
黄海水体生物发光所致离水辐亮度变化及其与固有光学性质和深度的联系
2022-06-06 10:07张雨潇陈树果刘召伟胡连波
光谱学与光谱分析 2022年6期
张雨潇,王 希,陈树果, 2, *,刘召伟,胡连波, 2
1.中国海洋大学海洋技术学院/三亚海洋研究院,山东 青岛 266100/海南 三亚 572024 2.青岛海洋科学与技术试点国家实验室,山东 青岛 266237 3.国家卫星海洋应用中心,北京 100081
引 言
机械刺激下的生物发光作为全球海洋水体普遍存在的自然现象,其定量化研究(光谱特征和发光动力学特征)无论对于海洋生态还是水下目标探测均具有重要的意义[1-4]。Gordon[5]于1984年通过一些假设对生物发光的星载传感器探测的可行性以及依赖性进行了理论上的分析。Yi等[6]对生物发光的可探测性进行深入的研究,然而这些研究都是理论上的探讨。2005年,Miller等[7]利用美国防卫气象卫星DMSP-OLS(夜间卫星)成功探测到位于印度洋的著名“牛奶海”的细菌发光现象,该项研究意义重大,虽然造成该发光的生物有机体是细菌而不是浮游生物,但其研究结果证明了海洋生物发光的实际星载传感器的可探测性,并进一步分析了星载可探测的生物发光强度的阈值范围。Moline[8]等与Oliver[9]等围绕机械刺激所致的生物发光的强度分布及其辐射传输特性展开研究,其研究结果表明,生物发光的辐射传输特性的决定因素分别是发光浮游生物的分布及水体的固有光学性质。黄海海域具有发光能力的生物有机体分布广泛,数量丰富,其中部分发光生物有较高的浓度,为所在地区的优势物种,并且生物发光现象在黄海水体普遍存在。我国黄海受长江及黄河输入的影响,其水体性质在时间和空间上都有着剧烈的变化,即黄海水体的固有光学性质在时间与空间分布上差异较大[10],从而影响生物发光的辐射传输特性。
基于机械刺激下浮游生物发光的重要意义,Moline等围绕美国新泽西州近岸水体展开了生物发光辐射传输特性的研究[8],本研究主要围绕固有光学性质差异较大的黄海,针对机械刺激下浮游生物发光的辐射传输特性展开研究。通过在黄海广泛测量的浮游生物发光强度及固有光学性质数据,模拟水下生物发光辐射传输过程,分析黄海水体浮游生物发光的辐射分布特征与季节变化趋势,并讨论黄海水体固有光学性质及光源深度对浮游生物发光光谱变化的影响。
