
猪粪辅料促进茄果类蔬菜废弃物堆肥品质的微生物机理
2022-05-29 12:33:36陆晓林杨玉欣洪春来朱为静朱凤香姚燕来洪磊东王卫平
农业环境科学学报 2022年5期
陆晓林,杨玉欣,2,洪春来,朱为静,朱凤香,姚燕来,洪磊东,王卫平*
(1.浙江省农业科学院环境资源与土壤肥料研究所,杭州 310021;2.西北农林科技大学资源环境学院,陕西 杨凌 712100)
随着全球人口和设施园艺作物种植面积的持续增长,大量蔬菜类废弃物的产生对自然环境造成了潜在风险。我国蔬菜类废弃物年产生量超过3.6 亿t并呈连年递增的趋势。高水分含量的蔬菜废弃物若未得到合理处置将引发一系列生态安全问题。茄果类蔬菜作物富含维生素、酚和抗氧化剂等营养物质,在我国各地被广泛种植,因此茄果类蔬菜废弃物在整个设施蔬菜废弃物中占比巨大。堆肥被认为是处理茄果类蔬菜废弃物并将其转化成有机肥的有效手段。
自然堆肥条件下蔬菜秸秆料堆土著微生物的代谢活性低,限制了物料的降解效率,造成发酵周期较长。大量研究表明,在秸秆堆肥系统中接种适量的外源性动物粪便能够显著提高堆肥产品的稳定性和安全性。从养分特性和粪源微生物特性出发,猪粪是性能较好的辅料。国内外学者在畜禽粪便辅料改善蔬菜类废弃物的堆肥腐熟度方面进行了研究,例如高宁等以番茄秸秆废弃物为原料开展堆肥试验,发现猪粪辅料的添加能够提高堆体发酵温度并延长高温期。MENG 等发现猪粪辅料的添加有利于堆肥品质的提高。而堆肥效率与品质常取决于嗜热期微生物的群落功能。PICRUSt 可以用来预测堆肥过程中微生物群落的代谢功能,且准确度较高。因此,阐明微生物代谢及其生化特性有利于解析微生物行为下驱动料堆基质高效转化和快速稳定化的工作机制。有学者发现微生物在感知堆肥环境的变化后可通过调控新陈代谢的多样性来促进自身的生长,这种代谢功能多样性上的差异更会影响堆肥进程。然而目前从微生物代谢角度分析猪粪辅料促进茄果类蔬菜废弃物堆肥腐熟的机理鲜有报道。
本研究设置了两组茄果类蔬菜废弃物堆肥试验,其中处理组添加新鲜猪粪,通过测定堆肥过程中理化指标评价堆肥腐熟度和肥效,并运用高通量测序技术对堆肥过程中微生物群落结构演替进行分析,用PICRUSt 和相关性热图分析不同处理下堆肥微生物组代谢功能特性及其影响因素。研究结果加强了对堆肥系统代谢组学的认识,并为动物粪便添加促进堆肥品质提供了新的视角。
1 材料与方法
1.1 试验材料
供试材料为茄果类蔬菜废弃物(以番茄秸秆残体为主)和新鲜猪粪。番茄秸秆残体取自浙江省杭州市当地蔬菜种植基地,将秸秆粉碎为粒径2~5 cm 的小段后混匀待用。猪粪取自浙江省杭州市萧山汇仁肥料公司,将收集的猪粪统一混匀。
1.2 试验设计
