
利用分子探针法研究稻田蜘蛛集团对褐飞虱的捕食作用
2022-06-03 04:34:46王雪芹王光华蒋月丽武予清祝增荣程家安
环境昆虫学报 2022年2期
王雪芹,王光华,蒋月丽,武予清*,祝增荣,程家安*
(1. 河南省农业科学院植物保护研究所,农业部华北南部有害生物治理重点实验室,河南省农作物病虫害防治重点实验室,郑州 450002;2. 浙江大学昆虫科学研究所,水稻生物学国家重点实验室,农业部作物病虫分子生物学重点开放实验室,杭州 310058)
褐飞虱Nilaparvatalugens(Stål)属半翅目Hemiptera飞虱科Delphacidae,是亚洲稻区主要害虫,以吸取水稻植株茎杆汁液进行危害;其危害轻则可使水稻黄叶或枯死,重则能使水稻植株大面积瘫痪倒伏,造成水稻严重减产甚至绝收(程家安, 1996; 程遐年等, 2003),给我国农业生产带来严重经济损失也威胁到了国家粮食安全(林拥军等, 2011)。另外,褐飞虱还能传播水稻齿叶矮缩病(Rice ragged stunt disease)和草状矮化病(Rice grassy stunt disease)等,这些病害曾在福建省的发生面积达到了7 × 104hm2(娄永根和程家安, 2011)。因此,为有效控制褐飞虱的为害,大量研究报道了该虫的成灾原因。这些研究指出,飞虱类(包括褐飞虱)的猖獗暴发与稻田系统自然调控功能的破坏有很大关系,而稻田天敌是有效抑制该类害虫发生的重要力量,特别是捕食性天敌(Kenmoreetal., 1984; Cook and Perfect, 1989; Cheng, 2009; Heong and Hardy, 2009; Bottrell and Schoenly, 2012; Cheng, 2014)。在这些捕食性天敌中,我国稻区的种类多达820种以上,以蛛形纲Arachnida、鞘翅目Coleoptera和半翅目为主,尤其蜘蛛类天敌是捕食稻飞虱的中坚力量(吕进等, 2013; Yangetal., 2018)。
传统上天敌的捕食作用研究方法往往是建立在试验观察和统计分析为主的基础上的,如田间直接人工观察法(Carteretal., 1984; Legaspietal., 1996; Heimpeletal., 1997)、功能反应法(Holling, 1959; Harssell and Varley, 1969)和昆虫生命表分析法(Morris and Miller, 1954)等,这显然不利于天敌捕食作用的研究发展。为此,一些基于生化和现代分子生物学原理的捕食作用分析方法和技术得到了长足发展和广泛应用,如蛋白质电泳分析法(Coreyetal., 1998; Traugott, 2001)、单克隆抗体技术和DNA分子技术(Sheppard and Harwood, 2005; Schmidtetal., 2021)等。其中,DNA分子追踪食物链的荧光定量PCR(Quantitative Real-time PCR, qPCR)检测方法,因其更快速的检测过程、更高的检测灵敏度得以广泛应用(Symondson, 2002; 王光华等, 2008; Gomez-Poloetal., 2016)。qPCR根据荧光染料的类型分为嵌合染料法和荧光探针法。研究指出,qPCR能用来分析植食性害虫与捕食天敌广泛的营养关系(Lundgrenetal., 2009)。