1 实验部分
1.1 数据
1.1.1 数据来源
本研究所使用的数据包括生物发光强度(the mechanical-stimulated bioluminescence potential, MBP)以及水体固有光学性质(inherent optical properties)数据。水体固有光学性质包括吸收系数、后向散射系数和衰减系数。为了获取上述所需的研究数据,从而研究黄海水体生物发光辐射传输特性,分别于2017年12月20日至次年1月10日(冬季航次),2018年3月28日至4月16日(春季航次)、2018年7月24日至8月8日(夏季航次),搭乘“东方红2号”科学考察船在黄海海域测量了3个季节整个水体剖面的黄海海域生物发光强度数据以及固有光学性质数据,测量站位分布如图1所示。

图1 测量站位分布图
1.1.2 测量仪器
本研究所使用的测量仪器在使用前后均进行了严格的实验室定标处理。其中,生物发光分析仪(underwater bioluminescence assessment tool,UBAT)用于生物发光强度的测量。吸收系数与衰减系数采用美国 WET Labs公司研发的反射管式水体吸收衰减测量仪(spectral absorption and attenuation meter,AC-S)测量。后向散射系数采用美国WET Labs公司研发的后向散射仪(Scattering Meter,BB9)测量。使用温盐深剖面仪Seabird SBE19 PLUS 进行同步测量水体温度、盐度以及深度信息。对于AC-S,利用同步测量的温度和盐度数据进行温度和盐度校正,然后根据散射误差与相应波段散射系数之间的比例关系进行散射校正。对于BB9,使用校正后的AC-S数据进行路径补偿校正。仪器手册给出了上述仪器的具体校正方法及数据的具体处理方法,本文不再赘述[11-13]。将以上仪器安装固定在光学剖面笼中,使用绞车以0.15 m·s-1的下降速度将光学剖面笼从水面下放至海底3 m以上深度(避免仪器触底遭受损坏),从而获取水体生物发光强度和固有光学性质等从表层至底层的连续剖面测量数据。
1.1.3 水下光学深度
