
烟草黑胫病植株根际土壤真菌群落多样性及结构分析
2022-05-28 09:30刘冬梅韩小女王瑞宝郭永升陈小龙王柄权程凌寒黄飞燕蔡永占刘佳妮
西南农业学报 2022年4期
方 宇,白 涛,刘冬梅,韩小女,刘 舜,王瑞宝,郭永升,陈小龙,王柄权,程凌寒,黄飞燕,余 磊,蔡永占,刘佳妮
(1.昆明学院/云南省都市特色农业工程技术研究中心,昆明 650214;2.云南省烟草公司曲靖市公司,云南 曲靖 655002;3.宣威市农业技术推广中心,云南 宣威 655499;4.河南中烟工业有限责任公司原料采购中心,郑州 450000)
【研究意义】烟草作为一种重要的经济作物在全国各省份均有所种植。然而,近年来由于土地资源匮乏,土地流转困难等原因导致烟草连作障碍问题突出,病害发生日趋严重[1]。其中,烟草黑胫病烟草整个生育期均有发生,近年来呈加重流行趋势且分布广泛,目前烟草黑胫病在中国除黑龙江省外其余各省份均有发生,且在云、贵、川等省份烟区尤为严重,发病严重地区发病率可达到75%以上,已成为制约烟草产业发展的重要影响因子[2-3]。【前人研究进展】微生物是土壤的重要组成,其在土壤中的分布特征和功能多样性在一定程度上反映了土壤质量好坏,有研究指出土壤中微生物菌群结构的失衡将会极大地增加作物感病几率[4]。植物土传病害的发生受到土壤理化性质、微生态环境等诸多因素影响,其中土壤微生物通过产生具有促进作物生长的植物生长激素和产生具有抑制病原菌生长作用的抗病活性物质在一定程度上提高植物的抗病性,因此根际土壤微生物的结构组成、多样性以及微生物营养获取方式等是植物土传病害发生的重要影响因素。Wang等[5]通过在种植苹果的土壤中添加生物炭改变了其土壤中的真菌群落结构,增加了真菌的相对丰度,同时降低了病原菌茄病镰刀菌(Fusariumsolani)在土壤中的相对丰度。童文杰等[6]发现深耕、深松措施对提高根际土壤真菌群落多样性、改善土壤真菌群落结构具有一定的促进作用,并能够抑制潜在土传病原真菌的生长,减少相关病害的发生。与此同时,根际土壤微生物群落结构也会随着植株的感病而发生改变,罗路云等[7]研究发现,南瓜白粉病病菌能够改变南瓜叶际细菌群落结构。张仁军等[8]研究发现,健康与根结线虫病田根际土壤真菌和细菌微生物群落在门和属水平上组成相似,但物种丰度存在显著差异。烟草黑胫病为真菌性土传病害,土壤是其重要的传播途径。该病原菌与众多微生物共同生存于烟株根际周围,烟株感病后势必会引起其根际微生物种群发生变化。张笑宇等[9]研究发现,烟草患黑胫病后其根际土壤微生物结构发生了改变,多样性明显降低,且相关病原菌呈现出积累趋势。向立刚等[10-11]发现,健康土壤中细菌群落的丰富度与多样性均比感青枯病和黑胫病的土壤要高。【本研究切入点】近年来,随着绿色优质烟叶生产和绿色防控体系总要求的提出,通过采用不同措施优化作物根际土壤微生物的群落分布从而达到抑制病原菌的生长已成为防治土传病害的新视角。本研究以云南烟区为背景,从烟株根际土壤微生物为出发点,利用Illumina MiSeq高通量测序技术对健康与患黑胫病植株的根际土壤真菌群落结构及多样性进行检测,并进一步分析根际土壤真菌与环境理化因子的相关性。【拟解决的关键问题】以期探明烟株感染烟草黑胫病对烟株根际土壤真菌群落的影响,揭示根际土壤真菌在黑胫病发生过程中的微生态理论机制,为从生态角度预防烟草黑胫病的发生提供一定的理论基础。
1 材料与方法
1.1 供试材料
2019年7月在云南省曲靖市宣威市热水关营试验地进行样品采集,品种为云烟100。在同一地块随机选取烟草黑胫病发病严重的烟株3株及其周边的健康烟株3株,将植株连根拔起并去除植株根系主土壤,收集根系表面2 mm根际土壤,带回实验室分装于50 mL离心管中,置于-80 ℃中冷冻保存,待用;感染黑胫病烟株根际土样编号为HJTY(XW-RS-P1、XW-RS-P2、XW-RS-P3);健康烟株根际土样编号为ZCTY(XW-RS-H1、XW-RS-H2、XW-RS-H3)。
1.2 土壤理化性质测定
