
基于ISSR的豚草和三裂叶豚草遗传多样性研究
2022-05-30 03:15付开赟丁新华吐尔逊阿合买提张国良付卫东加马力丁吾拉扎汗郭文超
生物安全学报 2022年2期
王 钿, 付开赟, 丁新华, 何 江, 吐尔逊·阿合买提, 张国良, 付卫东, 文 俊, 加马力丁·吾拉扎汗, 王 兰*, 郭文超
1塔里木大学植物科学学院,新疆 阿拉尔 843300; 2新疆农业科学院植物保护研究所/农业部 西北荒漠绿洲作物有害生物综合治理重点实验室,新疆 乌鲁木齐 830091; 3中国农业科学院 农业环境与可持续发展研究所,北京 100081; 4伊犁州治蝗灭鼠指挥中心,新疆 伊犁 835000; 5新疆农业科学院微生物应用研究所,新疆 乌鲁木齐 830091
豚草AmbrosiaartemisiifoliaL.和三裂叶豚草AmbrosiatrifidaL.均是恶性杂草,是世界上公认的有害植物;分别是我国首批和第二批公布的危害严重的外来有害生物(万方浩等,2005)。豚草和三裂叶豚草繁殖力和植株竞争力强,生长中易排斥其他植物,形成优势,成片生长,改变当地原有植物种类和群落类型,导致生物多样性下降;其侵入农田,与作物竞争导致作物产量损失,且其花粉对人体具有高致敏性(Bullocketal.,2012)。豚草别称艾叶破布草,一年生草本植物,属菊科,原产北美洲(包颖,2018)。近200年在全球范围内蔓延,已扩散到80多个国家(Montagnanietal.,2017),20世纪30年代首次传入中国。目前豚草广泛分布于我国东北、华北、华中、华东等地区(柳晓燕等,2016; 邵云玲和曹伟,2017; Chenetal.,2007; Qinetal.,2014)。三裂叶豚草别称大破布草,属菊科豚草属植物,原产北美洲,20世纪30年代传入我国, 入侵我国黑龙江、北京等地(马丽娟等,2020; 马倩倩等,2020)。2010年,在新疆伊犁河谷新源县首次发现豚草和三裂叶豚草入侵,随后迅速扩散蔓延,现呈大面积暴发趋势,已严重威胁到当地农牧民的生产和生活(何江等,2020; 梁巧玲和陆平,2014)。
简单序列重复区间扩增多态性(inter-simple sequence repeat, ISSR)是由Zietkiewiczetal.(1994)基于简单序列重复(simple sequence repeats, SSR)建立的一种分子标记技术,其在SSR的3′或 5′端加上2~4个锚定核苷酸,以提高反应的专一性,PCR产物经电泳分析后可获得遗传多样性信息(Fernandez & Figueiras,2002)。ISSR技术具有操作简便、重复性好和多态性高等优点,在遗传多样性研究中应用广泛(Godwinetal.,2010)。目前ISSR已在多种动植物的亲缘关系、种质鉴定等研究方面得到应用(王明明等,2007)。本研究采用ISSR技术对8个豚草居群和7个三裂叶豚草居群的遗传多样性与遗传分化进行分析,旨在揭示新疆伊犁河谷豚草和三裂叶豚草居群的遗传多样性水平、居群间和居群内的遗传分化程度以及不同居群间的亲缘关系。
1 材料与方法
1.1 材料