光学深度τ定义如式(1)所示,与几何深度z、衰减系数c以及波长有关,其中,z1为ACS测量剖面初始水深,z2为光源的几何深度,本研究所计算的为不同几何深度,不同峰值波长下的光学深度。

(1)
1.2 生物发光辐射传输模拟方法
水下生物发光所产生的离水辐射强度和光谱形状与水体衰减系数有关[14]。因此,为了揭示固有光学性质对生物发光所致离水辐亮度(Lw-bio)的幅值以及光谱变化的影响,本研究使用Hydrolight(v5.0)辐射传输软件[15-16],通过以下步骤进行辐射传输模拟,得到Lw-bio,从而构建生物发光模型。
(1)将实测浮游生物发光测量剖面划分为以1 m深度为间隔的多个水下发光源,将相应的输入数据按1 m间隔进行插值,实测数据从水下3 m起至海底3 m以上。
(2)输入参数:将实测黄海海域固有光学性质作为输入,使用Pope等所提供的纯水吸收系数[17],体积散射函数由测量的bb(λ)/b(λ)根据Petzold计算[18],bb(λ)为水体后向散射系数,b(λ)为水体总散射系数。本研究仅针对生物发光现象,因此需排除外部光源的影响,为此采用夜间模式,将太阳辐射设置为0,确保模拟所得辐亮度仅来自于生物发光。表1给出了Hydrolight辐射传输模型输入参数的配置情况。
(3)生成Lw-bio:不同生物体所产生的生物发光光谱是不同的,而对于黄海海域,引起生物发光的生物主要是甲藻,Moline等使用已知的光谱从甲藻等生物发光光谱中对生物发光谱进行了光谱重建[8]。因此,本研究使用Moline构建的光谱将MBP通过式(2)转化为不同波长下的能量形式E(W·m-3·nm-1)。其中h为普朗克常量,值为6.63×10-34J·s,c为光速,值为3×108m·s-1,P(percent of total photons)为Moline构建的光谱中光子数在不同波长下所占百分比,λ为波长,波长范围350~680 nm。E作为生物发光辐射传输模型的光源输入,通过辐射传输方程[19],最终得到光谱Lw-bio。其中,2.1节中所使用的Lw-bio为Lw-bio光谱积分所得。
E=hc×MBP×P/λ
(2)
2 结果与讨论
2.1 黄海水体生物发光所致离水辐亮度幅值季节变化及空间变化特征