尽管稻田系统中蜘蛛集团的物种较多,数量较大,对控制稻飞虱的发生起着重要的作用。但在研究捕食性天敌对稻飞虱的控制作用时多采用天敌数量、天敌和害虫的时间及空间生态位重叠、实验室控制条件下的单/多种天敌与单/多种害虫的捕食功能反应及作用来评价(刘向东等, 2002; 李剑泉等, 2002)。探针法qPCR可分析稻田捕食性天敌(如蜘蛛集团)对白背飞虱Sogatellafurcifera(Horvath)的捕食作用(Wangetal., 2013; Wangetal., 2017),然而,这种分子方法如何用来分析稻田蜘蛛集团对褐飞虱的捕食作用,尚需进一步的研究。因此,本研究通过在浙江省富阳地区连续两年调查稻田褐飞虱和蜘蛛种群密度和动态并采集蜘蛛样本,利用qPCR探针法检测稻田蜘蛛集团对褐飞虱的捕食作用,分析水稻不同生育期、不同年份及品种稻田蜘蛛集团对褐飞虱捕食率及蜘蛛优势种对褐飞虱的功能反应和捕食量,以期为更科学的评价稻田蜘蛛对褐飞虱的控害效能,增强保护和充分发挥自然天敌在褐飞虱绿色防控中的作用提供理论参考依据。
1 材料与方法
1.1 试验区概况及水稻种植
试验区位于浙江省富阳市(119°34′E,30°18′N),属于长江中下游亚热带湿润气候,主要种植常规稻和杂交稻单季晚稻;水稻一般6月中下旬移栽,10月中下旬收获;褐飞虱等飞虱类害虫是该稻区的最主要害虫(林拥军等, 2011)。
试验田设置在富阳市中国水稻所试验田中,种植的水稻品种为抗褐飞虱常规稻IR64和感褐飞虱杂交稻SY63(寒川一成等, 2003)。每个品种小区为完全随机区组种植,重复4次。每个小区面积为20 m2×22 m2。在整个试验期内,试验田进行相同的常规农事操作和管理,且未使用任何农药。
1.2 稻田节肢动物调查和蜘蛛集团捕食作用分析
1.2.1节肢动物发生量调查
于2011年和2012年在水稻的整个生育期对节肢动物群落共取样7次。第1次取样在水稻移栽后1周进行,随后每2周取样一次。取样时,先用大小25 cm × 25 cm × 90 cm的塑料罩罩住1丛水稻,再用真空吸虫器把里面的节肢动物吸取干净(Arida and Heong, 1992),每个品种小区重复了3丛。样本带回实验室后,在双筒体视镜下进行鉴定。统计分析水稻不同生育期和不同年份及品种褐飞虱密度、蜘蛛密度、蜘蛛与褐飞虱密度比等参数。
1.2.2蜘蛛样品采集和捕食作用分析
为测定分析蜘蛛集团对褐飞虱的捕食作用,于2011年、2012年在稻田采集并鉴定各类蜘蛛。采用盆拍法单头采集蜘蛛,放入Eppendorf管后立即装入冰盒,保存于-80℃超低温冰箱供用。采样时间从水稻移栽后3周开始,随后每2周采样一次,共采样6次。在对蜘蛛体内褐飞虱残留检测时,先用试剂盒单头提取蜘蛛基因组DNA,然后利用褐飞虱特异性探针进行qPCR检测,最后分析蜘蛛集团对褐飞虱的捕食作用。