堆肥试验于2018年7—9月在萧山汇仁肥料公司中试车间进行,持续45 d。试验分为两组:处理一(E处理),180 kg番茄秸秆自然堆肥处理;处理二(E-PM处理),180 kg番茄秸秆中加入猪粪辅料,辅料添加量为总堆体质量的10%。供试材料和各试验处理的初始理化性质见表1,均符合堆肥要求。将物料混匀后制堆,长×宽×高为0.9 m×0.8 m×0.8 m。为了确保堆料获得充足的氧气和把物料充分混匀,采用人工翻堆的方式进行翻堆,一周2次,即堆肥第0、3、6、9、10、12、15、18、21、23、25、28、32、36、40、44 天分别翻堆,并在堆肥第10天和第23天对各堆体进行表面喷施补水2 kg。在堆肥第0、2、9、16、23、31、38、45 天取样,将整个条垛平均分成3 段,每段采取等深度等量取样法,在每段的3 个位点(20~60 cm)随机采集等量样品,混合均匀后,为保证取样的代表性,采用四分法多次分取样品600 g(鲜质量)左右。样品分为3 份,其中一份于4 ℃保存并尽快测定含水率(Moisture content,MC)、pH、电 导 率(Electrical conductivity,EC)、发芽指数(Germination index,GI)和酶活性,一份于-80 ℃保存用于分析微生物群落多样性和结构,第三份自然风干,粉碎后用于理化性质的测定。
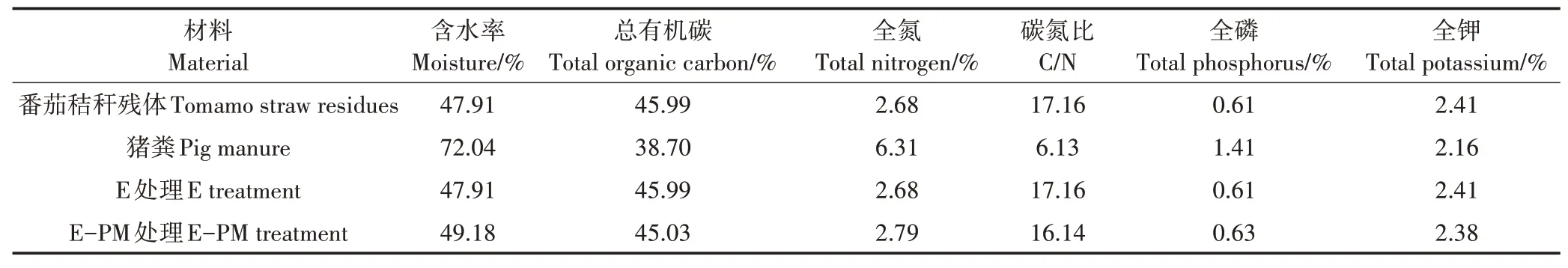
表1 供试材料的理化性质Table 1 Properties of the raw materials used in composting
1.3 测定指标与方法
1.3.1 堆肥不同时期理化指标的测定
堆体温度和环境温度的测定:将已校准的酒精温度计垂直插入堆体内部(约30 cm)测定堆体温度的变化,于每日8:00—9:00检测3次并取平均值记录。含水率采用105 ℃烘干称质量的方法测定。堆肥有机质(Organic matter content,OM)、全氮(Total nitrogen,TN)、全磷(Total phosphorus,TP)和全钾(Total potassium,TK)含量采用《有机肥料》(NY 525—2012)中的标准方法测定。新鲜样品与蒸馏水按照1∶10(/)混匀后用于pH 和电导率的测定。碳氮比为同一样品的总有机碳和全氮的比值。萝卜种子(浙农白雪春2号)用于测定堆肥鲜样的发芽指数。
1.3.2 堆肥过程中酶活性的测定
堆肥新鲜样品蛋白酶活性、纤维素酶活性及多酚氧化酶活性分别采用茚三酮比色法、水杨酸比色法及邻苯三酚比色法进行测定。蛋白酶活性以每日每克测定样品中生成氨基氮的毫克数表示(mg·g·d)。纤维素酶活性以每日每克测定样品中生成葡萄糖的毫克数表示(mg·g·d)。多酚氧化酶活性以每日每克测定样品中生成邻苯三酚的毫克数表示(mg·g·d)。
1.3.3 堆肥过程中微生物群落结构的测定
采用PowerSoil DNA Isolation Kit 提取堆肥样品中总DNA。PCR 产物使用1%琼脂糖凝胶和NanoDrop 2000 分光光度仪(Thermo Fisher Scientific,Waltham,MA,美国)进行DNA 浓度、纯度及完整性检测。随后分别使用引物338F-806R 和ITS1F-ITS2R 对细菌16S rRNA 和真菌18S rRNA 扩增,采用Illumina MiSeq PE300平台进行高通量测序。所有的PCR 扩增、文库准备与检测及上机测序分析均由上海美吉生物医药科技有限公司完成。此外,OTU分析和测序数据分析均在生物信息学云平台上(http://www.cloud.majorbio.com)完成。
