神经干细胞移植在神经退行性疾病中的研究进展
2022-05-28 09:11:12郭文文赵亚白敏张延英王颖花汪永锋师长宏
中国实验动物学报 2022年2期
郭文文 赵亚 白敏 张延英 王颖花汪永锋*师长宏*
(1.甘肃中医药大学,兰州 730000;2.空军军医大学实验动物中心,西安 710032;3.甘肃中医药大学/甘肃省道地药材质量标准化技术研究与推广工程实验室/甘肃省高校中(藏)药化学与质量研究省级重点实验室/甘肃省实验动物行业技术中心,兰州 730000;4.延安大学,陕西 延安 716000)
神经干细胞(neural stem cell,NSC)能够分化为多种细胞谱系,这些细胞谱系包括神经元、星形胶质细胞和少突胶质细胞等,在神经再生中起到重要的作用[1]。NSC 分布在整个发育中的大脑中,位于大脑中的两个主要神经源性生态位——侧脑室的脑室下区(subventricular zone,SVZ)和海马齿状回的颗粒下区(subgranular zone,SGZ)[2]。中枢神经系统(central nervous system,CNS)发生急性损伤、慢性损伤或退行性变后,内源性NSC 变得活跃并参与脑修复过程,但受伤的CNS 内的NSC 被限制在分散的位置,数量很少且被不支持神经元分化的微环境所包围,从而导致CNS 内在自我修复能力不足或无效[3]。
随着生命科学领域各项研究技术的不断成熟,对大脑进行全面深层次研究已经具备可能。其中,NSC 移植是近年来脑研究领域的重要热点之一,该技术已广泛应用于各种神经性疾病的治疗研究。有报道指出移植人神经诱导性干细胞可用于治疗大小鼠模型中包括创伤性脑损伤、脊髓损伤及中风等在内的多种神经疾病[4]。当前,大多数NSC 研究依赖于啮齿动物模型。但是,人与啮齿动物的NSC有很大不同,许多疾病风险基因在二者之间表达存在较大差异。因此,基于人神经细胞嵌合小鼠的模型制备可在动物水平实现对人脑的模拟,且相关神经诱导性干细胞移植已应用于各个领域,以便更加深入了解干细胞在受伤大脑中的治疗作用的潜在机制。NSC 的体外分离分析已被证明是揭示神经发生的细胞和分子机制的重要方法,能够进一步用于基于干细胞疗法的神经系统疾病和损伤的治疗[5]。因此,外源性NSC 移植是目前治疗神经退行性疾病及研究的热点。本文就NSC 的来源及其在神经退行性疾病治疗研究进展进行综述,以期为神经退行性疾病的治疗提供新的思路。
1 神经干细胞来源
NSC 移植所需要的细胞量巨大,细胞来源已成为NSC 移植应用于临床治疗及科学研究必须解决的首要问题[6]。研究表明,具有自我更新能力的NSC 能够从啮齿动物的胚胎组织和人脑组织中分离出来[7]。目前已明确可从3 种来源获得NSC,包括从原代组织中直接提取、从多能干细胞中分化以及从体细胞中转分化(见图1)。
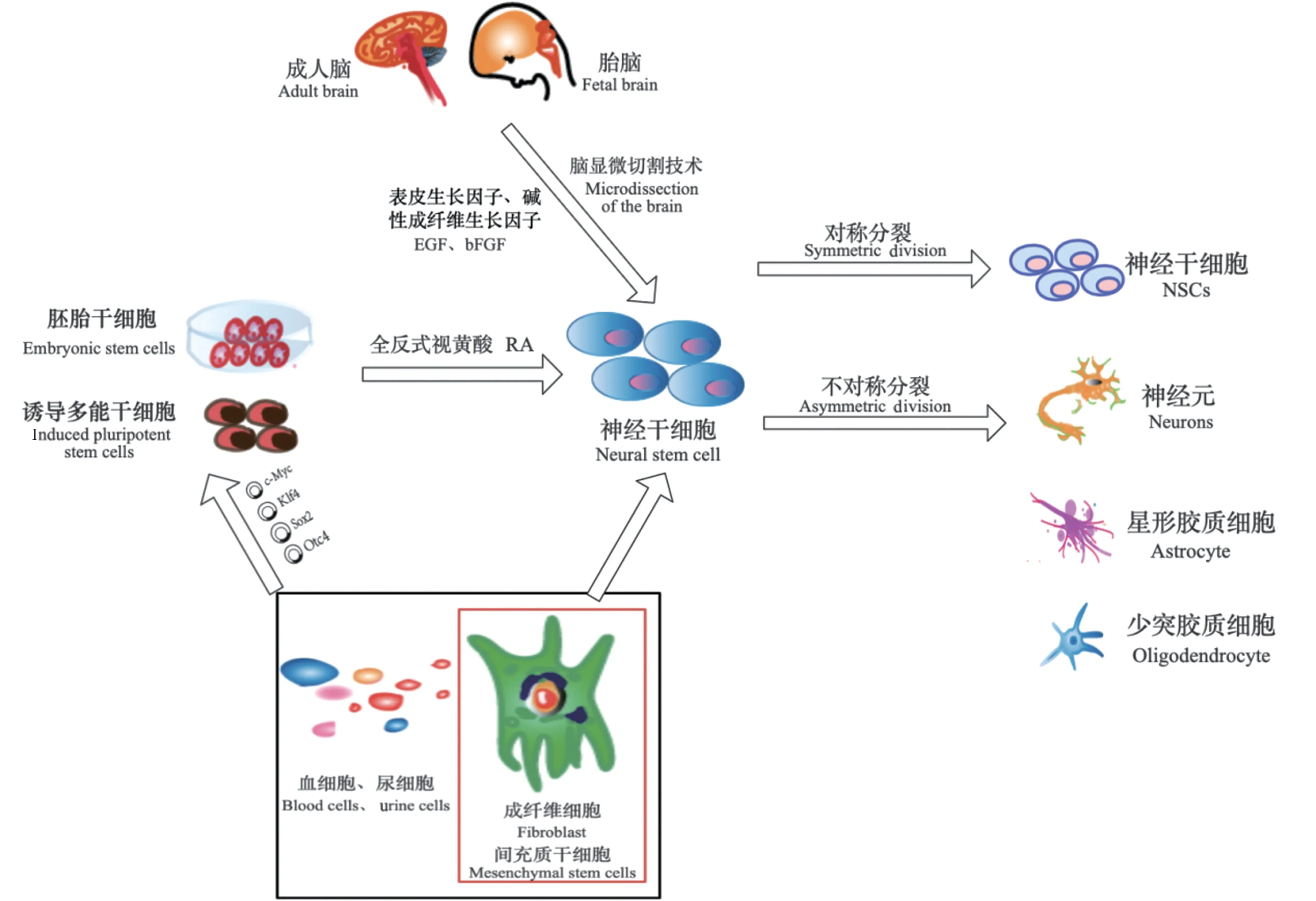
图1 神经干细胞的来源Figure 1 Source of neural stem cells
1.1 从原代组织中直接提取