蜘蛛集团对褐飞虱的捕食作用分析内容主要包括3个方面。(1)比较分析水稻不同生育期蜘蛛集团(科级)对褐飞虱的捕食阳性率;(2)比较分析不同年份及品种间稻田蜘蛛集团(科级)对褐飞虱的捕食作用;(3)整体比较分析两年2品种稻田4种蜘蛛优势种对褐飞虱阳性率与褐飞虱种群密度之间的关系,采用非线性模型P=aN/(1+bN)拟合褐飞虱阳性率与褐飞虱种群密度之间的功能反应曲线,其中P是蜘蛛对褐飞虱的阳性率,N是褐飞虱的种群密度;同时比较了此4种蜘蛛对褐飞虱的捕食量及在低阳性率(<50%)和高阳性率(≥50%)两个阶段的褐飞虱DNA残留量。
1.3 蜘蛛对褐飞虱捕食作用的分子检测方法
1.3.1褐飞虱特异性引物和探针的合成
利用褐飞虱核糖体DNA内转录间隔区(ITS)基因设计合成qPCR引物及特异性探针。qPCR引物和ALLGlo探针由上海超世生物技术公司合成,qPCR引物为BPH-F:TTGATTTCGAGGTTCGTCTGT ATG和BPH-R:ACATGCACAAATCACCACAGAGT,探针为MAR-TGCAGCATTCAATCTGACTGGA-MAR。
1.3.2蜘蛛体内褐飞虱残留检测体系的构建及测定
qPCR检测反应体系为体积20 μL反应体系。该体系各组成为:1 μL蜘蛛基因组DNA、2 μL 10×PCR buffer(100 mM Tris-HCl,pH7.4;500 mM KCl)、3.5 mM MgCl2、400 μM dNTP、2U r-Taq、80 nM BPH-F、80 nM BPH-R、200 nM BPH ALLGlo探针。优化后的该体系使用Bio-Rad CFX96(Bio-Rad,USA)qPCR检测系统进行检测。该体系的PCR反应程序为:95℃变性3 min,94℃变性30 s,60℃退火40 s;循环40次;每次延伸时采集荧光(Wangetal., 2017)。
利用该体系对提取的稻田蜘蛛样品总DNA进行检测,以ddH2O和饥饿1周的蜘蛛基因组DNA作为2种模板的阴性对照,同时以褐飞虱基因组DNA为阳性对照。在样品检测时每个样品设置2个重复,如果出现2个重复的扩增曲线和Ct值之差大于或等于1,则重新检测。捕食作用分析内容同上述1.2.2。
1.4 数据处理
根据试验的设计方法和统计分析原理,数据如符合正态分布且方差齐性则利用SPSS中的单因素方差分析进行多重比较(LSD法);否则利用非参数Mann-Whitney U和Kruskal-Wallis单因素ANOVA检验法进行统计分析;其中蜘蛛捕食率在年份间和品种间的分析采用SPSS的卡方适合性检验分析;采用F检验蜘蛛阳性率和褐飞虱种群密度间的功能反应曲线拟合程度,使用SigmaPlot软件作图(Wangetal., 2017)。
2 结果与分析
2.1 不同水稻生育期蜘蛛集团和褐飞虱的发生量及捕食作用
2.1.1蜘蛛集团和褐飞虱的发生量