1.4 数据处理与统计学分析
采用SPSS 17.0 和Originpro 2017c 对堆体理化性质和酶活性数据进行统计分析并绘图。同一取样时间下,不同数据组间的差异显著性采用Student's two-sample检验(<0.05 表示差异显著)。PICRUSt2和KEGG数据库用来预测细菌群落的代谢功能。Spearman 相关性分析用于鉴定相对丰度前10的优势细菌菌门与环境因子和群落代谢功能之间的相关性。试验数据以均值±标准误差(=3)表示。
2 结果与分析
2.1 猪粪添加对茄果类蔬菜废弃物堆肥理化性质和发芽指数的影响
2.1.1 猪粪添加对堆肥温度和发芽指数的影响
两个处理的堆体建立时,初始的堆体温度和环境温度均为36 ℃(图1a),但堆温上升较快。E-PM 处理的堆温在第1 天可达到60 ℃,而E 处理在第2 天堆温才达到60 ℃。此外,E-PM 处理和E 处理的嗜热期(>60 ℃)分别为12 d和8 d,且E-PM 处理的堆温大于E处理的时间为35 d,两者的最高堆温分别为66 ℃和65 ℃。伴随翻堆,整个堆肥过程存在二次发酵的现象,最终在刚进入降温期(<50 ℃)时,E-PM 处理堆体温度下降幅度较大,由51 ℃降低至41 ℃,而E处理仅从50 ℃降低至48 ℃。在堆肥前16 d,发芽指数一直小于60%(图1b),在第45 天,E-PM 处理的发芽指数达到82.11%,且显著高于E 处理(<0.05),这一现象充分证明添加猪粪后堆肥产品浸提液的促生能力提高。
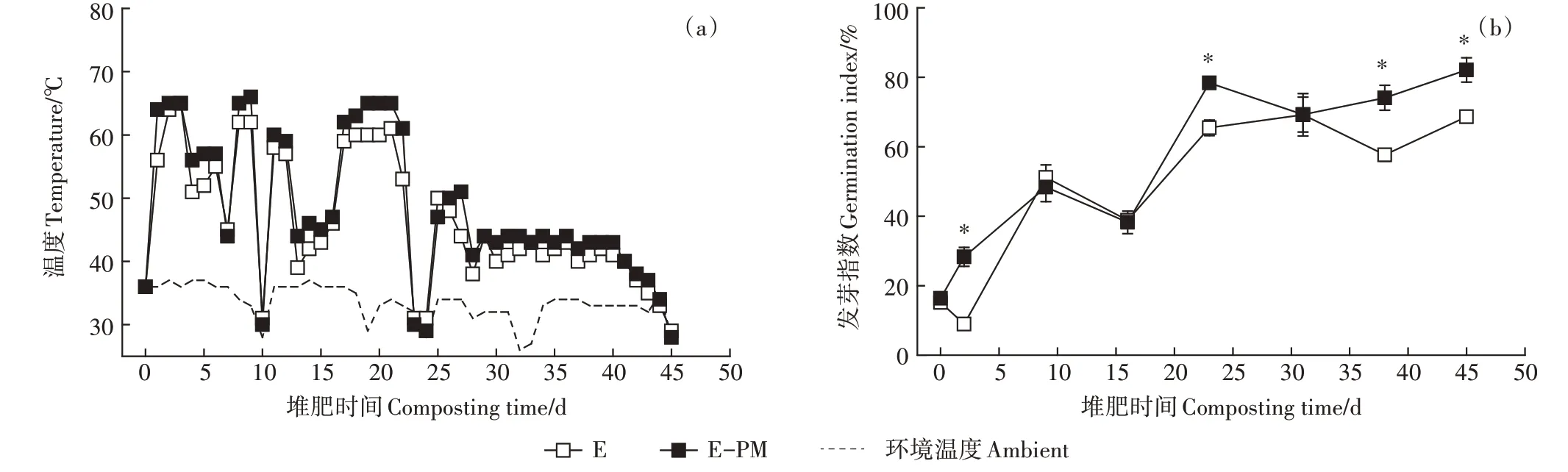
图1 堆肥过程中温度和发芽指数的变化Figure 1 Changes of temperature and germination index during composting process
2.1.2 猪粪添加对堆肥理化性质的影响
图2a 表明,由于堆肥前期反应剧烈,有机物水解生成的水分高于蒸发的水分,所有处理堆肥前期(0~16 d)的含水率均有上升的趋势,随后呈波动下降的趋势。最终E-PM 和E 处理的含水率分别为60.49%和61.11%,需要进一步对产品脱除水分从而达到土地利用要求。电导率在堆肥初期有一个短暂的下降阶段,随后呈先上升后降低的趋势(图2b)。与E处理相比,添加猪粪显著降低了茄果类蔬菜废弃物堆肥过程中的电导率(<0.05)。E-PM 处理和E处理初始堆体的pH 分别为9.31 和9.24,随后总体呈先降低后上升的趋势(图2c),添加猪粪对堆肥后期(在31~45 d)的pH 无显著影响(<0.05)。从图2d 可知,在堆肥初期,E-PM 处理和E 处理的有机质含量分别为77.63%和79.28%;堆肥至第23 d,其有机质含量分别为75.24%和82.82%;而堆肥结束时,其有机质含量分别67.84%和69.71%。