三裂叶豚草居群分别采自新疆新源县阿勒玛勒镇阿克其村(43°31′80″ N,83°17′17″ E,海拔806.6 m)、新疆新源县则克台镇沙哈村(43°32′25″ N,83°17′97″ E,海拔828.2 m)、新疆新源县铁热克阿吾孜村(43°31′29″ N,83°22′50″ E,海拔817.9 m)、新疆新源县吐尔根农场牧业队(43°32′24″ N,83°23′36″ E,海拔951.3 m)、新疆新源县则克台村(43°31′23″ N,83°20′49″ E,海拔801.6 m)、新疆新源县那拉提杏花村(43°32′23″ N,83°26′15″ E,海拔1039.5 m)、辽宁沈阳市沈河区(41°49′45″ N,123°33′26″ E,海拔37.9 m)7个居群。豚草居群分别采自新疆察布查尔县绰霍尔镇博孜墩村(43°49′53″ N,81°59′25″ E,海拔553.0 m)、贵州惠水县龙洞村(26°18′12″ N,106°39′20″ E,海拔973.1 m)、广西来宾市大湾镇石山村(23°31′50″ N,109°21′57″ E,海拔69.0 m)、广西来宾市平阳镇平阳村(23°22′33″ N,109°13′26″ E,海拔89.0 m)、广西来宾市武宣县金鸡乡鱼步村(23°32′18″ N,109°21′30″ E,海拔74.0 m)、新疆新源县铁木里克村(43°19′45″ N,83°50′23″ E,海拔845.2 m)、新疆新源县玉什开普台尔村(43°17′16″ N,83°23′49″ E,海拔832.6 m)、新疆新源县阿热勒托别镇哈拉西克村(43°15′59″ N,83°27′52″ E,海拔815.0 m)8个居群。每个居群采集植物样品16株,共240个样品。采样时,每个居群间隔2 km以上为一样点;内地样品由中国农业科学院农业环境与可持续发展研究所付卫东老师采集提供,疆内样品由新疆农业科学院植物保护研究所采集提供。每个居群内采样时每株间隔在20 m以上的植株为一份样品,选取健康且长势相同的植株叶片置于保鲜袋中,带回实验室用无菌水清洗,保存于-80 ℃的超低温冰箱中备用。
1.2 方法
1.2.1 基因组DNA的提取 取0.10 g叶片放至1.5 mL的标号EP管中,再将EP管放入液氮中15 s;取出后充分研磨至粉末状,加入800 μL的DNAiso Reagent,静置2 min;加入200 μL氯仿,漩涡60 s后15000 r·min-1、4 ℃离心10 min;移液枪吸取上清液移至吸附柱中,15000 r·min-1、4 ℃离心1 min,弃下清液;加入300 μL无水乙醇,15000r·min-1、4 ℃离心1 min,弃下清液;加入700 μL 75%酒精,7500 r·min-1、4 ℃离心1 min,弃下清液;加入700 μL 75%酒精,7500 r·min-1、4 ℃离心1 min,弃下清液;空转15000 r·min-1、4 ℃离心2 min;加入30 μL ddH2O(TE buffer),静置2 min,15000 r·min-1、4 ℃离心1 min,-20 ℃保存DNA。