两年两水稻品种稻田共采集到蜘蛛3 807头,分属9科27种。园蛛科Araneidae和肖蛸科Tetragnathidae种类数最多,均为6个种,分别为卵形园蛛Araneusinustus、黄斑园蛛A.ejusmodi、霍氏新园蛛Neosconadoenitzi、嗜水新园蛛N.nautica、四点高亮腹蛛Hypsosingapygmaea、四突艾蛛Cyclosasedeculata和栉齿锯螯蛛Dyschiriognathadentata、四斑锯螯蛛D.quadrimaculata、锥腹肖蛸Tetragnathamaxillosa、华丽肖蛸T.nitens、前齿肖蛸T.praedonia、圆尾肖蛸T.vermiformis;跳蛛科Salticidae为4个种,即贺氏菱头蛛Bianorhotingchiechi、长腹蝇狮Marpissaelongata、纵条蝇狮M.magister和黑色蝇虎Plexippuspaykulli;狼蛛科Lycosidae和皿蛛科Linyphiidae各为3个种,分别是沟渠豹蛛Pardosalaura、拟环纹豹蛛P.pseudoannulata、拟水狼蛛Piratasubpiraticus、草间小黑蛛Erigonidiumgraminicolum、驼背额角蛛Gnathonariumgibberum和食虫瘤胸蛛Ummeliatainsecticeps;蟹蛛科Thomisidae为鞍形花蟹蛛Xysticusephippiatus和三突花蛛Misumenopstricuspidatus2个种;球腹蛛科Theridiidae、管巢蛛科Clubionidae和盗蛛科Pisauridae均为1个种,分别为八斑球腹蛛Coleosomaoctomaculatum、日本管巢蛛Clubionajaponicola和白跗狡蛛Dolomedespallitarsis。其中,锥腹肖蛸、拟环纹豹蛛、食虫瘤胸蛛、八斑球腹蛛为稻田蜘蛛优势种。
褐飞虱密度随调查时间呈规律性增加,且在调查的最后时间点DAT(Days after transplanting,DAT)91达最大值76.29±6.21头/丛,显著高于DAT7、DAT21、DAT35和DAT49的值(P<0.05);蜘蛛密度及蜘蛛与褐飞虱的密度比值随调查时间呈先增加后下降的规律性变化,其中,蜘蛛密度在水稻移栽后DAT63达到最大值6.89±0.54头/丛,该密度显著高于水稻其它生育期的蜘蛛密度(P<0.05);蜘蛛与褐飞虱的密度比值在水稻移栽后DAT35达到最大值39.74±5.80,显著高于DAT7、DAT63、DAT77和DAT91的值(P<0.05)(图1)。
2.1.2蜘蛛集团对褐飞虱的捕食阳性率
稻田采集和分析的蜘蛛样品除其它科(包括蟹蛛科、管巢蛛科和盗蛛科)外,剩余各科蜘蛛对褐飞虱的捕食阳性率在DAT21和DAT91之间都存在显著性差异(P<0.05)(图2);整体上,狼蛛科、皿蛛科、球腹蛛科、肖蛸科、跳蛛科和园蛛科捕食率在DAT21、DAT35和DAT77、DAT91之间有显著性差异(P<0.05);另外,各科蜘蛛对褐飞虱的捕食阳性率均随水稻生育期的发展而逐渐增加,特别在水稻移栽后的DAT77和DAT91,狼蛛科、球腹蛛科、肖蛸科和跳蛛科蜘蛛对褐飞虱的阳性率均达到了最大值(图2)。