因此,相比于堆肥前,E-PM 处理和E 处理有机质平均降解率分别为12.61% 和12.07%,这一结果表明添加猪粪可加快堆肥早期有机质的降解并促进整个堆肥过程中有机质的降解。所有处理全氮含量的变化总体呈先降低后上升的趋势(图2e),E-PM 处理在0~23 d 全氮含量均显著高于E 处理(<0.05),最终E-PM 和E 处理的全氮含量分别为2.66%和2.68%(>0.05)。图2f表明添加猪粪显著降低了第0~23 d 的料堆碳氮比(<0.05)。初始的全磷和全钾含量在处理之间无显著差异(图2g 和图2h),这主要受到供试原料、料堆含水率、料堆密度、质地及采样等多因素影响。但随着堆体原料被微生物不断分解,E-PM和E处理的料堆体积不断减小,堆体中全磷和全钾的相对含量逐步增加,出现典型的“富集效应”。堆肥过程中处理之间的全磷和全钾含量差异显著(<0.05),在堆肥结束时,E-PM处理的全磷和全钾含量均显著高于E 处理(<0.05),分别高达1.22%和2.99%,表明猪粪的添加显著提高了肥效。
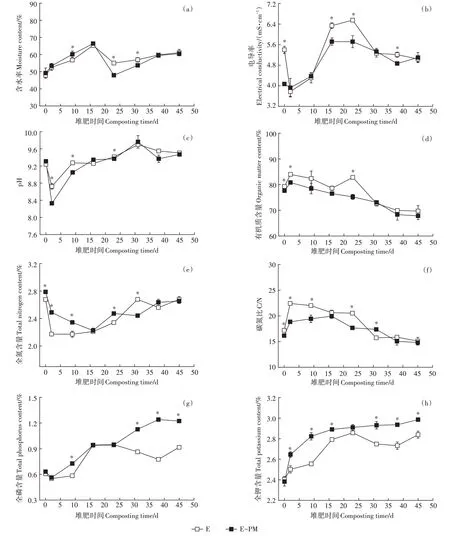
图2 不同处理下茄果类蔬菜废弃物堆肥过程中理化性质的动态变化Figure 2 Dynamic of physicochemical parameters during solanaceous vegetable wastes composting under different treatments
2.2 猪粪添加对茄果类蔬菜废弃物堆肥酶活性和微生物群落结构的影响
2.2.1 猪粪添加对堆肥酶活性的影响
总体来说,所有处理堆肥后期蛋白酶活性均高于堆肥前期(图3a)。伴随着堆肥的进行,E-PM 处理和E 处理的蛋白酶活性均呈先降低后升高的趋势,且在嗜热期(第2 天和第9 天)均表现为E-PM 处理显著高于E 处理(<0.05)。值得注意的是,在腐熟期E-PM处理的蛋白酶活性显著低于E 处理(<0.05)。EPM 和E 处理的多酚氧化酶活性在堆肥过程中总体呈先升高后降低再升高的趋势(图3b),最大值分别出现在第2 天(1.725 mg·g·d)和第38 天(1.974 mg·g·d)。类似地,猪粪的添加也仅促进了堆肥嗜热期多酚氧化酶的活性,这有利于加速堆料的腐殖化进程。对于堆体纤维素酶活性,相比于E 处理,猪粪添加显著促进嗜热期纤维素酶活性的升高(<0.05),且所有处理纤维素酶活性最大值均出现在第9 d(图3c)。而随着堆体温度的降低,E-PM 处理的纤维素酶活性显著低于E处理。
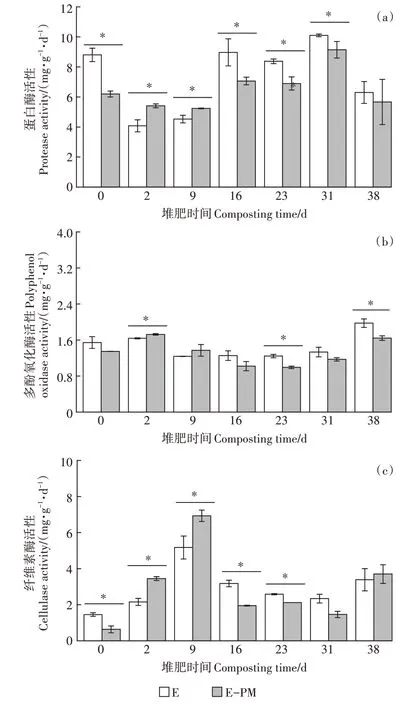
图3 堆肥过程中蛋白酶活性、多酚氧化酶活性和纤维素酶活性的变化Figure 3 Changes of protease activity,polyphenol oxidase activity,and cellulase activity during composting process