通过Lw-bio,结合实测MBP分析黄海水体Lw-bio幅值的分布特征。图2—图4展示了黄海海域不同季节不同光源深度下Lw-bio空间分布(不同海域水深不同,为确保不同深度下均有数据,选取4,8和15 m深度为发光光源)。
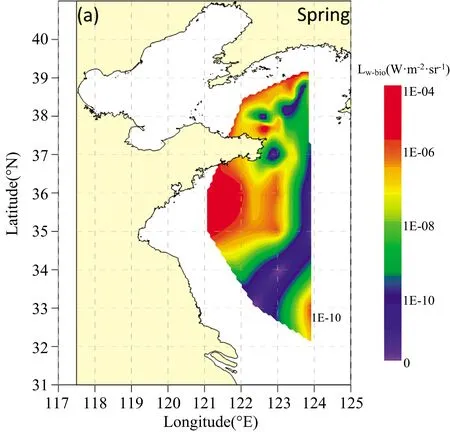
图2 春季黄海海域光源深度为4 m(a),8 m(b),15 m(c)时Lw-bio空间分布
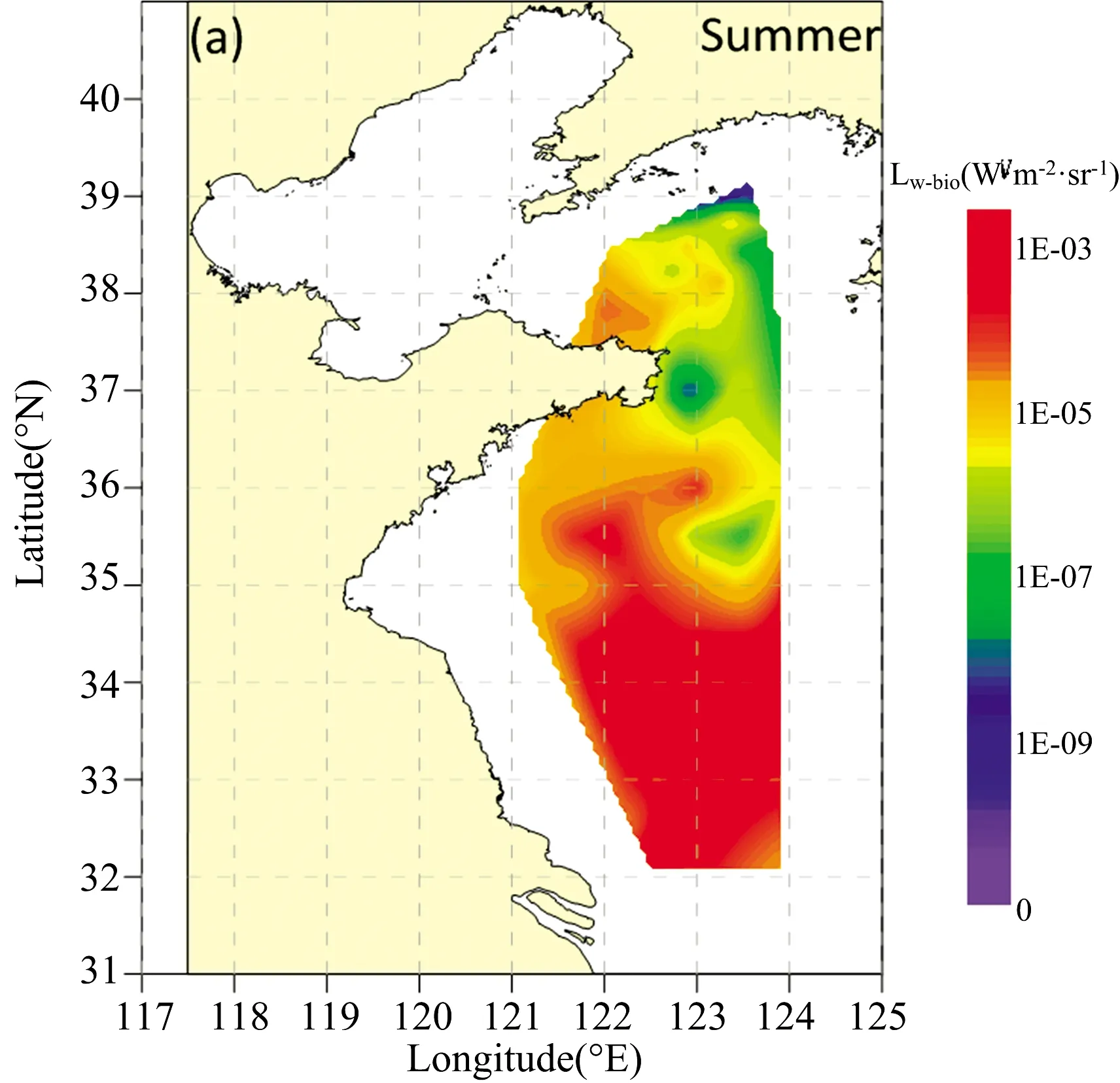
图3 夏季黄海海域光源深度为4 m(a),8 m(b),15 m(c)时Lw-bio空间分布

图4 冬季黄海海域光源深度为4 m(a),8 m(b),15 m(c)时Lw-bio空间分布
由图2,春季黄海海域Lw-bio高值区位于山东半岛威海附近海域、山东半岛以南海域以及北黄海东北部海域,上述海域衰减系数较低,发光浮游生物丰度较高,MBP在整个黄海海域处于较高水平[18]。北黄海中部海域、山东半岛东南部近岸海域以及南黄海东部海域的Lw-bio处于较低的水平。随着光源深度的增加,春季黄海海域的Lw-bio幅值仅在数值上呈现减小趋势,分布特征并未出现明显的变化。整体上,春季Lw-bio幅值呈近岸高远岸低的特征。
由图3,夏季黄海海域Lw-bio低值区集中在北黄海海域与山东半岛东部海域,高值区位于南黄海海域,当光源深度为4 m时,Lw-bio的分布特征呈北低南高。随着海水深度增加,MBP含量发生改变,当光源深度加深时,Lw-bio分布特征也发生了改变:光源深度为8 m时,低值区由山东半岛东部海域延伸至山东半岛南部海域及南黄海中部海域,高值区缩小至南黄海东部海域;光源深度为15 m时,低值区缩减至南黄海东北海域。不同光源深度下,夏季Lw-bio分布呈现差异,整体上,呈现南部高北部低的特征。
在钢结构桥梁工程施工中,对于钢结构构件,可由加工工厂统一制作,各个结构的精准度和质量比较高。另外,钢结构重量比较小,结构连接形式简单,运输便捷。通过桥梁上部结构与下部结构同事实施施工,可缩短施工周期。
由图4,在冬季黄海海域,当光源深度位于为水下4 m时,低值区分别出现在北黄海中部、南黄海南部海域以及山东半岛东南部近岸海域。高值区位于北黄海东北部海域和山东半岛以南海域。随着生物发光光源深度的加深,冬季黄海海域的Lw-bio仅在数值上呈现减小趋势,分布特征并未出现明显的变化。整体上,冬季黄海海域Lw-bio呈现中部高南部低的特征。
