡的影响 a:CCK-8法检测rBMSCs增殖情况;b、c:Annexin V-FITC/PI双染法流式细胞术检测rBMSCs 凋亡情况[Dex:地塞米松;ICA8:淫羊藿苷(1×10-8 mol/L);ICA7:淫羊藿苷(1×10-7 mol/L);ICA6:淫羊藿苷(1×10-6 mol/L);ICA5:淫羊藿苷(1×10-5 mol/L);与对照组比较,*P<0.05;与地塞米松组比较,#P<0.05]
二、淫羊藿苷可有效改善地塞米松对rBMSCs成骨分化的抑制作用和对其成脂分化的促进作用
本研究显示,与对照组比较,地塞米松组中rBMSCs 的成骨分化被明显抑制,而联合干预组中rBMSCs 成骨分化增强(图2 a);此外,与对照组比较,地塞米松组中成骨分化相关基因Runx2 和ALP表达水平明显下降,OPN 表达水平无变化;而与地塞米松组比较,联合干预组中Runx2、ALP和OPN表达水平明显增加(图2 b)。此外,地塞米松可显著增强成脂分化关键蛋白PPAR-γ表达,而淫羊藿苷可下调该基因在蛋白水平的表达(图2 c)。油红O 染色显示地塞米松可促进rBMSCs成脂分化,而淫羊藿苷可削弱地塞米松对rBMSCs成脂分化的促进作用(图2 d)。

图2 地塞米松和不同浓度淫羊藿苷对rBMSCs 分化的影响 a:茜素红-S 染色检测rBMSCs 成骨分化,比例尺=400 μm;b:实时检测Runx2、ALP、OPN的mRNA表达;c:Western blot 检测PPAR-γ蛋白表达;d:油红-O染色检测rBMSCs成脂分化,比例尺=400 μm[Dex:地塞米松;ICA8:淫羊藿苷(1×10-8 mol/L);ICA7:淫羊藿苷(1×10-7 mol/L);ICA6:淫羊藿苷(1×10-6 mol/L);ICA5:淫羊藿苷(1×10-5 mol/L);与对照组比较,*P<0.05;与地塞米松组比较,#P<0.05]
三、淫羊藿苷可能通过促进TAZ 表达来削弱地塞米松对rBMSCs增殖和成骨分化的影响