2.2.2 猪粪添加对堆肥微生物群落结构的影响
由表2 可知,对于细菌群落结构和真菌群落结构,E-PM 处理和E 处理的Shannon指数和Chao1指数在第2天和第23天均无显著差异。在第38天,相比E处理,E-PM 处理仅显著促进了Shannon 指数的增加(<0.05),这表明猪粪添加对腐熟期微生物群落结构的多样性有显著影响,而对微生物群落结构的丰富度无显著影响。

表2 堆肥过程中所观测到的OTU数和微生物群落的α多样性指数Table 2 Observed OTU number and Alpha diversity indices in three sampling stages with different treatments
为了寻找微生物在属水平上各组样本之间优势物种的差异,选取在属分类水平上平均相对丰度前15 的物种,见表3。本试验结果表明,在堆肥第2 天,E 处理和E-PM 处理相对丰度最高的优势细菌均为(芽孢杆菌属,22.7%~23.7%),优势真菌均为(曲霉菌属,74.4%~80.7%)。此外,相比于E 处理,此阶段(梭菌属)的相对丰度在E-PM 处理中显著提高,而(糖多孢菌属)的相对丰度显著降低(<0.05)。在堆肥第23 天,所有处理中和的相对丰度明显降低,(枝顶孢霉菌属)的相对丰度大幅增加(17.6%~58.0%)。值得注意的是,猪粪添加显著提高了此阶段下(大洋芽孢杆菌属)、和的相对丰度,却显著降低了(微囊菌属)和的相对丰度(<0.05)。在堆肥第38 天,、、(纤细芽孢杆菌属)、(嗜热霉菌属)和在E-PM 处理的 相 对丰度显著 高于E 处理(<0.05),而(糖单胞菌属)、、、(远洋杆菌属)、和的相对丰度显著低于E处理(<0.05)。
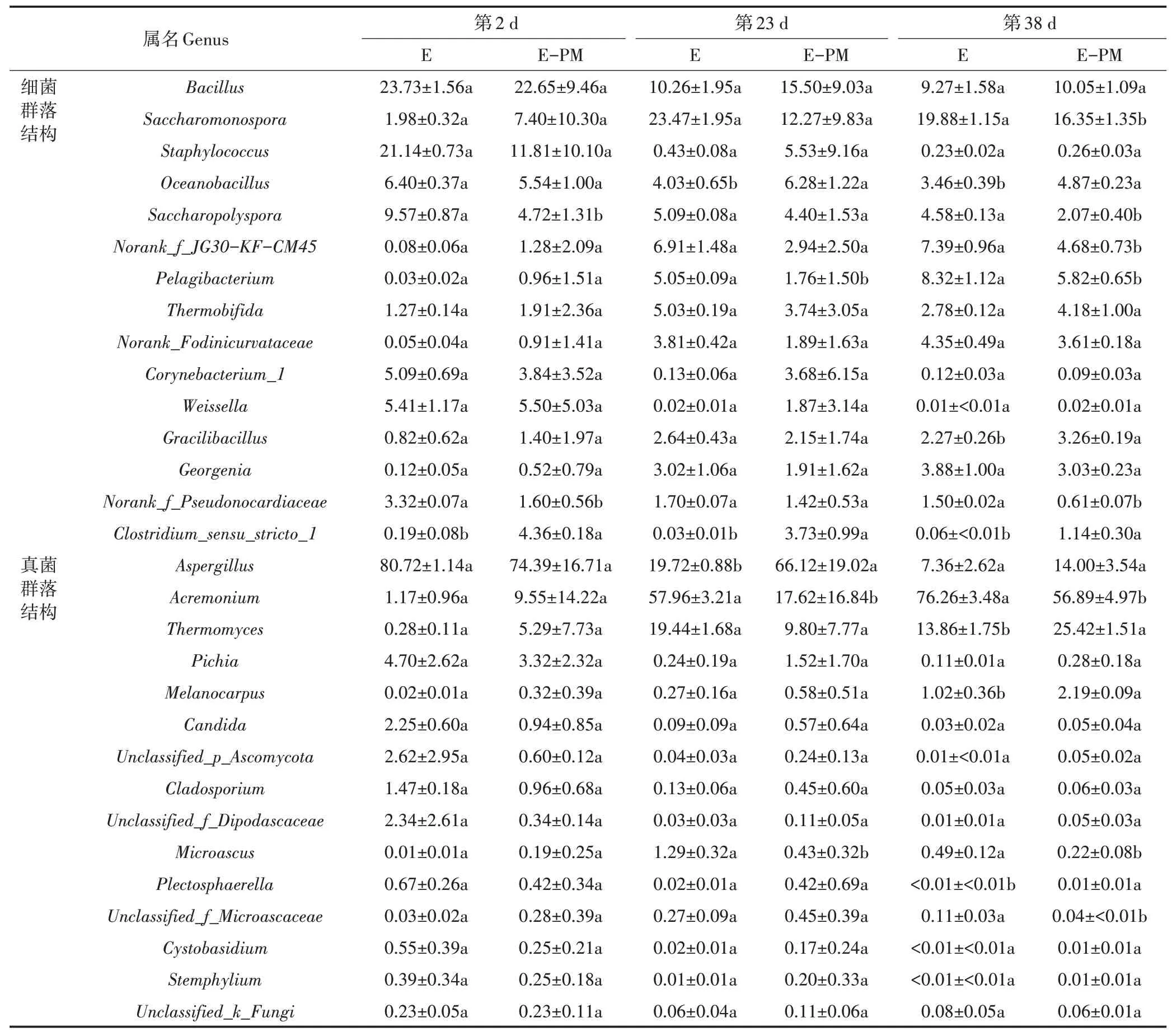
表3 堆肥过程中优势菌群在属水平的相对丰度(%)Table 3 The relative abundance of dominant bacterial community at the genus level during composting process(%)
2.3 基于PICRUSt 探索微生物功能代谢对猪粪添加的响应
本研究利用不同处理下嗜热期细菌群落生物化学代谢途径和细菌代谢功能来探索堆肥物料有机物质降解的潜在机制(图4 和图5)。由图4a 可知,猪粪添加对嗜热期茄果类蔬菜废弃物堆肥系统中的微生物生物代谢具有明显的促进作用。与此同时,基于KEGG 代谢通路进行预测发现,被注释到代谢和细胞进程的基因序列丰度在E-PM 处理中有所提高,较E处理分别增幅5.68%和10.26%。此外,由图4b 可知,在KEGG 二级函数水平下,相比于E 处理,20 条代谢功能谱相关序列的丰度在E-PM 处理组更高,特别是代谢通路下的碳水化合物代谢和氨基酸代谢相关序列的丰度。在基因信息处理、环境信息处理和细胞进程下的多条代谢功能谱相关序列的丰度同样在EPM 处理中明显增加。这些结果表明,添加猪粪较快地改善了微生物细胞的加工功能,包括转录、翻译及复制等强大功能,这有助于具有纤维素降解能力和氨基酸代谢能力的细菌生长繁殖,还有利于膜转运相关代谢活性的增强。