三个季节的Lw-bio空间分布特征呈现较大差异,Lw-bio季节变化趋势十分显著。自冬季至春季,山东半岛威海附近海域Lw-bio由低值区转为高值区,南黄海东部海域的Lw-bio转变为低值区,总体特征由中部高南部低转变为近岸高远岸低(西部高东部低)。自春季至夏季,山东半岛东南部近岸海域Lw-bio始终保持较低水平,北黄海东北部海域、山东半岛以南海域和山东半岛威海附近海域Lw-bio由高值区变为低值区,南黄海东部海域Lw-bio由低值区变为高值区,分布特征由近岸高远岸低(西部高东部低)转变为南部高北部低。
光源深度为4 m时(表2),春季Lw-bio范围在4.59×10-10~2.60×10-4W·m-2·sr-1之间,平均值为2.84×10-5W·m-2·sr-1,夏季Lw-bio范围在7.86×10-9~4.71×10-3W·m-2·sr-1之间,平均值为2.69×10-4W·m-2·sr-1,冬季Lw-bio范围在1.81×10-9~1.39×10-4W·m-2·sr-1之间,平均值为1.50×10-5W·m-2·sr-1。随着光源深度加深,Lw-bio整体呈减小趋势。当光源深度一定时,自冬季至春季,春季至夏季,Lw-bio依次增大,具有显著季节变化特征。引起Lw-bio幅值变化的主要原因有以下两点:(1)由表3可知,发光浮游生物MBP由冬季至夏季逐渐增大;(2)固有光学性质影响着Lw-bio幅值,由实测数据及前人研究[10]可知,黄海海域吸收系数、衰减系数、及后向散射系数由冬季至夏季逐渐减小。
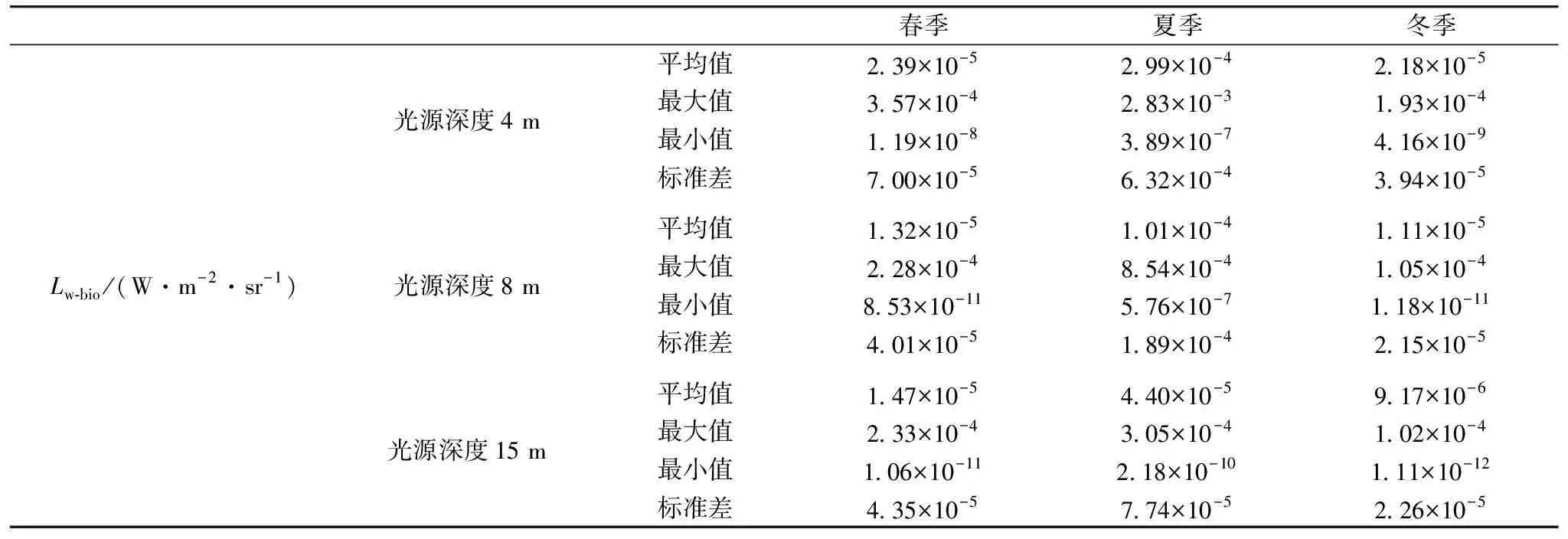
表2 不同季节不同光源深度下Lw-bio分布

表3 不同季节不同光源深度下MBP分布
2.2 黄海水体生物发光所致离水辐亮度的光谱变化