本研究显示地塞米松可抑制rBMSCs 中TAZ 蛋白表达,而淫羊藿苷可有效改善地塞米松对TAZ蛋白的表达抑制作用(图3 a)。使用siRNA 沉默rBMSCs 中TAZ蛋白表达,24 h后siTAZ组中TAZ蛋白表达被明显抑制(图3 b)。再采用地塞米松+淫羊藿苷处理上述细胞3 d,结果显示,地塞米松可显著抑制CTGF 及Runx2 蛋白表达;而地塞米松+淫羊藿苷可显著增加CTGF 及Runx2 蛋白表达。这提示淫羊藿苷可有效改善地塞米松对CTGF 及Runx2 蛋白的表达抑制作用。当TAZ 表达被沉默后,其下游的CTGF 蛋白和成骨分化相关蛋白Runx2 表达均发生不同程度的下降(图3 c)。本部分实验提示,地塞米松可能通过抑制TAZ 表达来抑制Runx2,而淫羊藿苷则可能通过促进TAZ 表达来达到削弱地塞米松对rBMSCs增殖和成骨分化的调控作用。
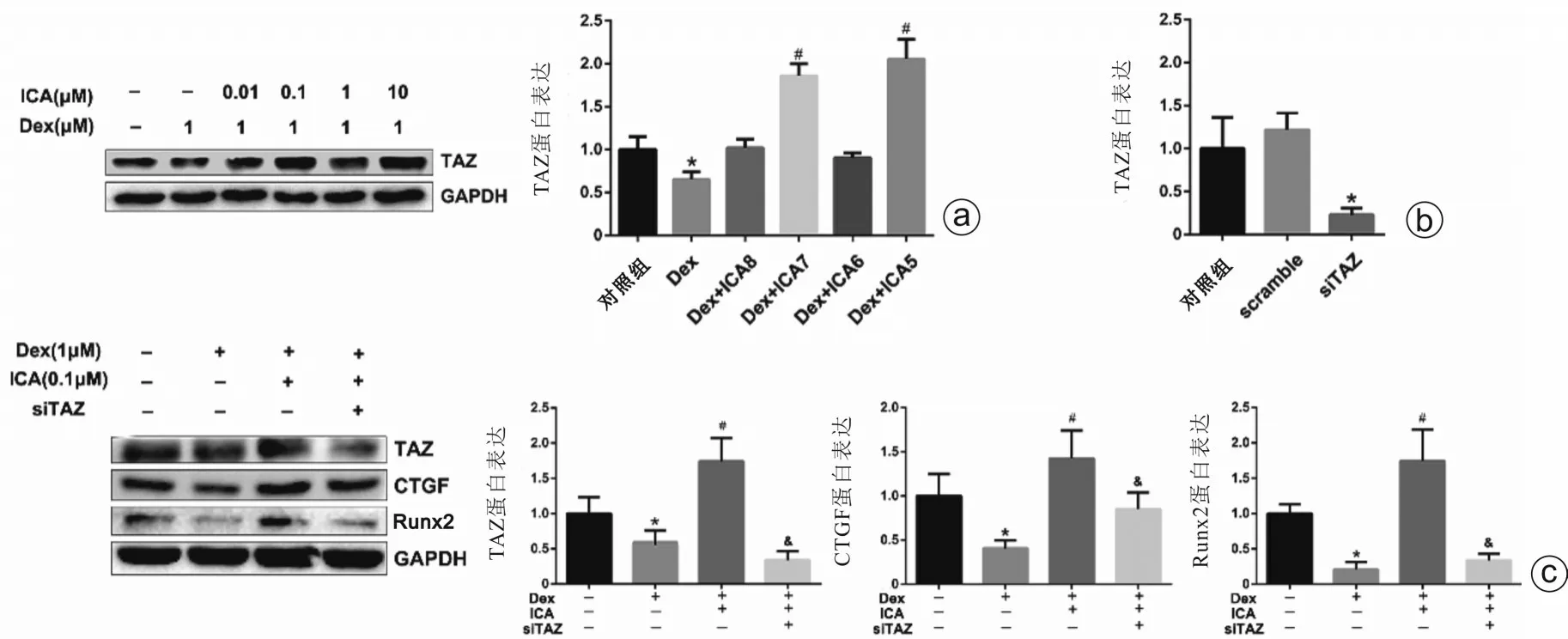
图3 Western blot 检测地塞米松和不同浓度淫羊藿苷对rBMSCs 中TAZ、CTGF、Runx2 蛋白表达的影响 a:各组中TAZ 蛋白表达情况;b:siRNA 转染rBMSCs 后TAZ 蛋白表达情况;c:siRNA 转染rBMSCs 后,各组中TAZ、CTGF、Runx2 蛋白表达情况[Dex:地塞米松;ICA8:淫羊藿苷(1×10-8 mol/L);ICA7:淫羊藿苷(1×10-7 mol/L);ICA6:淫羊藿苷(1×10-6 mol/L);ICA5:淫羊藿苷(1×10-5 mol/L);scramble:无义对照;与对照组比较,*P<0.05;与地塞米松组比较,#P<0.05;与联合干预组比较,&P<0.05]
四、淫羊藿苷能显著改善激素相关的股骨头坏死发生
HE染色结果显示,对照组中大鼠骨坏死发生率为0,甲强龙组骨坏死发生率为90%(18/20),联合干预组骨坏死发生率为35%(7/20)。甲强龙组中可见大量坏死组织及空骨陷窝,而联合干预组则少见(图4)。TUNEL 凋亡染色结果显示,与对照组比较,甲强龙组大鼠股骨头软骨下骨小梁的骨细胞凋亡明显增加(图5 a),而联合干预组大鼠骨细胞凋亡减少(图5 a)。免疫组化结果也提示,甲强龙可明显抑制OCN和TAZ蛋白表达,而联合干预组中OCN和TAZ蛋白表达均显著上调(图5 b、c)。
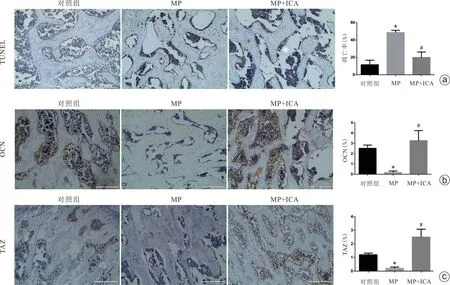