图4 堆肥过程中细菌功能谱差异的PICRUSt分析Figure 4 Variation in bacterial function profiles during composting process analyzed by PICRUSt
KEGG 途径分析表明(图5a),与E 处理相比,所选择的7 种与有机物质降解相关的功能酶序列丰度在E-PM 处理组明显富集。因此,E-PM 处理下堆肥嗜热期内产生的酶量较多,这有利于堆肥物质的高效降解,从而产生更多有效的活性养分促进微生物菌群的生长。此外,对嗜热期产温室气体甲烷(CH)的潜力功能进行评估,结果表明(图5b),辅酶420-0 氢化酶的序列丰度在E-PM 处理组低于E 处理,而另外3种酶的序列丰度在E-PM 处理组高于E 处理。因此,可以推测施加猪粪降低了嗜热期产甲烷的酶数量并提高了氧化甲烷的酶数量,从而可能有效减少堆肥化进程中甲烷的排放。
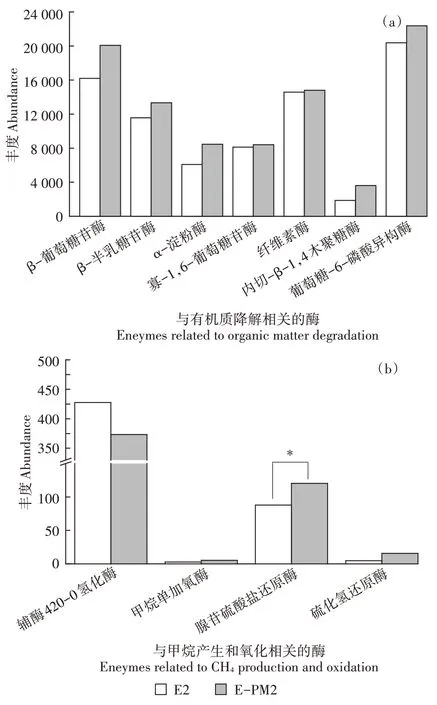
图5 相关酶基因丰度的PICRUSt预测Figure 5 Abundance of related enzymes predicted by PICRUSt
2.4 优势菌门分布及其与环境因素和代谢功能的相互关系
相关性分析(图6b)表明,堆肥发芽指数、含水率和全氮含量均与Bacteroidetes(拟杆菌门)、Deinococcus-Thermus 和Planctomycetes(浮霉菌门)之间呈正相关(<0.05)。此外,与含水率、pH 和全磷含量之间呈显著正相关(<0.05)的Proteobacteria(变形菌门)和Chloroflexi(绿湾菌门)又与氨基酸代谢、碳水化合物代谢、能量代谢和脂类代谢之间呈极显著正相关(<0.001),表明含水率、pH 和全磷含量对这些菌群的代谢产生深远的影响。特别地,多种代谢通路均与Proteobacteria 和Chloroflexi 等8 个菌门之间呈显著正相关,而Firmicutes(厚壁菌门)等2 个菌门却与这些代谢功能特性之间呈极显著负相关(<0.001),表明代谢能影响细菌群落的变化。因此,相比于Firmicutes,嗜热期独特的堆肥环境促进了E-PM处理中Proteobacteria和Chloroflexi的大量生长(图6a),而这些优势菌又参与了多条代谢途径并有助于茄果类蔬菜废弃物的高效堆肥。
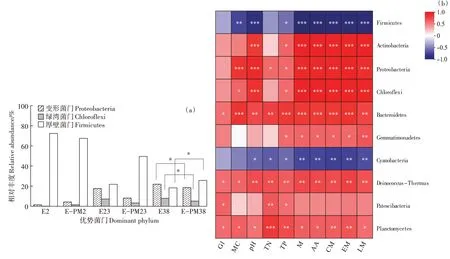
图6 优势菌门的分布特征及其与环境因子和代谢功能的相关性分析Figure 6 Analysis of distribution characteristics of dominant phylum and their correlation with environmental factors and metabolic function