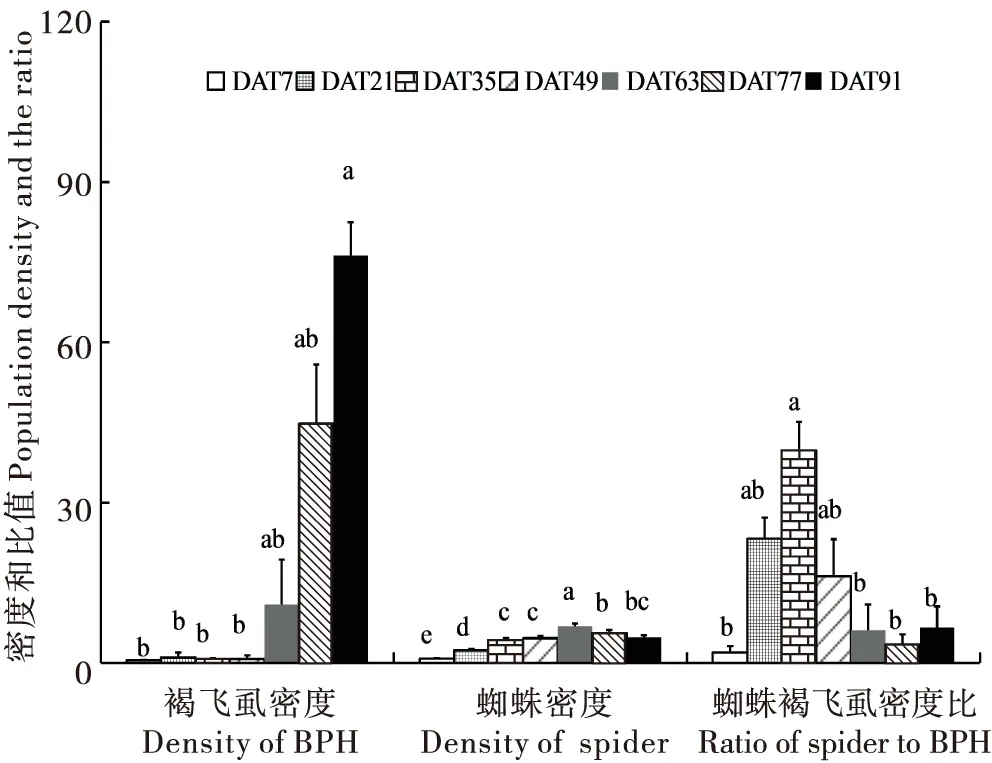
图1 水稻不同生育期稻田每丛水稻蜘蛛和褐飞虱密度及密度比值的比较Fig.1 Comparison of density of BPH, spider, the ratio of spiders to BPH among crop stages注:图上小写字母分别代表不同处理间的差异显著性(P<0.05)。下同。Note: Different lowercase letters in the columns indicated significant differences. Asterisk meant significant difference between years and fields planted with resistant and susceptible varieties (P<0.05).The same below.
2.2 不同年份及品种稻田蜘蛛集团褐飞虱的发生量及捕食作用
2.2.1蜘蛛集团和褐飞虱田间发生量
2012年褐飞虱密度、蜘蛛密度均显著高于2011年(P<0.05)(图3);另外,2011年褐飞虱密度很低(0.46±0.23),使得2011年蜘蛛密度与褐飞虱密度的比值(46.39±20.76)显著高于2012年的比值(2.65±1.36)(P<0.05)(图3)。
两年间感性品种SY63稻田的褐飞虱密度、蜘蛛密度均显著高于抗性品种IR64的两者密度(P<0.05),抗性品系稻田蜘蛛与褐飞虱密度比值(28.46±14.29)显著高于感性品种比值(2.81±1.70)(P<0.05)(图3)。
2.2.2蜘蛛集团对褐飞虱的田间捕食阳性率
各科蜘蛛对褐飞虱捕食阳性率年份间均有显著性差异(P<0.05),且2012年褐飞虱捕食阳性了显著高于2011年;感性品种与抗性品种的褐飞虱捕食阳性率只有狼蛛科和球腹蛛科之间存在显著性差异,狼蛛科在感性品种稻田的捕食阳性率显著高于抗性品种,而球腹蛛科在感性品种稻田的捕食阳性率显著低于抗性品种(P<0.05)(表1)。
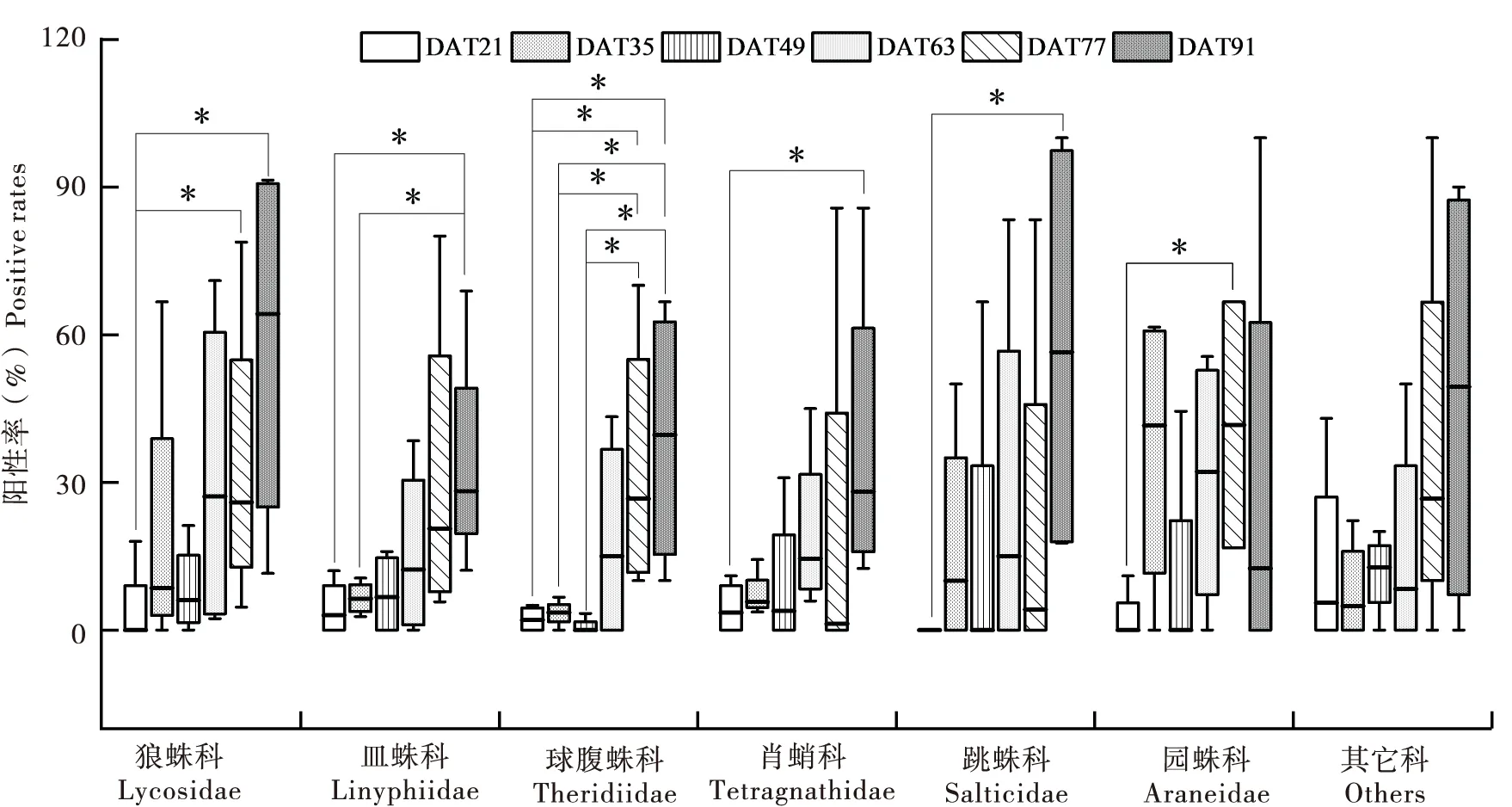
图2 水稻不同时期同类蜘蛛捕食褐飞虱阳性率的动态比较Fig.2 Comparison of the positive rates to BPH for each taxon of spiders among crop stages注:图上*代表水稻移栽后不同天数的差异显著性(P<0.05),箱体中间线为阳性率的中位数,箱体的上下线分别是上四分位数和下四分位数。下同。Note: Asterisk meant significant difference among different DAT (P<0.05). The same below.