1.2.2 ISSR-PCR扩增 依据不列颠哥伦比亚大学(The University of British Columbia,UBC)公布的100条ISSR通用引物(https:∥wenku.baidu.com/view/62eea8f1fc4ffe473268ab76),由上海生物工程公司合成。以豚草和三裂叶豚草DNA为模板,用GT9612型96梯度PCR仪进行ISSR通用引物的筛选和体系程序优化。选取多态性高、重复性好的引物,对DNA模板的浓度、引物用量和反应温度等进行优化。根据已优化的体系和程序,用筛选得到的引物对所有豚草和三裂叶豚草样品进行ISSR-PCR扩增。将PCR产物点样置于2%(琼脂糖质量与TAE缓冲液体积比)琼脂糖凝胶(含溴化乙锭)电泳检测;检测电压75 V,35 min。在凝胶成像仪中拍照记录。
1.3 数据分析
以ISSR-PCR照片为依据,用Excel表统计谱带数量有谱带处标“1”,无谱带处标“0”,建立 0/1 矩阵;用POPGEN 32软件计算Nei′s基因多样性指数和Shannon′s信息指数2个遗传参数;用NTSYS 2.10软件对15个居群进行聚类分析,绘制亲缘关系树状聚类图;利用经纬度计算各居群间的直线距离,用SAS软件进行地理距离和遗传距离的相关性分析。
2 结果与分析
2.1 ISSR扩增结果
经优化获得最佳反应体系(25 μL):12.5 μL 2×Taq PCR Mix,2 μL引物,2 μL模板DNA,8.5 μL ddH2O。PCR程序设置:94 ℃预变性3 min;94 ℃变性30 s,55 ℃退火30 s,72 ℃延伸1 min,35个循环;72 ℃延伸5 min。取部分DNA样品(4 ℃保存),将其作为模板进行引物筛选。筛选出13条多态性高、谱带清晰且重复性好的引物(表1)。
采用优化后的反应体系与程序,用13条引物对240份样品进行 ISSR-PCR扩增实验,其中引物UBC884对三裂叶豚草扩增多态性条带比率最低,为78.57%;引物UBC815对豚草扩增多态性比率最低,为87.50%。引物UBC812、 UBC815、 UBC822、UBC823、 UBC841、UBC848、UBC866对三裂叶豚草扩增多态性条带比率达到100%;引物UBC823、UBC825、 UBC848、 UBC866、 UBC884、UBC887对豚草扩增多态性条带比率达到100%。由于三裂叶豚草在新疆伊犁哈萨克自治州新源县那拉提杏花村发生危害严重,因此,选取引物UBC841对三裂叶豚草居群的ISSR进行扩增,扩增图谱如图1所示。

表1 筛选出的引物基本信息Table 1 Basic information of screened primers
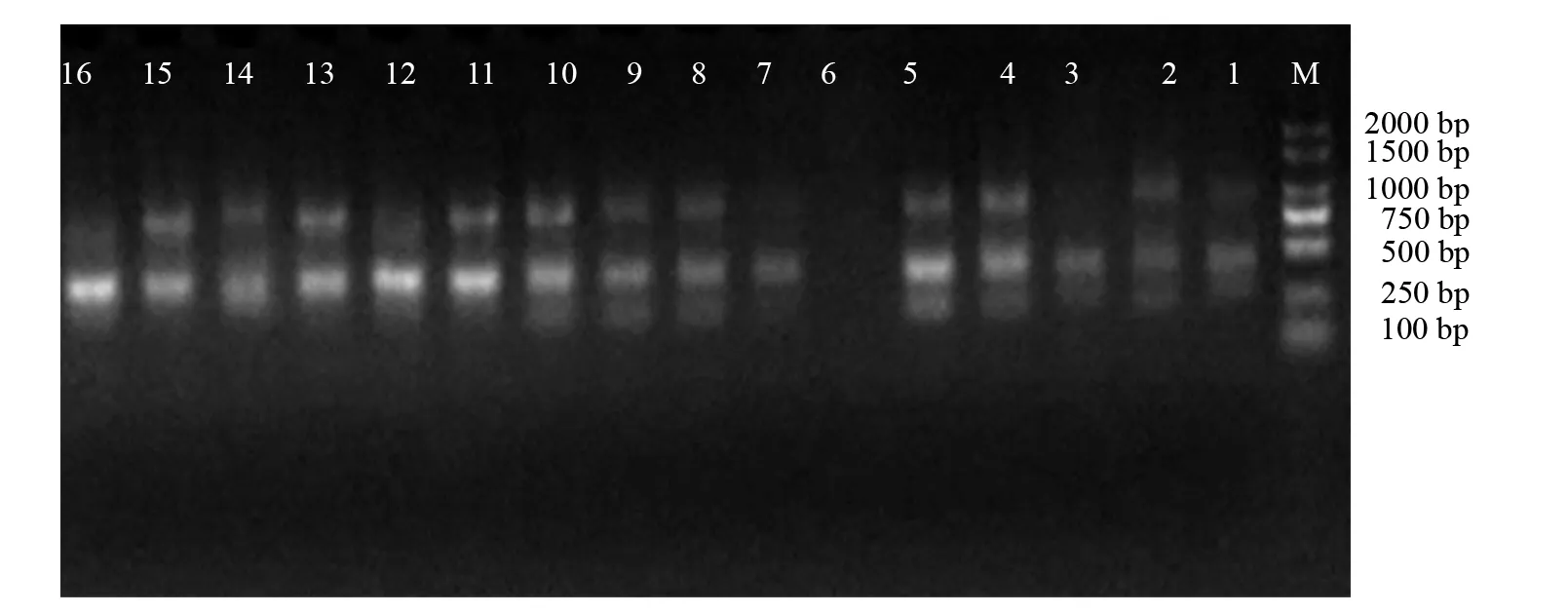
图1 引物UBC841对三裂叶豚草新疆新源县杏花村居群的扩增图谱Fig.1 Amplification map of primer UBC841 on A. trifida populations in Xinghua Village, Xinyuan County, Xinjiang 1~16表示该居群16个样品的编号,M表示DNA marker DL2000。 1-16 indicates the number of the 16 samples of the population, and M indicates DNA marker DL2000.