3 讨论
3.1 粪源辅料促进料堆堆肥品质及其微生物学机制
相比于单一物料自然堆肥,多种原料复配进行共堆肥通常可以促进堆肥物料的降解并提高堆肥的品质,这得益于共堆肥体系下微生物总量的多样化和适合微生物生长的养分更加全面。本试验结果表明,猪粪的添加促进了物料嗜热期的酶活性和有机物质的生物降解率,进而加速堆体腐熟并提高产品肥效。已有研究也表明,蚓粪的接种显著增加了麦秸料堆早期Firmicutes 和Betaproteobacteria 的相对丰度,改善了关键种的生存环境,并且增加了与碳水化合物代谢、氨基酸代谢和能量代谢相关基因的相对丰度。MENG 等研究发现,在菌糠和稻壳的共堆肥中猪粪的添加提高了堆体(假单胞菌属)的多样性和和的相对丰度,这些菌群在促进堆肥基质有效降解和降低物料植物药害性上发挥着重要作用。此外,碳水化合物代谢和氨基酸代谢处于细菌群落代谢功能谱的中心环节,细菌在嗜热期常分泌多种水解酶用于底物的生物降解。本研究发现添加猪粪提高了嗜热期菌群的代谢能力并促进了有机碳的分解。对此相应潜在的微生物机制可能是:作为能源和碳源的氨基酸在堆肥物料降解过程中产生,一方面氨基酸被微生物摄取助其活性的提高,另一方面氨基酸提前参与腐殖质的合成并改善堆肥养分状况。
3.2 粪源辅料对堆肥土著菌群的影响及其影响因素
据报道,新鲜猪粪携带了高丰度的Firmicutes、、、和优势生理群,进而对茄果类秸秆堆肥过程中的土著菌群造成显著影响。因此,相比于E 处理,可能来源于猪粪的___在E-PM 处理嗜热期显著富集(<0.05)。除此之外,嗜热期Proteobacteria 和Chloroflexi的相对丰度同样在E-PM 处理中较高,而Firmicutes 的相对丰度却有所下降。据报道,Proteobacteria、Chloroflexi 和均有效参与木质纤维素的生物降解途径并使料堆品质提高,这些高代谢活性的微生物生理群可能在E-PM 处理的早期显著促进了堆体有机质的降解。在堆肥后期,猪粪的添加显著增加了、___和的 相 对 丰度,但由于堆体腐熟度的差异使E-PM 处理的菌群代谢活力和酶活性低于E处理。此外,包含多种植物病原菌的的相对丰度在E-PM 处理显著低于E处理,表明添加猪粪有助于提升后期产品的安全性。相关性分析表明,关键的菌群代谢功能谱主要由Proteabacteria 和Chloroflexi 主导,并且这些微生物物种的生长代谢受到水分、pH 和全磷含量的调控。譬如堆体pH 的变化会改变养分的有效性,进而影响Proteabacteria 和Chloroflexi 的丰度。本试验发现猪粪的添加降低了堆体嗜热期的pH 以及Firmicutes 的相对丰度,先前研究表明低pH 可减少氨挥发并降低Firmicutes 与氨气排放两者之间的正相关关系,因而解释了E-PM 处理在第2 天至第9 天料堆全氮含量显著高于E处理的原因。