生物发光光谱呈近高斯分布特征,生物发光光源处所产生的上行生物发光辐亮度(Lu-bio)峰值波长在474 nm附近,随着光传输至海面,Lw-bio峰值波长发生改变,图5汇总了三个季节黄海海域不同发光源深度下Lw-bio的光谱分布。在不同海域、不同深度下,Lw-bio峰值波长漂移(Δλ)的范围在2~104 nm。一方面,Δλ随着光源深度加深而增大;另一方面,Δλ与固有光学性质也有紧密的联系:结合实测固有光学性质数据,在水体剖面不包含纯水的488 nm波段衰减系数(c488)低于1 m-1,吸收系数(a488)低于0.15 m-1,后向散射系数(bb488)低于0.02 m-1的海域,Δλ小于30 nm;在c488介于1~5 m-1之间,a488介于0.15~0.3 m-1之间,bb488介于0.02~0.1 m-1之间的海域,Δλ介于30~70 nm;在c488大于5 m-1,a488大于0.3 m-1,bb488大于0.1 m-1的海域,Δλ大于70 nm。较大的衰减系数可使得峰值波长由蓝光波段变为绿光波段(474 nm附近移动至578 nm附近)。而在衰减系数较小的水体中,辐亮度光谱变化较弱,若光源位于水深10 m以内,其Δλ仅在5 nm以内,即使光源位于水深30 m以下,其峰值波长最大仅漂移26 nm,仍在蓝光波段范围内。

图5 黄海海域不同发光源深度下Lw-bio光谱(不同曲线代表不同深度)
Moline等基于新泽西州近岸水体的实测生物发光数据进行辐射传输模拟,并指出随着生物发光光源深度的增加,Lw-bio峰值波长由蓝光波段移动至绿光波段,波长漂移大于90 nm。本研究模拟了固有光学性质差异较大的黄海海域生物发光辐射传输过程,结果表明,Δλ不仅与光源几何深度有关,与固有光学性质也存在紧密的联系:在固有光学性质其值较大的水体,随着光源深度的增加,其Δλ大于90 nm,而清洁水体中发光生物所产生的Δλ小于30 nm。
2.3 黄海水体生物发光光源的几何深度及光学深度与Lw-bio的联系
为了更好地将本研究应用于其他领域,如生物发光的光源反演,本研究分析了光源的光学深度τ和几何深度d分别与Δλ及峰值Lw-bio的关系。图6与图7分别展示了τ,d,与Δλ及峰值Lw-bio之间的联系:(1)从平均值上看,Δλ随着τ的增大而增大,与d无显著的递增或递减关系;从整体上看,在不同海域,由于固有光学性质的差异,即使光源的几何深度或光学深度相同,Δλ也呈现较大波动。(2)峰值Lw-bio与τ以及d存在一定的关系:峰值Lw-bio随着τ与d的加深而减小。