图5 大鼠股骨头TUNEL染色及OCN蛋白、TAZ蛋白表达情况,比例尺=200 μm a:TUNEL染色检测大鼠股骨头软骨下骨细胞凋亡情况;b、c:免疫组化染色示大鼠股骨头软骨下OCN和TAZ表达情况(MP:甲强龙;ICA:淫羊藿苷;与对照组比较,*P<0.05;与甲强龙组比较,#P<0.05)
Micro-CT 显示甲强龙组BMD、BV/TV、Tb.N 和Tb.Th 较对照组显著降低,同时Tb.Sp 增加,提示激素可破坏大鼠股骨头近端软骨下骨小梁结构(图6)。与甲强龙组比较,联合干预组大鼠的骨破坏显著减轻,其BMD、BV/TV、Tb.N 和Tb.Th 与甲强龙组比较显著增加,而Tb.Sp减少(图6)。
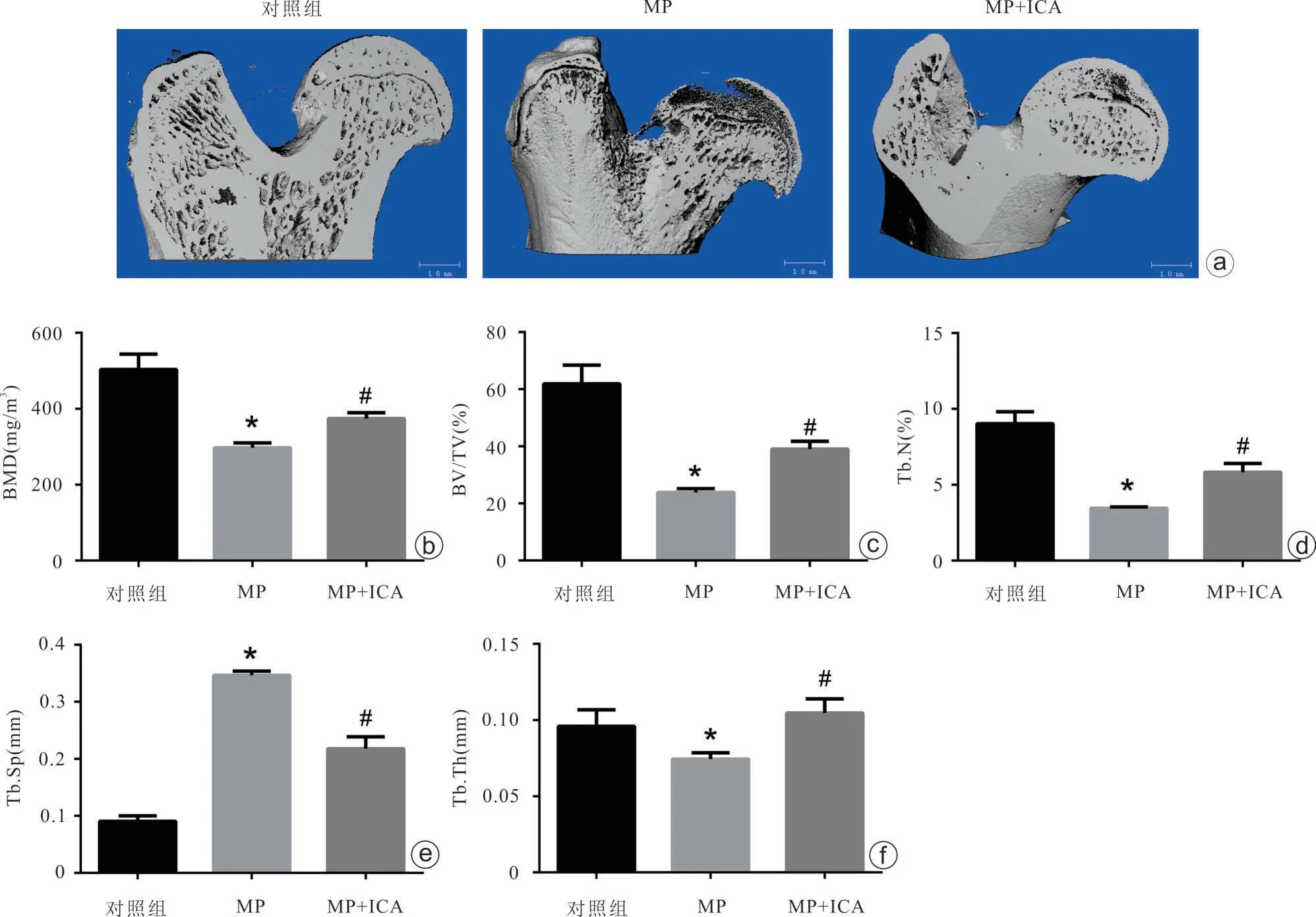
图6 Micro-CT分析股骨头微观结构 a:股骨头代表性Micro-CT图像;b~f:软骨下骨小梁微观结构参数(MP:甲强龙;ICA:淫羊藿苷;与对照组比较,*P<0.05;与甲强龙组比较,#P<0.05)
五、淫羊藿苷可有效改善激素对股骨头内血液供应的破坏作用
血管重建三维CT 显示,与对照组比较,甲强龙干预可显著降低大鼠股骨头血管体积,而联合干预组的股骨头内血管体积较甲强龙组明显增加(图7 a)。此外,通过免疫组化染色检测股骨头血管内皮细胞标志物CD31和VEGF的结果显示,甲强龙组中CD31 和VEGF 表达明显被抑制,而联合干预组中CD31 和VEGF 蛋白表达水平较甲强龙组显著增加(图7 b、c)。上述结果提示甲强龙可抑制大鼠股骨头内的血供,而淫羊藿苷可显著改善激素对大鼠股骨头血流灌注的抑制效应。
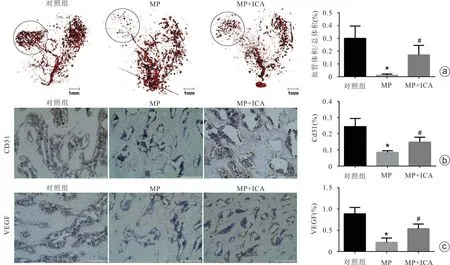
图7 大鼠股骨头血供和血管生成情况 a:大鼠股骨头血供的三维图像及血管体积与总体积的比率;b、c:免疫组化显示大鼠股骨头软骨下CD31和VEGF表达情况,比例尺=200 μm(MP:甲强龙;ICA:淫羊藿苷;与对照组比较,*P<0.05;与甲强龙组比较,#P<0.05)
讨 论