从原代组织中直接提取的方法是直接从胎儿或成人的脑和脊髓组织中直接提取NSC,进而进行体外诱导分化。NSC 位于成人大脑和胚胎的不同区域,它们有产生不同类型的神经元和胶质细胞的趋势。成年哺乳动物大脑中的脑室下区和胚胎脑中的神经节隆起是富含NSC 的区域,为NSC 提供了良好的来源[8]。1992 年Reynolds 等[9]首次从成年哺乳动物脑室管膜下区分离鉴定出NSC;最近Farshad 等[10]通过改进脑显微切割技术,缩短手术时间,在适宜的环境温度和无菌条件下从胚胎小鼠颅骨中取出整个脑组织并分离出较高比例的NSC。而通过大脑纹状体和胸脊髓的显微解剖可获得原始NSC,将分离出的原始NSC 在含有表皮生长因子(epidermal growth factor,EGF)和碱性成纤维细胞生长因子(basic fibroblast growth factor,bFGF)的单细胞悬浮液中进行体外培养,可形成称为“神经球”的非粘附性球形簇,也可以选择性的在培养基中进行单层贴壁培养。但是由于提供三维环境的胚状体(embryoid body,EB)更有利于NSC 的分化和培养,因此,EB 培养是目前分离和维持NSC 最流行和最可靠的方法[11]。
1.2 多能干细胞向神经干细胞的分化
多能干细胞包括胚胎干细胞(embryonic stem cells,ESCs)和诱导多能干细胞(induced pluripotent stem cells,iPSCs),两者都可通过分化产生所需要的细胞[12]。ESC 源自植入前胚胎,具有多能性,可以分化为外胚层、内胚层和中胚层[13]。有充分证据表明,ESC 可以分化为NSC,然后分化为神经元或神经胶质[14]。诱导ESCs 向NSC 转分化的实验技术方法众多,其中,通过采用全反式视黄酸(all-trans retinoic acid,RA)诱导NSC 的方法最为经典。研究表明,RA 可以调节ESC 的生殖细胞分化和神经分化,建立并改善ESC 分化为NSC 所依赖的微环境[15]。诱导分化过程中加入RA 的时间点及浓度的不同会导致细胞命运的不同。例如,高RA 浓度会增加神经分化的速度,而低RA 浓度会诱导ESCs向心肌细胞分化;高RA 浓度诱导ESCs 为背侧表型,低RA 浓度诱导ESCs 腹侧表型[16]。然而这种方法分化效率低,可能导致组织移植物中神经过度生长或形成肿瘤,同时也包括伦理安全等问题,因此,不适用于移植治疗[17]。
2006 年,日本京都大学Takahashi 等[13]首次报道了有关诱导多能干细胞的研究,该研究使用逆转录病毒转染多能性基因(Oct3/4、Sox2、c-Myc 和Klf4),通过适当的信号输入有效地将成纤维细胞转分化为iPSCS。随着技术发展,人类iPSCs(hiPSCs)可以通过使用一组相同的多能性相关基因获得[18],从其他细胞类型如人角质形成细胞、人外周血细胞、脂肪细胞、肝细胞、胃细胞等获得hiPSCs 也成为可能[19]。Kozhich 等[20]通过在无血清和缺乏营养的培养基单层培养中,使iPSCs 经过神经“玫瑰花结”阶段直接分化为NSC[21],这种方法适用于从人类iPSCs 标准化分化和生成神经细胞。在形态学、电生理特性和基因表达谱方面,iPSCs 衍生的神经元与人类神经元具有相似的特征。因此,iPSCs 衍生的NSC 已成为细胞疗法的一个理想的来源[22]。
1.3 从体细胞中转分化
随着诱导多能干细胞(iPSCs)技术的发展,使用来自胚胎或胎儿神经组织的同种异体NSC 的研究方法正逐渐成为主流。通过重编程成体细胞获得hiPSC 是研究疾病发生和发病机制的代表性方法系统[23],然而,这种方式需要较长时间才能将这些细胞分化为功能性神经细胞,且在重编程过程中表观出遗传信息的重置。相反,将体细胞直接转分化为神经元细胞的速度更快、效率更高,细胞治疗更安全,并且可以保留捐赠者的年龄特征[24]。