3.3 粪源辅料调控菌群代谢功能谱
碳水化合物代谢和氨基酸代谢为微生物的生长提供了能源和碳源,并高效驱动着堆肥基质中木质纤维素的分解和腐殖质的合成。本试验结果表明,猪粪的添加极大地促进了嗜热期堆体微生物群落中与碳水化合物代谢和氨基酸代谢相关的序列丰度,其有利于茄果类蔬菜废弃物有机物质的降解和腐殖质的合成。β-葡萄糖苷酶(EC:3.2.1.21)、β-半乳糖苷酶(EC:3.2.1.23)、α-淀粉酶(EC:3.2.1.1)、寡-1,6-葡萄糖苷酶(EC:3.2.1.10)、纤维素酶(EC:3.2.1.4)、内切-β-1,4-木聚糖酶(EC:3.2.1.8)和葡萄糖-6-磷酸异构酶(EC:5.3.1.9)与生物聚合物、纤维素及半纤维素的降解密切相关。辅酶420-0 氢化酶(EC:1.12.98.1)通常促进甲烷的生成,而甲烷单加氧酶(EC:1.14.13.25)、腺苷硫酸盐还原酶(EC:1.8.99.2)和硫化氢还原酶(EC:1.8.99.3)促进甲烷的氧化。通过PICRUSt预测发现,在嗜热期与有机质降解相关功能酶的序列丰度和与甲烷氧化相关功能酶的序列丰度在E-PM 处理中均大幅提高,这进一步表明猪粪添加不仅有利于物料基质的快速降解,而且有利于甲烷的减排。随着堆肥的进行,E-PM 处理在第23天和第38天的菌群代谢活性和堆体酶活性均低于E处理(图3 和图4a)。本研究认为动物粪便的添加提前促进了堆体的腐熟,因此微生物菌群活性在堆体稳定时并未表现较高的水平,这与在麦秸堆肥系统中蚓粪添加抑制了后期微生物菌群的碳源代谢活性相一致。由于物料氮硫转化显著影响着堆体质量,后续的研究一方面应更关注工艺上的优化(如超高温堆肥),并积极探索在猪粪与蔬菜秸秆复混体系下通过光谱及电镜技术研究氮素与硫素等相关环境废气减排机制,另一方面应联合运用多组学的手段解析物料中复杂生物有机物质的降解途径。
4 结论
(1)添加猪粪有效增加了茄果类蔬菜废弃物早期堆肥中Proteobacteria 和Chloroflexi 的相对丰度,进而促进了堆肥升温,延长嗜热期,最终显著提高产品的安全性。
(2)猪粪的加入能够有效提高堆体嗜热期细菌群落的代谢活性及有机质的降解率,促进堆体的快速腐熟。
猜你喜欢
粮食与饲料工业(2023年5期)2023-10-30 11:11:00
南京农业大学学报(2023年1期)2023-02-02 11:33:16
安徽农业科学(2022年22期)2022-12-20 07:10:46
中国农业大学学报(2022年3期)2022-05-19 00:41:08
现代畜牧科技(2021年3期)2021-07-21 08:41:48
今日农业(2020年19期)2020-12-14 14:16:52
农民致富之友(2019年7期)2019-05-23 01:14:42
猪业科学(2018年4期)2018-05-19 02:04:38
上海农业科技(2018年3期)2018-01-17 03:50:52
农家顾问(2016年11期)2017-01-06 13:01:48