既往研究显示长期使用糖皮质激素可导致股骨头坏死发生[13-15]。激素相关性股骨头坏死病人的BMSCs 增殖和成骨分化能力均低于未使用激素对照组病人[16]。也有研究发现激素可通过降低骨形态发生蛋白-2 的表达来抑制BMSCs 的成骨分化[17]。此外,激素还能促使BMSCs由成骨分化转向成脂分化[18]。在本研究中,我们亦发现地塞米松可促进rBMSCs的凋亡和成脂分化,并抑制rBMSCs的增殖、成骨分化和钙质沉积。我们还发现了淫羊藿苷可显著逆转地塞米松的上述效应,其最佳浓度为10-7mol/L。
既往研究提示不同浓度的地塞米松对BMSCs分化具有截然相反的效应。低浓度地塞米松(10-8mol/L)能促进TAZ 表达和BMSCs 成骨分化,该作用可被糖皮质激素受体拮抗剂RU486 阻断[19]。高浓度地塞米松(10-7mol/L)却能抑制TAZ 表达和BMSCs 成骨分化[19]。本实验中,我们重点关注了高浓度地塞米松(10-6mol/L)和淫羊藿苷对rBMSCs 内TAZ表达和细胞分化的调控作用。结果显示地塞米松显著抑制了rBMSCs 中TAZ 及其下游靶基因CTGF表达,也能抑制Runx2的表达。而淫羊藿苷能削弱地塞米松对rBMSCs的作用,部分上调TAZ、CTGF和Runx2 蛋白的表达。Runx2 已被证实是控制BMSCs成骨分化最重要的转录因子[20]。而TAZ可与位于Runx2 调控区域的序列基序Pro-Pro-X-Tyr 结合,进而促进成骨分化过程的启动[21]。我们实验证实,使用siRNA-TAZ 沉默TAZ 表达后,地塞米松和淫羊藿苷组Runx2 蛋白表达显著降低。以上结果均提示,TAZ 应该是地塞米松和淫羊藿苷调控Runx2 表达和rBMSCs成骨分化的关键靶点。
HE染色是评价骨坏死的组织学金标准[22]。HE染色结果显示激素干预可显著促进大鼠股骨头软骨下骨小梁骨坏死病变进展,大量空骨陷窝被坏死骨髓包围,而淫羊藿苷能显著缓解激素导致的股骨头坏死。骨坏死的主要病理变化是骨细胞和成骨细胞的凋亡[23-25]。TUNEL 染色进一步证实,激素能明显增加骨细胞凋亡,而淫羊藿苷处理可明显缓解激素诱导的骨细胞凋亡。此外,免疫组化证实激素能抑制TAZ蛋白表达,而淫羊藿苷能消除激素对TAZ表达的影响。这些都进一步证明TAZ 在激素和淫羊藿苷调控大鼠股骨头坏死中发挥了关键作用。