转分化,也称为谱系重编程,最初是由塞尔曼和卡法托斯在1974 年创造的[25]。在这个过程中,一种类型的成熟体细胞可不经历中间多能状态直接转化为另一种类型的成熟体细胞,其主要特征在于iPSC 阶段的缺失或不明显,以及通过重编程增殖细胞(例如成纤维细胞或星形胶质细胞)直接获得NSC[24]。
研究证明谱系特异性转录因子(transcription factor,TF)的强制表达可以重新编程各种体细胞类型的发育潜力,将TF 与培养基补充剂(例如稳定生成NSC 所需的小分子和生长因子)结合使用,可实现体细胞直接转分化为NSC[26]。与iPSC 生成相比,能够转化为iNSC 的TF 的选择非常多样化。第一个直接转化策略是通过在小鼠成纤维细胞中过表达三种TF,即Ascl1、Brn2 和Myt1l(BAM 因子),以将人类成纤维细胞转化为NSC[27]。近年来,人们发现多种TF 组合可以用来从成纤维细胞产生亚型特异性蛋白,而在TF 筛选转化研究中已经发现了额外的促神经元因子,如brn3a/b/c、brn4s 和ezh2[28]。此外,使用病毒载体将外源性基因引入细胞是目前NSC 诱导转分化中最突出的方法,其中慢病毒和逆转录病毒因其能够有效地直接整合到宿主细胞的基因组中并赋予适当水平的TF 表达,是目前最常使用的病毒载体[29]。然而,TFs 的病毒传递具有不良副作用,可能导致肿瘤发生及突变,影响其临床应用[30]。除了TF 外,在体外培养中,还可利用小分子调控特异性靶点和表观遗传机制来产生神经祖细胞和神经元[31]。小分子可以介导施加病毒剂的TF 输送,以提高重编程效率。据报道,通过将细胞连续暴露于12 个小分子的组合中,可以有效且直接地将人类成纤维细胞重编程为谷氨酸能神经元,这些化学诱导神经元(ciN)显示出神经元转录网络及成熟的放电模式,并形成功能性突触[32]。
除此之外,通过向细胞培养基中添加多种因子,特别是各种生长因子,如脑源性神经营养因子(BDNF)、血小板源性生长因子(PDGF)、表皮生长因子(EGF)、成纤维细胞生长因子2(FGF-2)和全反式视黄酸(RA),能够将骨髓来源的人类间充质干细胞(Mesenchymal stem cell,MSC)重编程为神经干细胞[33]。
与多能干细胞诱导得到的神经元相比,转分化的NSC 具有明显优势,它能够在体外进行扩增,并保留产生多种神经元亚型和胶质细胞的能力[34]。而这种来自体细胞如皮肤成纤维细胞、尿细胞和血细胞转分化在临床上容易实现[35]。
NSC 作为中枢神经系统中的储备细胞,通过神经祖细胞阶段分化为神经元、星形胶质细胞和少突胶质细胞,替代凋亡和死亡细胞进行损伤后的修复。还可分泌神经营养因子、生长因子和细胞因子促进突触生长,保护现有神经细胞免受原位损伤[36]。以上3 种来源特别是从iPSCs、ESCs 中获得的NSC 和胶质细胞可帮助急慢性损伤后的神经功能恢复,用于神经退行性疾病的治疗[37]。
2 神经干细胞在神经退行性疾病治疗中的作用