土壤pH采用2.5∶1.0的水土比,用电位计法测定;有机质采用重铬酸钾—硫酸外加热法测定;碱解氮采用扩散法进行测定;有效磷采用NaHCO3浸提,钼锑抗比色法测定;速效钾采用醋酸铵浸提,火焰光度计法测定,具体试验方法依照NY/T 1377—2007、NY/T 1121.6—2006、DB51/T 1975—2014、NY/T 1121.7—2014、NY/T889—2004进行。
1.3 土壤DNA的提取与扩增
参照PowerSoil®DNA土壤基因组DNA提取试剂盒,对健康烟株和感黑胫病烟株根际土壤进行DNA的提取。以1 ng/μL DNA作为模板,选用特异性引物ITS1F和ITS2R进行PCR扩增,扩增程序为预变性,95 ℃ 3 min;变性,95 ℃ 30 s;退火,55 ℃ 30 s;延伸,72 ℃ 45 s;再延伸,72 ℃,10 min,共36个循环,扩增体系为20 μL(表1)。

表1 PCR扩增反应体系
扩增后产物进行质量检测后,按照AxyPrepDNA试剂盒方法进行纯化回收,并委托上海美吉生物进行MiSeq测序。
1.4 下机数据的质控与分析
剔除下机数据中的标签及引物序列,进行序列拼接得到原始数据;使用软件Usearch(version 7.0)进行过滤、去嵌合体序列得到有效序列;利用软件Mothur绘制稀释度曲线,判定此次测序量能否较为真实的反映样品中微生物实际情况;同时计算文库覆盖率、Chao1、ACE、Shannon、Simpson指数,对根际土壤微生物群落中的物种多样性和丰富度进行评价;在97%相似性水平上用软件Uparse进行OTUs聚类分析;利用SILVA数据库和软件RDP classifier对物种进行注释,并运用Student’s T检验对排名前10各菌属的相对丰度数据进行假设检验;使用软件Qiime进行主成分分析,并利用R语言中的Vegan软件作图进行RDA冗余分析,检测环境因子、样本、菌群三者间的关系;通过Spearman相关系数计算不同环境因子与微生物物种组成的关系,利用相关性Heatmap图评估微生物分类与环境变量之间的相关性。
1.5 数据处理
采用软件Excel 2017进行数据处理;采用SPSS 23.0数据处理系统进行统计分析。
2 结果与分析
2.1 根际土壤理化性质
患病烟株与健康烟株根际土壤理化性质差异性分析如表2所示,pH、有机质、碱解氮、有效磷和速效钾含量在两根际土壤中存在一定差异。其中pH在两者间存在显著差异,患病烟株根际土中pH较低;而有机质、碱解氮和有效磷在两处理中无显著差异,均表现出HJTY>ZCTY的规律,且HJTY较ZCTY分别增加了8.29%、18.38%和19.76%,而速效钾含量则在ZCTY中较高,相对HJTY增加了19.74%。
2.2 土壤样品测序深度验证
如图1所示,通过稀释曲线可以判断本次测序是否合理,稀释曲线虽未最终到达平台期,当测序深度达到50 000时,曲线趋于平缓,说明序列数的进一步增加不会引起较多新OTU的出现。由此可见,本次对烟株根际土壤的测序深度合理,真菌群落置信度较高,能够较为全面的反映根际土壤样品真菌群落与结构的真实情况,具有一定的研究意义,为后续的进一步分析奠定了基础。
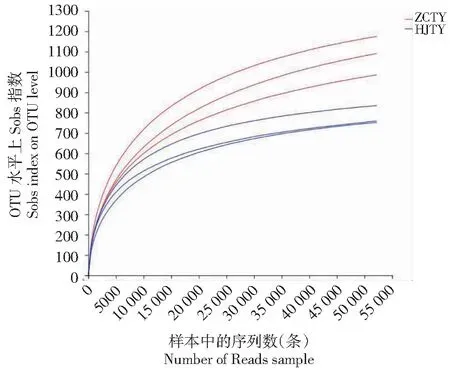
图1 健康与患病烟株根际土壤样品真菌物种丰富度稀释曲线
2.3 真菌的OTU丰度和Alpha多样性
在97%相似水平上感黑胫病与健康烟株根际土壤真菌OTU丰度和Alpha多样性如表2所示,感黑胫病和健康根际土壤真菌测序覆盖度均在99%以上,表明各土壤样品中的绝大多数序列均被检出,测序读长能够进行后续的相关分析。

表2 健康烟株与患病烟株根际土壤理化性状