2.2 豚草和三裂叶豚草居群遗传多样性分析
从表2可以看出,豚草8个居群的多态位点率平均值为98.68%,其中1、3、5、6、7和8居群的多态位点率为100%;平均有效等位基因数为1.7788,其中8居群最高;各居群 Nei′s 基因多样性指数介于0.4016~0.4619之间,其变化趋势与有效等位基因数的变化趋势一样;8个居群的Shannon′s信息指数平均值为0.6194。6个遗传多样性指标的变化表明,豚草在物种水平上具有较高的遗传多样性。三裂叶豚草7个居群的多态位点率平均值为98.50%,其中10、13和14居群达到100%;平均有效等位基因数为1.7965,其中15居群最高,13居群最小;各居群Nei′s基因多样性指数介于0.4281~0.4478,其变化趋势与有效等位基因的变化趋势一样,7个居群的Shannon′s信息指数平均值为0.6224。6个遗传多样性指标的变化表明,三裂叶豚草在物种水平上具有较高的遗传多样性。

表2 豚草和三裂叶豚草居群遗传多样性分析Table 2 Analysis on genetic diversity of A. artemisiifolia and A. trifida populations
2.3 豚草和三裂叶豚草不同居群遗传分化
由表3可知,豚草总遗传多样性为0.4788,居群内基因多样性平均值0.4319;8个居群的基因分化系数为0.0981,说明总的变异中9.81%的变异来自于居群间,90.19%的变异来自于局群内;基因流为4.5994,表明豚草居群间基因流具有一定的流动性。三裂叶豚草总遗传多样性为0.4670,居群内基因多样性平均值为0.4348;7个居群的基因分化系数为0.0688,说明遗传变异中有6.88%的变异来自于居群间,93.12%的变异来自于居群内;基因流为6.7662,表明三裂叶豚草居群间基因流具有较高的流动性。

表3 豚草和三裂叶豚草的Nei′s基因多样性分析Table 3 Nei′s genetic diversity analysis of A. artemisiifolia and A. trifida
2.4 UPGMA聚类分析
由图2可知,系统聚类时遗传一致度为0.89~0.97。豚草贵州惠水县龙洞村居群和广西平阳镇平阳村居群为一支,广西大湾镇石山村居群和广西金鸡乡鱼步村居群为一支,然后合为一支再与新疆察布查尔锡伯自治县博孜墩村居群聚为一支;新疆新源县铁木里克村居群、新疆新源县玉什开普台尔村居群和新疆新源县哈拉西克村居群聚为另一支。当遗传相似系数为0.91时可分为2组:贵州惠水县龙洞村居群、广西平阳镇平阳村居群、广西大湾镇居石山村居群、广西金鸡乡鱼步村居群和新疆察布查尔县博孜墩村居群为一组;新疆新源县铁木里克村居群、新疆新源县玉什开普台尔村居群和新疆新源县哈拉西克村居群为另一组。豚草聚类结果与遗传距离的结果一致,各居群间相似系数高,相似性大。
三裂叶豚草新疆新源县阿克其村居群和新疆新源县沙哈村居群聚为一支;辽宁省沈阳市沈河区居群和新疆新源县铁热克阿吾孜村居群聚为一支,后与新疆新源县图尔根农场居群聚为一支;新疆新源县则克台村居群和新疆新源县杏花村居群聚为一支。当遗传相似系数为0.91时,7个居群分为2组:新源县阿克其村居群和新疆新源县沙哈村居群分为一组;辽宁省沈阳市沈河区居群、新疆新源县铁热克阿吾孜村居群、新疆新源县吐尔根农场居群、新疆新源县则克台村居群和新疆新源县杏花村居群为另一组。三裂叶豚草聚类结果与遗传距离的结果一致,各居群间相似系数高,相似性大(图2)。
2.5 遗传距离与地理距离间的相关性分析