神经退行性疾病(neurodegenerative diseases,ND)是一种因外周或中枢神经系统内的神经元细胞丢失而引起的疾病状态,内源性神经细胞的可修复性不足或治疗失败是治疗该类疾病的重要障碍[38]。基于干细胞的移植,特别是使用来自iPSC、ESC 或NSC 的神经元和神经胶质细胞,将成为神经退行性疾病的常规治疗方法,并且可能通过不同的机制,如神经营养因子的产生、神经炎症的降低、突触可塑性的增强和细胞的替代,对神经退行性疾病产生有益的作用[39]。这一类疾病主要包括创伤性脑损伤(traumatic brain Injury,TBI)、脊髓损伤(Spinal cord Injury,SCI)、中风(Stroke)、阿尔茨海默病(Alzheimer disease,AD) 及帕金森综合征(Parkinson,s disease,PD)等。
2.1 创伤性脑损伤
创伤性脑损伤(traumatic brain injury,TBI)是一种常见疾病,其主要损害是血脑屏障(BBB)破坏和神经元坏死[40]。血脑屏障破坏导致缺氧增加,造成神经损伤和死亡,损伤神经元释放异常神经递质导致兴奋性毒性细胞损伤和死亡,继而出现慢性炎症及进一步的细胞损伤等病理表现[41]。
研究发现,细胞治疗已成为TBI 的一种新型治疗方式,多种类型的细胞都可用于TBI 的治疗,如NSC、ESC、NPC、MSCs、人源性畸胎瘤细胞(NTera2,NT2)、多能成体祖细胞和内皮祖细胞等[42]。其中通过NSC 和MSCs 的分化促进细胞置换是TBI 后的重要治疗作用机制[43]。NSC 可通过“生物桥”在SVZ 和损伤部位之间形成通路,直接从SVZ 迁移到损伤部位并分化为神经元和神经胶质细胞,从而增强宿主神经源性细胞的迁移[44]。此外,内皮细胞通过产生可溶性生长因子,如脑源性神经营养因子(brain-derived neurotrophic factor,BDNF)和血管内皮生长因子(vascular endothelial growth factor,VEGF)刺激NSC 的增殖和分化以及成神经细胞的迁移[45]。
目前,由于NSC 和NPCs 难以大量产生,在临床试验中受到限制,而MSCs 由于易于分离、免疫抑制特性以及缺乏伦理争议的特点,被认为是具有临床应用前景的治疗细胞[46]。相关研究证明TBI 后,MSC 移植细胞可以在损伤部位存活和迁移,有助于神经保护、神经修复和运动功能。在Mahmood等[47]的临床前研究中,将MSC 直接递送到大脑中的受损组织,或通过静脉内或动脉内注射间接递送,可显着改善TBI 引起的运动和认知缺陷,并且输注的MSC 能够绕过血脑屏障提高TBI 后大脑中神经保护因子的表达。Munoz 等[48]的研究表明,将MSCs 直接植入海马体后,MSCs 促进了内源性NSC的增殖、迁移和分化。此外,MSC 的分泌组可以通过降低脑组织中细胞因子的表达来调节TBI 后的炎症反应。Zhang 等[49]在TBI 患者的颅骨修复手术期间将MSCs 直接移植到受伤脑组织中未显示出不良反应,表明MSCs 在临床应用中治疗TBI 的安全性。
2.2 脊髓损伤
脊髓损伤(spinal cord injury,SCI)可破坏神经脉冲传导信号,导致脊髓的正常功能发生暂时或永久的变化,是最致残的神经系统疾病之一[50]。目前,SCI 的治疗策略主要是促进大脑和脊髓受损神经元的神经再生及突触重塑以恢复神经功能。Zhao等[51]研究证明,移植的NSC 可以在宿主脊髓中分化为神经元,并向损伤部位迁移,促进神经纤维的修复和再生,使得脊髓神经元的凋亡显著减少。通过斜面测试、后肢运动功能评分(包括后肢运动的频率和质量)以及运用Mp150 数据采集分析系统评估运动神经传导等显示,NSC 移植可显著改善脊髓横断模型大鼠的运动功能。