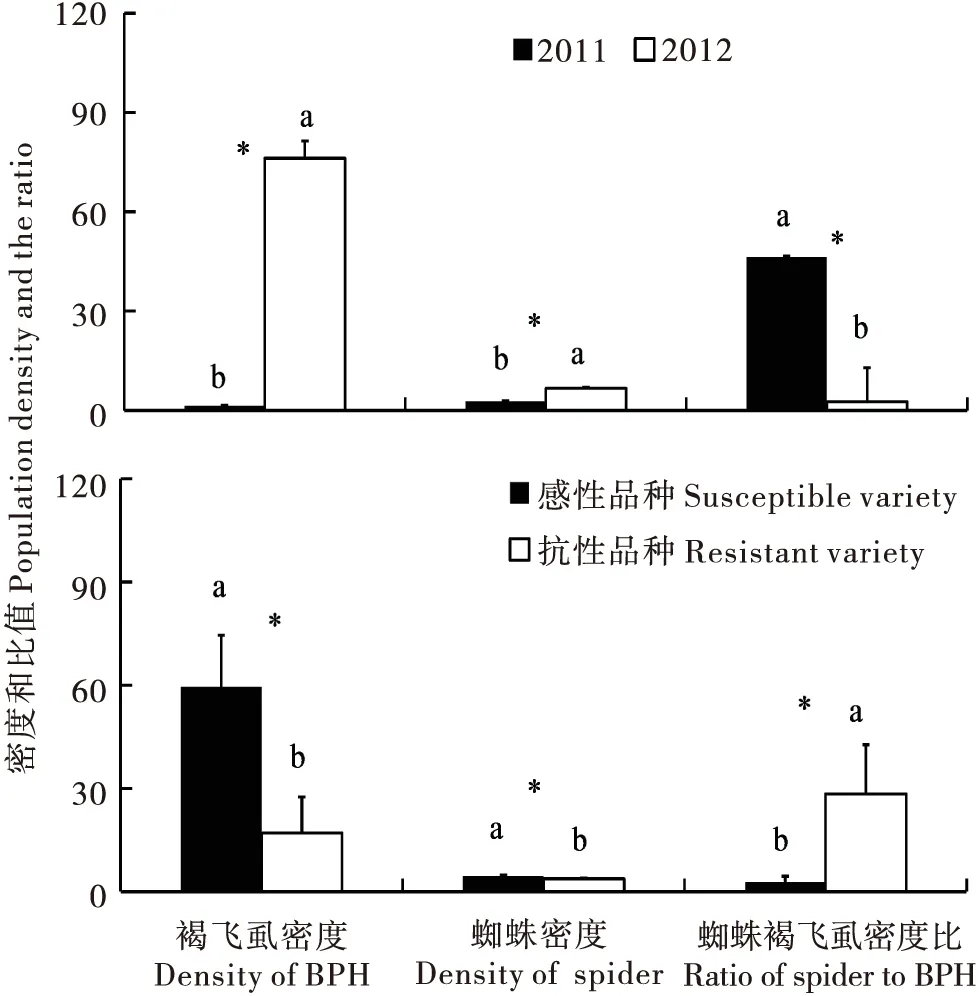
图3 褐飞虱和蜘蛛密度及密度比值在不同年份、不同抗性水稻品种间的比较Fig.3 Comparison of the population density and the ratio between years and varieties
2.3 稻田4种蜘蛛优势种对褐飞虱的功能反应及捕食量
2.3.1稻田4种蜘蛛优势种对褐飞虱的功能反应
4种蜘蛛优势种捕食阳性率均随着褐飞虱种群密度的增加而增加;除锥腹肖蛸外,其他蜘蛛捕食阳性率与褐飞虱密度间均呈饱和反应曲线关系(图4)。在褐飞虱密度为10~20头/丛时,拟环纹豹蛛、八斑球腹蛛和食虫瘤胸蛛的捕食阳性率基本呈线性增长,但当褐飞虱密度达到50~100头/丛时,除锥腹肖蛸外,它们的这种捕食阳性率就会呈现相对饱和状态,而锥腹肖蛸的捕食阳性率呈线性增长趋势(图4);拟环纹豹蛛和八斑球腹蛛的a值分别是9.59±5.59和3.02±1.21,显著大于锥腹肖蛸的0.53±0.24和食虫瘤胸蛛的1.38±0.48(P<0.05)。拟环纹豹蛛的b值是0.12±0.08,其均大于八斑球腹蛛(0.04±0.02)、食虫瘤胸蛛(0.04±0.02)和锥腹肖蛸(0.01±0.00)的值。
2.3.2稻田4种蜘蛛优势种对褐飞虱的捕食量
蜘蛛消化道内褐飞虱DNA残留量(Ct值)qPCR检测结果表明,4种蜘蛛的Ct值存在显著性差异(P<0.05),食虫瘤胸蛛的Ct值(28.94±0.32)显著高于八斑球腹蛛(26.93±0.33)和拟环纹豹蛛(25.89±0.51)的Ct值(P<0.05);锥腹肖蛸和八斑球腹蛛的Ct值(28.11±0.44)均显著高于拟环纹豹蛛的Ct值(P<0.05)(图5)。同时,比较了这4种蜘蛛整体在低阳性率(<50%)和高阳性率(≥50%)两个阶段的褐飞虱DNA残留量,结果显示低和高阳性率两个阶段的Ct值分别是28.97±0.39和26.95±0.23,存在显著性差异(P<0.05),这表明在高阳性率阶段这4种蜘蛛体内有更多的褐飞虱猎物DNA残留。

表1 两年两品种稻田蜘蛛集团发生量及其对褐飞虱的捕食阳性率比较
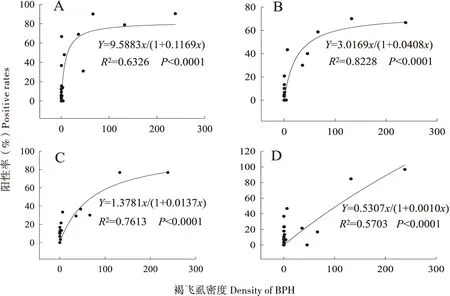
图4 田间4种蜘蛛对褐飞虱阳性率随褐飞虱种群密度增加的非线性饱和反应模型Fig.4 Non-linear relationships between prey (BPH) densities and positive rates for 4 spider species in field condition注:A,拟环纹豹蛛;B,八斑球腹蛛;C,食虫瘤胸蛛;D,锥腹肖蛸。Note: A, Pardosa pseudoannulata; B, Coleosoma octomaculata; C, Ummeliata insecticeps; D, Tetragnatha maxillosa.