基于此,使用τ与d分别与Δλ及峰值Lw-bio进行二元线性拟合,结果如图8和图9所示。d与Δλ及Log10(Lw-bio)二元拟合的决定系数R2=0.64,均方根误差RMSE=9.39,拟合关系为d=17.42-16.91Log10(Lw-bio)-7.458Δλ。τ与Δλ及Log10(Lw-bio)二元拟合的决定系数R2=0.90,均方根误差RMSE=4.99,拟合关系为τ=14-14.47Log10(Lw-bio)+1.18Δλ,τ包含了水体的固有光学性质信息及峰值波长信息,因此τ的拟合结果优于d。
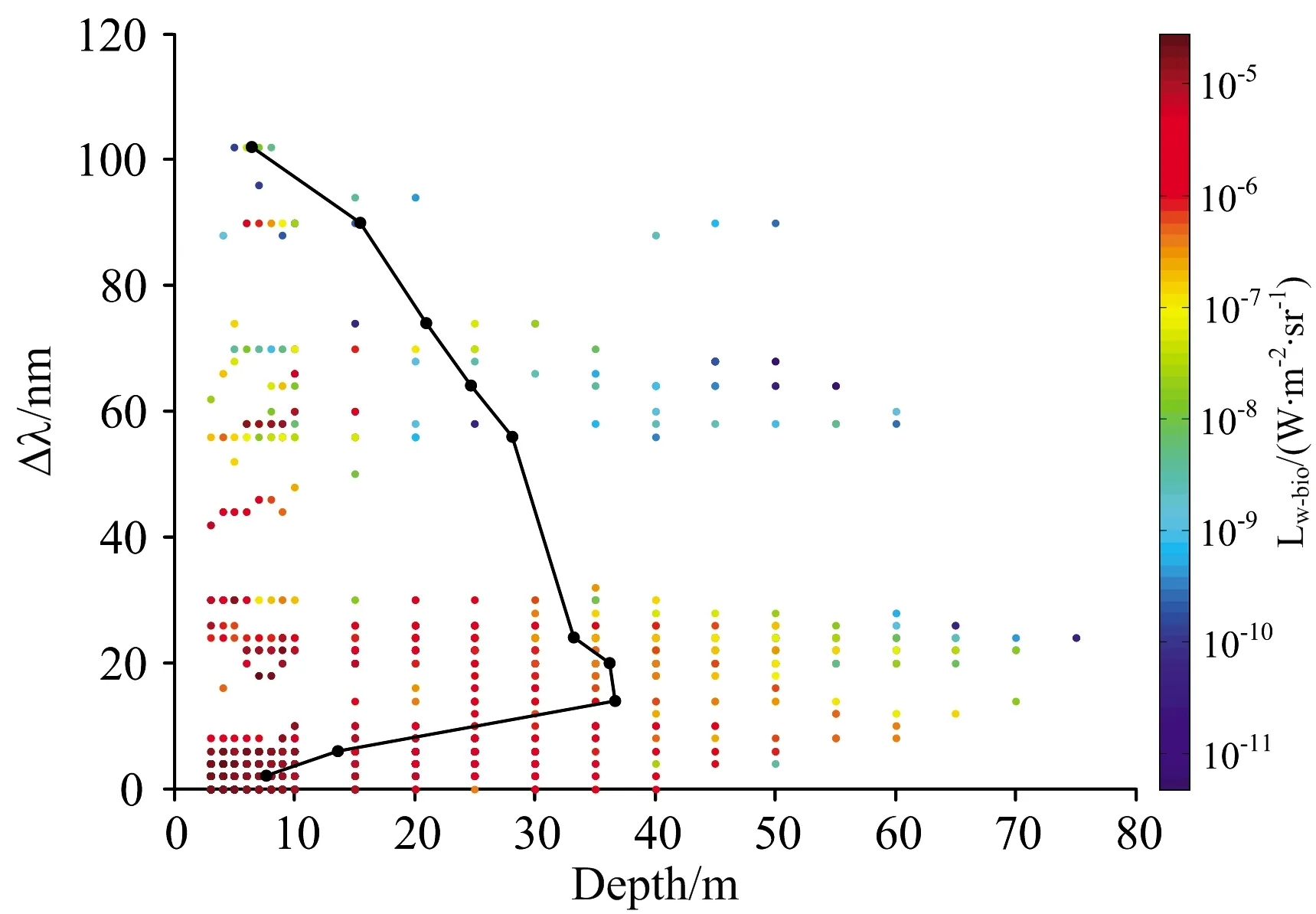
图6 几何深度d、Δλ以及峰值Lw-bio的变化
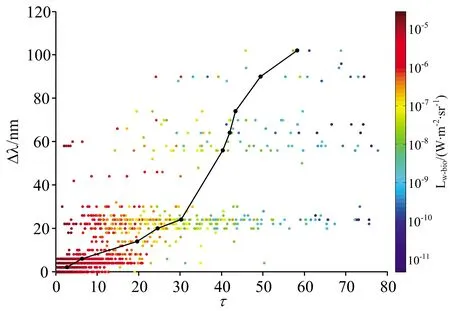
图7 光学深度τ、Δλ以及峰值Lw-bio的变化
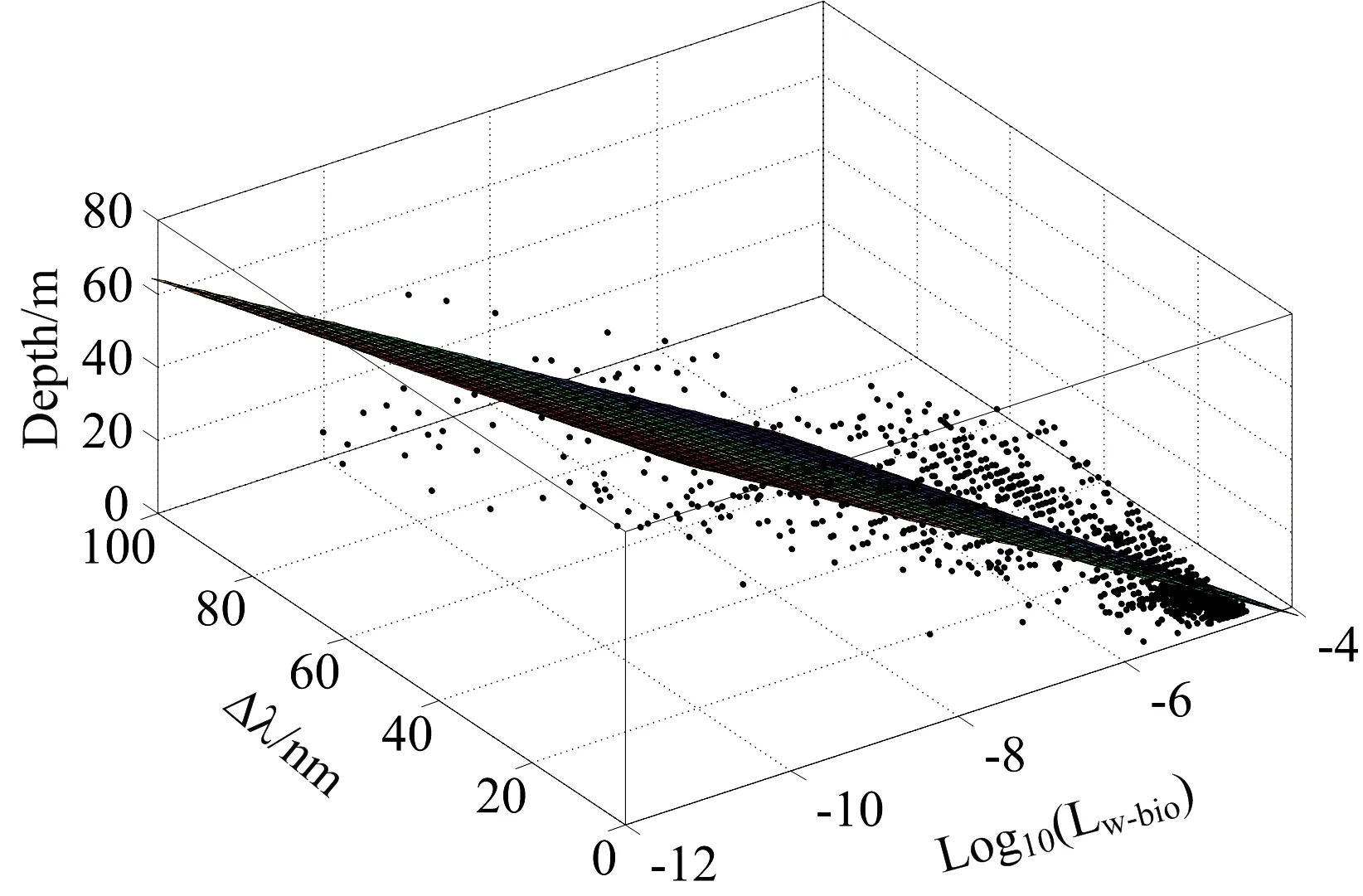
图8 几何深度d与Δλ及峰值Lw-bio拟合结果

图9 光学深度τ与Δλ及峰值Lw-bio拟合结果
基于上述结论,本研究考虑将光谱Lw-bio作为输入,用于增加水体光学信息及光谱信息,以此讨论生物发光光源的几何深度反演可行性。本节构建了简单的神经网络模型,将模拟所得光谱Lw-bio(波长数44)作为输入,输出为光源的几何深度。结果如图10与图11所示,回归系数R=1.00,均方误差MSE=0.14,最大误差为1.84 m(由图11可知),且超过97%的数据,其误差小于1 m,反演精度显著提升。
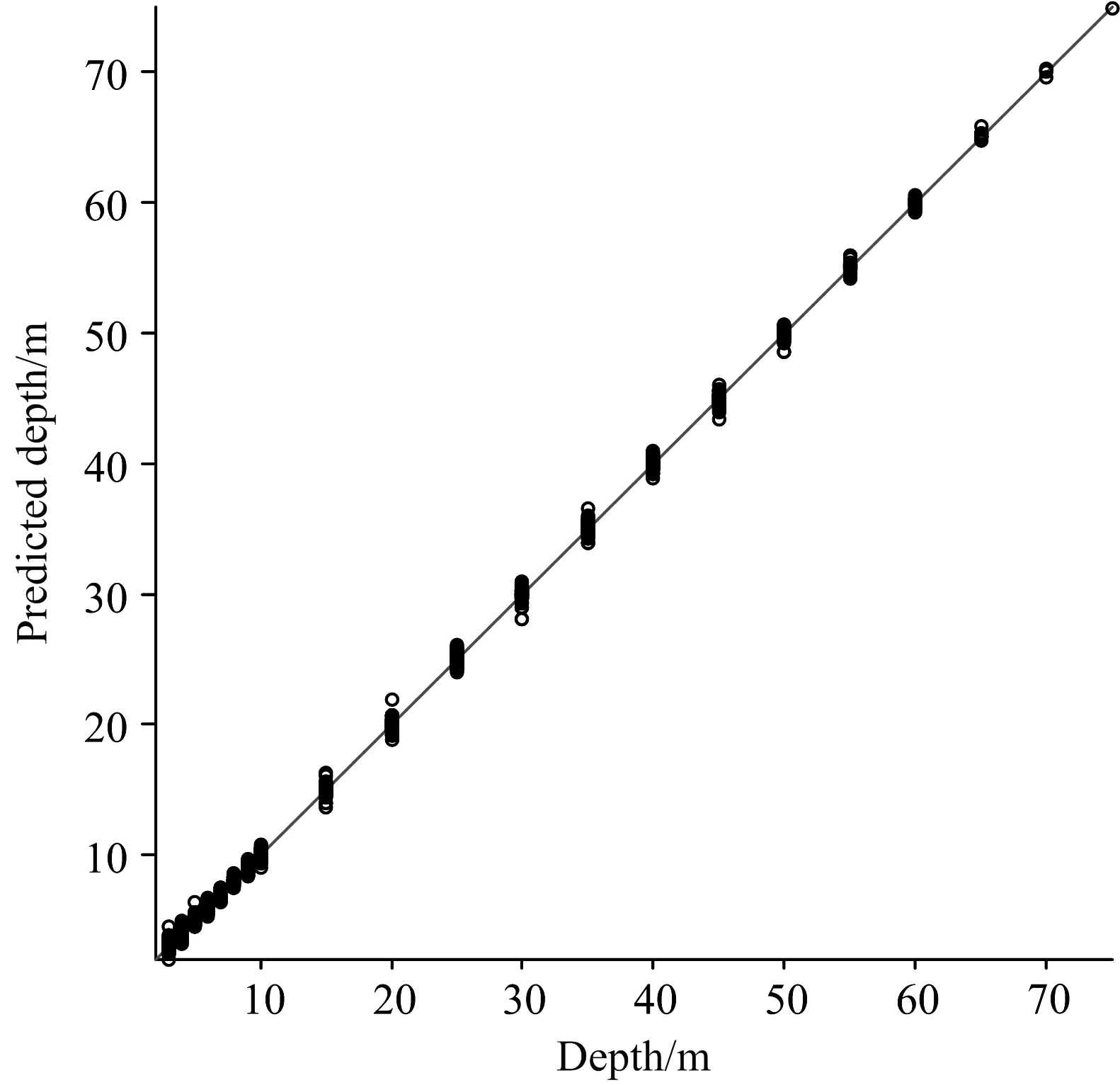
图10 生物发光光源的几何深度反演结果(直线为1∶1线)
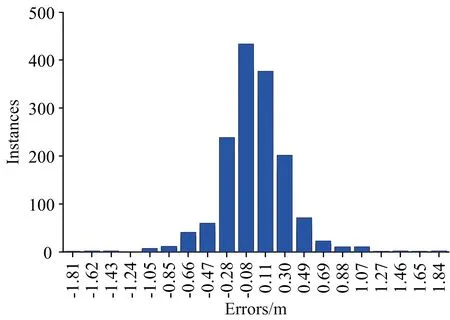
图11 几何深度反演结果误差统计(误差=实际值-预测值)
本节仅对光谱Lw-bio与几何深度的关系进行了初步的神经网络拟合,结果表明,生物发光光源的几何深度可以通过神经网络模型,由光谱Lw-bio反演所得。在未来,我们将围绕反演深度所需的最少波长数,展开进一步的研究。
3 结 论
利用在不同季节获取的黄海水体生物发光强度数据,结合同步测量的固有光学性质数据,基于辐射传输模拟,分析了生物发光所致离水辐亮度(Lw-bio)的数值变化和光谱变化特征,并讨论其与固有光学性质及生物发光所处深度的联系。