Micro-CT结果也显示,激素干预6周后,大鼠股骨头软骨下骨小梁出现了明显骨丢失,其股骨头软骨下骨小梁的BMD、BV/TV、Tb.N 和Tb.Th 显著降低,Tb.Sp增加。而淫羊藿苷治疗可消除激素对大鼠股骨头的不良影响,具体表现为BMD、BV/TV、Tb.N和Tb.Th 增加,而Tb.Sp 降低。此外,我们还对大鼠股骨头内的血管分布进行了研究,结果显示激素干预使大鼠股骨头血管体积明显减小,而淫羊藿苷可显著增加大鼠股骨头血供,这提示淫羊藿苷可能通过增加股骨头血供来抑制骨坏死的进展。免疫组化进一步证实激素可抑制股骨头CD31 和子VEGF 表达,而淫羊藿苷显著消除了激素对CD31和VEGF蛋白表达的抑制作用。
综上所述,我们研究证实淫羊藿苷可以通过拮抗糖皮质激素,来调控rBMSCs 的凋亡和分化,而TAZ则可以作为地塞米松和淫羊藿苷的共同作用靶点起到调控rBMSCs的作用。此外,我们的体内实验也证实淫羊藿苷能显著逆转激素对大鼠股骨头内TAZ表达、骨和血管形成的抑制性作用,淫羊藿苷可能通过促进TAZ 蛋白表达来预防股骨头坏死的发生发展。在下一步的实验计划中,我们拟进一步构建能够缓释淫羊藿苷的生物材料对股骨头坏死病灶清除后的骨缺损部位进行移植手术,以观察其对骨坏死病灶的修复作用。总体而言,本实验对激素性股骨头坏死的早期预防和治疗意义重大,为激素性股骨头坏死提供了新的潜在治疗靶点。
猜你喜欢
食品与发酵工业(2023年21期)2023-11-26 07:50:24
江西中医药(2022年8期)2022-08-22 02:01:26
口腔医学(2021年10期)2021-12-02 02:08:00
中成药(2018年12期)2018-12-29 12:26:00
中国中医药现代远程教育(2018年22期)2018-02-09 02:12:04
湖南林业科技(2017年6期)2018-01-30 03:48:06
中成药(2017年4期)2017-05-17 06:09:49
中华老年口腔医学杂志(2016年2期)2017-01-15 14:24:47
中国病理生理杂志(2015年8期)2015-12-21 12:38:14
天津护理(2015年4期)2015-11-10 06:11:41