Lu 等[52]的一项研究显示,NSC 移植能够特异性支持脊髓损伤后的轴突再生。这项研究中,用特异性ELISA 检测发现,移植前体外培养的NSC 能够分泌大量神经营养因子,移植2 周后在SCI 模型小鼠中观察到,NSC 在脊髓损伤部位容易存活,有良好的血管形成,并能短距离迁移,最主要的是宿主中对NSC 所分泌的神经营养因子敏感的轴突细胞得到显著生长。
2.3 中风
中风(stroke)是一种急性脑血管疾病,包括缺血性和出血性中风。已有报道发现,人类iPSCs 或ESCs 衍生的NSC 系在缺血性和出血性中风动物模型中提供神经营养因子并促进血管生成和神经生成[53]。即移植的NSC 可以通过分化为成熟的神经细胞类型并整合到宿主脑组织中充当细胞替代疗法,促进血管生成并启动神经修复机制,包括突触重组和神经发生[54]。近年来,能够产生成熟神经细胞等多种细胞类型的NSC 系得到发展,包括从胎儿组织、胚胎干细胞(ESCs) 和诱导多能干细胞(iPSCs)中分离出的NSC,都被证明能促进中风和类似神经障碍后的恢复。胎儿来源的NSC 是通过在孕后7~21 d 分离人胎儿皮层、中脑或脊髓组织而产生的。当这些细胞在有丝分裂原如表皮生长因子(EGF)和成纤维细胞生长因-2(FGF-2)中培养时,即使没有遗传修饰,也能长期扩增,同时保持其神经源性和胶质源性多分化潜能[55]。然而,也有研究表明,胎儿NSC 比其他NSC 系衰老速度更快,不利于大规模诱导分化[56]。胚胎干细胞诱导的神经干细胞(ESC-NSC)的治疗用途可能优于胎儿来源的NSC,因为理论上有无限永生的ESC-NSC 来源,可以扩大细胞制造以满足临床需求[57]。
目前为止,还没有临床试验测试ESC-NSC 用于中风,但是Daadi 等[58]在补充有EGF、bFGF 和白血病抑制因子的无血清培养基中,从hESCs 中分离出一种同质的可自我更新的hNSC 系,并在大脑中动脉闭塞(MCAO)中风模型大鼠中测试这些细胞的功效,结果显示移植的细胞存活良好,并分化成神经元、星形胶质细胞和少突胶质细胞。行为学分析表明,与移植前组和载体治疗组相比,移植NSC 显著增强了受损对侧前肢的独立使用。该研究克服了伦理和监管方面的挑战,解决了从ESCs 衍生治疗性NSC 系的问题,为未来ESC-NSC 治疗中风的临床试验铺平了道路。
2.4 阿尔茨海默病
阿尔茨海默病(Alzheimer’s disease,AD)是一种因大脑皮层和皮层下区域神经元数量显著减少导致的记忆丧失和认知障碍疾病[59]。在病理学上,AD 的特征是突触和神经元的广泛丢失、炎症以及β-淀粉样蛋白(Aβ)和过度磷酸化的tau 神经原纤维缠结[60],且Poon 等[61]在AD 患者及动物模型中观察到神经营养素表达减少和神经营养素轴突运输受损。由于神经营养因子在突触可塑性、神经元健康以及学习和记忆中发挥重要作用,因此恢复AD大脑中丢失的神经营养因子是治疗的关键所在。Marsh 等[62]的研究将正常小鼠NSC 移植到老年3xTg-AD 小鼠的海马体中观察到,NSC 移植可以改善海马突触密度,挽救海马体依赖性学习和记忆,为改善认知提供了潜在的结构基础。但是NSC 移植后淀粉样蛋白和tau 的水平没有变化,而是由于NSC 产生高水平的神经营养因子显著影响了认知和突触可塑性。