主要结论如下:
(1)随着光源深度加深,Lw-bio整体呈减小趋势。当光源深度一定时,自冬季至春季,春季至夏季,Lw-bio依次增大,具有显著季节变化特征。在空间变化方面,春季黄海海域Lw-bio幅值分布呈近岸高远岸低的特征,夏季分布呈现南部高北部低的特征,冬季分布呈现中部高南部低的特征。
(2)在光谱变化方面,Δλ随着光源深度加深而增大,并与固有光学性质有紧密的联系:在c488低于1m-1,a488低于0.15 m-1,bb488低于0.02 m-1的海域,Δλ小于30 nm;在c488介于1~5 m-1之间,a488介于0.15~0.3 m-1之间,bb488介于0.02~0.1 m-1之间的海域,Δλ介于30~70 nm;在c488大于5 m-1,a488大于0.3 m-1,bb488大于0.1 m-1的海域,Δλ大于70 nm。较大的衰减系数可使得峰值波长由蓝光波段变为绿光波段(474 nm附近移动至578 nm附近)。而在衰减系数较小的水体中,辐亮度光谱变化较弱,若光源位于水深10 m以内,其Δλ仅在5 nm以内,即使光源位于水深30 m以下,其峰值波长最大仅漂移26 nm,仍在蓝光波段范围内。
(3)黄海海域宽泛的固有光学性质对生物发光光源的几何深度反演影响较大,但可通过Lw-bio的光谱信息,反演光源的几何深度。
致谢:感谢“东方红2号”科考船上工作人员的支持,感谢出海期间所有采集数据的人,感谢课题组薛程、史鑫皓、李艺师、甘思铭四位同学的帮助。
猜你喜欢
阅读(科学探秘)(2021年8期)2021-09-01
上海工艺美术(2021年4期)2021-04-24
华人时刊(2020年21期)2021-01-14
高中生之友(中旬刊)(2019年9期)2019-10-23
电子制作(2018年2期)2018-04-18
上海农业学报(2017年3期)2017-04-10
中南大学学报(自然科学版)(2016年2期)2017-01-19
中国照明(2016年4期)2016-05-17
中国当代医药(2015年26期)2015-03-01
资源节约与环保(2015年9期)2015-01-27