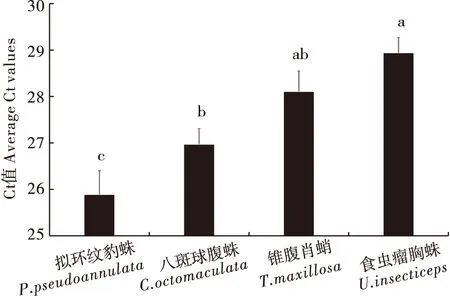
图 5 4种主要蜘蛛消化道内褐飞虱残留量(Ct值)的比较Fig.5 Comparison of the quantitative evaluations for predation on BPH among 4 spider species
3 结论与讨论
本文以亚热带浙江省富阳市稻田生态系统为研究对象,调查了两年不同品种稻田褐飞虱和蜘蛛的发生密度后,利用荧光定量PCR分子探针技术分析了蜘蛛对褐飞虱的捕食作用。结果表明,稻田生态系统包含9科27种蜘蛛,在水稻不同生育期,蜘蛛密度与褐飞虱发生量间具有跟随效应;两年不同品种稻田蜘蛛和褐飞虱密度及密度比值在年份间和品种间存在显著性差异;同时,捕食作用的分子检测结果指出,蜘蛛对褐飞虱的捕食率在年份间存在显著性差异,狼蛛科和球腹蛛科蜘蛛捕食率在品种间也存在显著性差异。稻田4种蜘蛛优势种对褐飞虱的捕食阳性率与褐飞虱的种群密度均呈饱和反应曲线或线性反应模型,拟环纹豹蛛捕食褐飞虱的DNA残留量显著高于八斑球腹蛛、锥腹肖蛸和食虫瘤胸蛛。
3.1 水稻不同生育期蜘蛛集团和褐飞虱的发生量及捕食作用
稻田生态系统是水陆复合的具有丰富生物多样性和复杂稳定生态系统服务功能的开放的农业生态系统,包含了数千种的节肢动物种类,这些种类根据功能可以分为植食者、腐食者、寄生者和捕食者等(程家安, 1996; Settleetal., 1996)。蜘蛛是稻田捕食者种类和数量组成中最重要的部分,连续两年的调查和采集,仅仅在浙江省富阳市中国水稻所试验田就采集到9科27种蜘蛛;相关研究也指出,在全国稻田所采集到的蜘蛛标本中,已鉴定373种,隶属23科109属,其中东南稻区(包括浙江)因气候适宜、地形和景观复杂多样化,蜘蛛种类以228种居全国之首(吕仲贤, 2012)。
稻田蜘蛛除其它科外,剩余各科蜘蛛对褐飞虱的捕食阳性率在DAT21和DAT91之间都存在显著性差异(P<0.05)(图2);整体上,狼蛛科、皿蛛科、球腹蛛科、肖蛸科、跳蛛科和园蛛科捕食率主要在DAT21、DAT35和DAT77,DAT91之间有显著性差异(P<0.05);另外,各科蜘蛛对褐飞虱的捕食阳性率均随水稻生育期的发展而逐渐增加,特别在DAT77和DAT91,狼蛛科、球腹蛛科、肖蛸科和跳蛛科蜘蛛对褐飞虱的阳性率均达到了最大值(图2)。该结果表明,首先,蜘蛛对特定猎物褐飞虱的捕食阳性率与其密度密切相关,稻田主要害虫褐飞虱在田间群落中占有较大优势,因此即便是广食性捕食者蜘蛛对目标猎物仍然存在着紧密跟随现象;其次,水稻分蘖期、拔节期是褐飞虱迁入和定殖期,也是自然天敌调控稻飞虱的关键时期,对稻飞虱的种群发展起着至关重要的作用(Sigsgaard, 2007; Qianetal., 2021)。
3.2 不同年份及品种稻田蜘蛛集团和褐飞虱的发生量及捕食作用
2012年褐飞虱、蜘蛛密度均显著高于2011年(P<0.05)(图3),说明该年是褐飞虱暴发的年份,虫量远远高于2011年;根据天敌的跟随效应,2012年田间天敌蜘蛛密度也显著高于2011年(P<0.05)。两年间感性品种SY63稻田的褐飞虱密度、蜘蛛密度均显著高于抗性品种IR64的两者密度(P<0.05)(图3),该结果表明,对于感性品种稻田,褐飞虱具有更高的繁殖力和更大的数量,稻田蜘蛛对褐飞虱也有一定的跟随性。相关的研究表明,在20世纪80年代末,我国的褐飞虱从生物型1变为生物型2,而在此之前,褐飞虱以生物型1为主,不为害带有抗虫基因Bph1的杂交水稻SY63,后来汕优系列对褐飞虱的抗性逐步丧失,1997年SY63开始丧失对褐飞虱的抗性,造成了褐飞虱在长江中下游稻区的暴发(寒川一成等, 2003)。同时调查发现,由于2011年褐飞虱密度很低,使得2011年蜘蛛密度与褐飞虱密度的比值(46.39±20.76)显著高于2012年的比值(P<0.05),而且抗性品系稻田蜘蛛与褐飞虱密度比值显著高于感性品种比值(P<0.05)(图3),这说明抗性品种和自然天敌对于繁殖快、周期短、易暴发的褐飞虱等稻飞虱来说依然是最重要的防控策略(寒川一成等, 2003),而这也是热带稻区比亚热带稻区稻飞虱暴发少、为害轻的最主要原因(Huetal., 2014)。