早期研究表明,神经生长因子可防止神经元死亡,改善兴奋性毒性和淀粉样蛋白毒性动物模型的记忆功能[63],用于治疗AD 脑中的神经元变性和细胞死亡。然而,神经生长因子不能穿过血脑屏障,通过外周给药无法将神经生长因子输送到大脑。最新的技术方法采用生物基因治疗方法(基因修饰细胞),神经生长因子可直接插入大脑并扩散2~5 mm[64]。Tuszynski 等[65]在8 名轻度AD 患者中进行了一项体外生物基因传递的Ⅰ期临床试验,将自体成纤维细胞经基因修饰后表达人神经生长因子植入大脑。经过22 个月的随访,没有发现副作用。最终研究显示NSC 可以进行遗传修饰产生新基因,并且在移植后具有高迁移能力,可用来代替纤维母细胞,纤维母细胞在移植后主要用于递送神经生长因子以防止基底前脑胆碱能神经元的退化[66]。
2.5 帕金森综合征
帕金森综合征(Parkinson’s disease,PD)是仅次于阿尔茨海默病的第2 种最常见的神经退行性疾病,影响全球超过600 万人,其临床表现主要是是运动功能障碍,具体特征表现为运动迟缓、肌强直、静止性震颤和姿势不稳[67]。大多数与PD 相关的神经退行性过程是由病理性α-突触核蛋白(α-syn)的积累驱动的,α-突触核蛋白是一种突触前神经元蛋白,在神经系统中聚集形成路易小体(Lewy bodies,LBs)和路易神经突(Lewy neurites,LNs)[68]。此外,在病理学方面,中脑黑质致密部(SNpc)中多巴胺能(DA)神经元的缺失以及剩余DA 细胞中称为LB 的细胞质蛋白聚集体的存在是PD 的标志[69]。因此SNpc 神经元的缺失导致纹状体多巴胺能(DA)神经元的缺乏,是PD 发病的主要原因。
DA 细胞移植被认为是最有前途的细胞替代疗法,将中脑DA 神经元移植到DA 耗竭的纹状体中可以恢复DA 神经传递以替代PD 患者丢失的神经元。最近Zhang 等[70]的研究表明神经干细胞体外分化过程中,使用二苯乙烯苷(tetrahydroxystilbene glucoside,TSG)(中药何首乌的活性成分),能够增加tuj-1 阳性神经元的数量,提高酪氨酸羟化酶(th)阳性细胞及多巴胺转运体阳性神经元的比例,而后者是成熟DA 神经元的晚期标志。此外中脑DA 神经元的纹状体移植物可以从各种来源获得,例如胎儿组织、颈动脉体细胞及未成熟的视网膜细胞[71]。而神经干细胞能够分化为支持性神经胶质亚型,如星形胶质细胞,这些亚型可以产生关键的生长因子来影响突触可塑性和神经元功能,并调节脑血管形成,可能在功能恢复中发挥重要作用,因此许多人尝试使用NSC 进行神经元置换,推测在神经元分化后细胞将与宿主神经元回路整合,以替换在PD中死亡的神经元[72]。然而,这项工作具有挑战性,且结果喜忧参半。相反,最近的研究表明[73],神经营养因子(BDNF、GDNF、IGF-1、NGF)的内源性分泌或遗传过度表达可能为基于NSC 的治疗提供一种替代且可能更有前景的方法。移植NSC 分泌的神经营养因子能够增加突触可塑性,并促进神经元存活,从而改善认知和运动表现,增强长期治疗作用。
3 总结与展望