豚草和三裂叶豚草居群间遗传距离与相应地理距离的相关性回归分析如图3所示,豚草的回归方程为Y=2E-0.5X+0.0692,利用SAS软件进行的相关性分析可知其相关系数R2=0.4860>r0.05(P=0.001<0.05)。豚草各居群间的遗传距离与地理距离间有显著的相关关系,居群间的基因交流受到地理距离的限制。三裂叶豚草的回归方程为Y=-7.958X+1.895,利用SAS软件进行的相关性分析可知其相关系数R2=0.0694
3 讨论
沙伟和周福军(1999)、沙伟等(2000)采用水平切片淀粉凝胶电泳测定了东北地区分布不同生态环境条件下7个豚草居群的遗传结构,揭示了东北豚草7个居群间的变异仅占总变异的7.3%,大多数(92.7%)变异存在于居群内部。邓旭等(2011)应用ISSR分子检测技术研究了5个湖南豚草居群的遗传多样性,总遗传变异中有29.61%是存在于居群间的,居群内遗传变异占总变异的70.39%。黄久香等(2012)等应用ISSR技术对广东豚草居群进行遗传多样性分析,67.37%的遗传变异发生在居群内,32.63%的遗传变异发生在居群间。本研究对新疆地区豚草和三裂叶豚草研究结果表明,遗传变异主要发生在居群内部,这与上述研究结果一致。
遗传多样性分析表明,无论是多态性位点比率,还是居群的相似性指数均较高,2物种可能来源于同一源种群,是导致其遗传相似度相对较高的主要原因。新疆地区豚草遗传相似度和广西、贵州较高,因此入侵新疆的豚草可能来源于广西、贵州。

图2 豚草和三裂叶豚草UPGMA聚类结果Fig.2 UPGMA clustering results of A. artemisiifolia and A. trifida
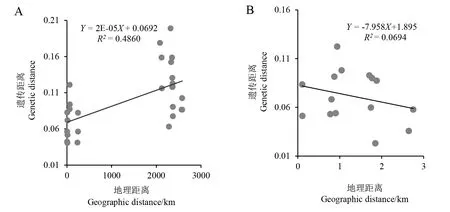
图3 豚草(A)和三裂叶豚草遗(B)传距离和地理距离的回归分析Fig.3 Regression analysis of genetic distance and geographical distance between A. artemisiifolia (A)and A. trifida(B)
本研究发现,新疆地区豚草和三裂叶豚草各居群遗传距离较近。这一现象可能是由于豚草和三裂叶豚草为一年生草本植物,其结实量大,存活率和增殖率都很高;同时伊犁河谷属农牧交错区,牛羊等牲畜的携带以及人为活动使其在短期内迅速蔓延扩展,导致各居群的遗传距离较近。由于新疆地区豚草发生分布面积相对较大,三裂叶豚草发生分布相对较小,导致三裂叶豚草各样点的距离相对较近,可能是豚草遗传距离和地理距离有显著相关性,三裂叶豚草无显著相关性的一个重要原因。因此,要真正揭示新疆三裂叶豚草和豚草来源以及遗传特性还需要研究更大范围更多数量的居群。
4 结论
本研究从豚草和三裂叶豚草中共筛选出13条多态性引物,豚草获得304条特征条带,三裂叶豚草获得266条特征条带。ISSR分析结果表明:豚草和三裂叶豚草在居群间具一定的遗传稳定性;居群内具有丰富的遗传多样性;遗传变异主要来源于居群内部。豚草遗传距离和地理距离有显著相关性,三裂叶豚草无显著相关性。本研究可为新疆地区豚草和三裂叶豚草的防治和进一步的研究提供理论基础。
猜你喜欢
植物研究(2022年5期)2022-10-12
河北科技师范学院学报(2022年2期)2022-08-26
世界科学技术-中医药现代化(2022年3期)2022-08-22
江西农业学报(2022年5期)2022-08-10
昆明医科大学学报(2022年2期)2022-03-29
小学生作文(低年级适用)(2022年3期)2022-03-23
昆明医科大学学报(2021年6期)2021-07-31
昆明医科大学学报(2021年3期)2021-07-22
浙江中医药大学学报(2021年6期)2021-07-12
黄河之声(2021年1期)2021-03-29