两年间所测的各科蜘蛛褐飞虱捕食阳性率均为2012年的捕食率显著高于2011年的捕食率(P<0.05),感性品种与抗性品种的褐飞虱阳性率只有狼蛛科和球腹蛛科之间存在显著性差异(P<0.05)(表1);该结果表明,在褐飞虱暴发的2012年,尽管各科蜘蛛褐飞虱捕食率都显著高于非暴发的2011年,有比较高的捕食率,但仍不足以控制褐飞虱种群的发展,这与褐飞虱迁入动态、增长倍数高等环境因素有关(程家安等, 2008)。
3.3 稻田4种蜘蛛优势种对褐飞虱的功能反应及捕食量
本研究中4种蜘蛛优势种对褐飞虱的捕食阳性率与褐飞虱的种群密度均呈饱和反应曲线或线性反应模型,说明这4种优势蜘蛛虽为广食性天敌,但均能随着褐飞虱种群密度的增长而主要捕食褐飞虱。其中拟环纹豹蛛具有最高的捕食功能,在褐飞虱种群发展的早期阶段,该蛛能更快地转向捕食褐飞虱,并能在较低褐飞虱种群密度下达到较高的捕食阳性率,其次为八斑球腹蛛;这与Wang等(2017)用同样方法对稻田这4种蜘蛛优势种对白背飞虱捕食功能的研究结果相似。相关研究也指出,稻田狼蛛和肖蛸对盲蝽科害虫Stenotusrubrovittatus捕食率同样随猎物密度增加而增加(Kobayashietal., 2011),研究者是在该虫种群密度高峰期在稻田选择20个采样点,调查S.rubrovittatus密度与检测狼蛛和肖蛸对其捕食阳性率;相比较而言,本文通过水稻全生育期的6次褐飞虱密度调查和天敌蜘蛛系统采样,能更准确的反映褐飞虱的迁入、定殖、繁殖和增长等田间种群动态和4种蜘蛛优势种捕食率对其种群密度变化的响应。
通过研究4种蜘蛛优势种消化道内褐飞虱DNA残留量(Ct值)发现,食虫瘤胸蛛的Ct值显著高于八斑球腹蛛和拟环纹豹蛛;锥腹肖蛸和八斑球腹蛛的Ct值均显著高于拟环纹豹蛛的Ct值(图5)。这表明拟环纹豹蛛消化道猎物DNA残留量显著高于八斑球腹蛛、锥腹肖蛸和食虫瘤胸蛛消化道的残留量,而八斑球腹蛛则显著高于食虫瘤胸蛛(图5)。就整个水稻生育期来讲,拟环纹豹蛛消化道内有最高的褐飞虱残留量,这与拟环纹豹蛛属于游猎型蜘蛛、食性贪婪、食量较大有关,而相关的研究指出,拟环纹豹蛛在水稻生态系统对稻飞虱起着最重要的控害作用(Wangetal., 2017)。在茶园生态系统,Yang等(2011)利用探针法发现茶园游猎型蜘蛛优势种白斑猎蛛Evarchaalbaria对主要害虫假眼小绿叶蝉Empoascavitis的捕食阳性率和残留量显著高于鞍形花蟹蛛Xysticusephippiatus、漂亮美蒂蛛Meotipapulcherrima和八斑球腹蛛,在假眼小绿叶蝉种群调控中起着关键作用(Yangetal., 2011)。本研究中比较这4种蜘蛛整体在低阳性率阶段(<50%)和高阳性率阶段(≥50%)两个阶段的褐飞虱DNA残留量,结果表明这两个阶段之间残留量存在显著性差异,说明高阳性率阶段有更多的褐飞虱猎物DNA残留,即随着褐飞虱种群密度的增长,这4种蜘蛛个体对褐飞虱不仅有更高的捕食率,同时有更大的捕食量。同样在稻田生态系统,Hsu等通过同位素(δ13C和δ15N)分析发现,随着稻田植食者丰度的增加,捕食性天敌对植食者的相对捕食量从水稻分蘖期的34%~35%增长到成熟期的90%~93%(Hsuetal., 2021)。
综上所述,本文深入研究了种类丰富、数量众多的蜘蛛在不同年份和不同品种稻田对褐飞虱的自然调控作用。结果表明,稻田蜘蛛集团对褐飞虱具有较强的控害功能,而助增、前置和增强各类蜘蛛集团对褐飞虱的生态控害功能可通过种植抗性品种、稻田生态工程、减施和晚施杀虫剂等措施的有机结合来实现(祝增荣等, 2012; Zhuetal., 2018; Qianetal., 2021)。
猜你喜欢
云南农业(2024年3期)2024-03-11 10:58:46
中国水稻科学(2023年3期)2023-05-18 02:17:50
昆虫学报(2021年8期)2021-09-06 05:42:40
趣味(作文与阅读)(2021年5期)2021-08-19 02:48:54
今日农业(2020年21期)2020-12-19 13:52:28
昆虫学报(2020年8期)2020-10-15 10:50:34
作文大王·低年级(2019年2期)2019-01-23 11:35:18
创新作文(小学版)(2018年19期)2018-11-30 01:56:08
小朋友·快乐手工(2018年3期)2018-04-22 11:48:52
小学阅读指南·低年级版(2017年6期)2017-06-12 08:22:47