将外源性NSC 通过不同的途径移植到ND 模型动物体内后可通过不同的机制,其发挥的作用可能有[74](见图2):(1)NSC 再分化为神经元和胶质细胞,分泌神经营养因子等,改善受损神经元的存活情况;(2)促进损伤部位的血管生成和神经再生;(3)启动小胶质细胞或周围造血细胞所介导的抗炎作用,保护新生血管;(4)促进神经纤维的修复和再生,恢复神经动作电位。
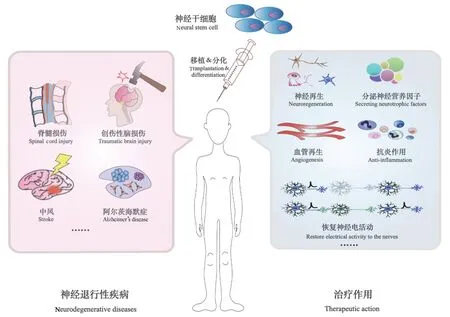
图2 神经干细胞的治疗作用Figure 2 Therapeutic effects of neural stem cells
对于大多数CNS 损伤,已经提出应用不同来源的干细胞来进行自发恢复,但大多只能产生出有限的细胞表型。最近研究建议将诱导多能干细胞(iPSCs)用于自体移植,但在应用过程中,这些基因调控的细胞由于逆转录病毒载体的不受控制整合和重组事件的发生将导致癌症形成[75],因而目前临床上用于神经性疾病治疗的最佳选择还是NSC。NSC 与受损细胞具有相同的组织来源,通常被认为是细胞介导的神经疾病治疗的最佳细胞类型[76]。
NSC 移植到哺乳动物CNS 的基础研究和临床应用已经进行了几十年,尽管神经移植物可以促进受损神经元的再生与重建,恢复神经功能缺陷,但目前要实现临床应用,仍存在一些关键障碍需要克服,其中包括[6-14]:(1)NSC 的诱导分化和增殖的机制尚不明确,在目前研究水平下尚不能精准控制NSC 的定向分化,从而影响了NSC 移植的治疗效果;(2)移植物与宿主之间的多种免疫学问题,鼠源性NSC 与人源性NSC 的种属差异以及动物内环境与人体内环境的差异导致的移植后排斥反应;(3)移植的NSC 具有成瘤的高风险以及伦理学问题等;(4)NSC 在活生物体内生存率低和治疗效果不佳。
即使有诸多障碍与困难存在,但是在干细胞治疗和研究方面,NSC 科学为人们更好理解疾病打下了基础,NSC 移植的治疗潜力已经在不同的ND 动物模型中得到证实,这将是神经退行性疾病最具前景的治疗方法。目前,NSC 治疗的潜在机制尚不清楚,还需要更多的基础研究进一步解决上述问题。但是当前大多数NSCs 研究依赖于啮齿动物模型,而人与啮齿动物的NSCs 有很大不同,许多疾病风险基因在二者之间表达存在较大差异。因此本文通过综述NSCs 的基本特性、来源及目前细胞移植的技术手段在各种神经退行性疾病中的应用进展,以期为建立人神经干细胞嵌合动物模型的成熟方法提供参考,从而在动物水平实现对人脑的模拟,更好地为神经科学研究提供动物模型和研究基础。
猜你喜欢
今日农业(2022年13期)2022-09-15 01:21:20
神经损伤与功能重建(2020年11期)2020-12-01 05:01:54
生物学通报(2020年10期)2020-08-13 08:52:26
知识经济·中国直销(2017年10期)2017-11-07 02:39:52
湖南中医药大学学报(2016年1期)2016-12-01 04:08:21
中国医药生物技术(2015年4期)2015-12-26 08:26:36
中国现代医学杂志(2015年26期)2015-12-23 11:04:22
磁共振成像(2015年1期)2015-12-23 08:52:21
西南国防医药(2015年11期)2015-02-28 19:38:46
中国卫生(2014年2期)2014-11-12 13:00:14