由表3可知,健康根际土(ZCTY)与感黑胫病根际土(HJTY)两者的真菌OTU丰度存在明显差异,ZCTY是HJTY的1.22倍;对两根际土壤的OTU数进一步分析发现(图2),ZCTY与HJTY共有的真菌OTU 943个,从各自特有物种OTU进行分析发现,ZCTY中特有OTU 866个,占物种总量的36.93%;HJTY中特有OTU 536个,占物种总量的22.86%,ZCTY中特有OTU 是患病烟株的1.62倍,两根际土壤特有物种量存在显著差异。此外,烟株感染黑胫病后还会对其根际土壤真菌的多样性和丰富度存在一定的影响,HJTY中ACE、Chao1指数较ZCTY相比,分别降低了31.87%、30.47%,表明烟株感染黑胫病后能够降低根际土壤真菌的丰富度;Shannon、Simpson指数是反映微生物群落多样性高低的重要指数,在此次测序中发现两指数之间无显著差异,表明烟株感染黑胫病后对其根际土壤真菌的多样性影响较小。

表3 健康与患病烟株根际土壤真菌OTU丰度和Alpha多样性
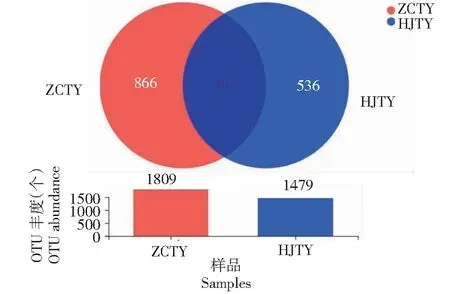
图2 根际土壤真菌OTUs韦恩图
2.4 真菌群落组成与结构分析
2.4.1 门水平真菌群落分布 如图3所示,二者根际土壤微生物区系在门水平上组成差异明显,其中ZCTY中优势菌门为子囊菌门(Ascomycota,60.08%)、油壶菌门(Olpidiomycota,16.86%)、被孢霉门(Mortierellomycota,13.03%)、担子菌门(Basidiomycota,6.05%)、壶菌门(Chytridiomycota,2.27%);HJTY中优势菌门为子囊菌门(68.89%)、担子菌门(14.37%)、被孢霉门(13.24%)、壶菌门(1.58%),其中子囊菌门、担子菌门、被孢霉门为烟株土壤的优势菌门,且其在HJTY中的相对丰度较ZCTY分别增加了14.66%、137.52%、1.61%,而壶菌门的则降低了30.39%,另外,油壶菌门为ZCTY中特有的菌门。

图3 各样品中真菌的相对丰度(门水平)
2.4.2 属水平真菌群落分布 进一步分析感黑胫病与健康根际土壤属水平上的差异(图4)发现,二者属水平的物种相对丰度差异较大。ZCTY中主要真菌有被孢霉属(Mortierella,12.41%)、油壶菌属(Olpidium,16.84%)、Soilcoccozyma(4.36%)、镰刀菌属(Fusarium,5.56%)、绿僵菌属(Metarhizium,5.59%)、毛壳菌属(Chaetomium,5.37%)、赤霉菌属(Gibberella,4.19%)、青霉属(Penicillium,1.31%)、新赤壳属(Neocosmospora,1.68%)、Plectosphaerella(2.48%)、曲霉菌属(Aspergillus,1.53%)、枝孢属(Cladosporium,1%)、篮状菌属(Talaromyces,0.54%);HJTY中优势菌属为被孢霉属(13.17%)、油壶菌属(0.07%)、Soilcoccozyma(12.08%)、镰刀菌属(8.58%)、绿僵菌属(6.53%)、毛壳菌属(5%)、赤霉菌属(2.71%)、Spiromastix(4.48%)、青霉属(3.2%)、新赤壳属(1.95%)、Papulaspora(2.8%)、Plectosphaerella(0.86%)、Aspergillus(1.63%)、外瓶柄霉属(Exophiala,1.63%)、枝孢属(1.1%)、篮状菌属(1.39%),其中HJTY中的被孢霉属、Soilcoccozyma、镰刀菌属、绿僵菌属、青霉属、新赤壳属、Aspergillus、枝孢属、篮状菌属较ZCTY均有所增加,分别增加了6.12%、177.06%、54.32%、16.82%、144.27%、16.07%、6.54%、10%、157.41%,其中Soilcoccozyma、镰刀菌属、青霉属、篮状菌属增加幅度较大,而油壶菌属、毛壳菌属、赤霉菌属、Plectosphaerella相对丰度在一定程度上有所降低,分别降低了99.58%、6.89%、35.32%、65.32%。另外Spiromastix、Papulaspora、外瓶柄霉属为HJTY特有菌属,进一步说明了烟株感染黑胫病后对其根际土壤真菌群落丰度存在一定的影响。
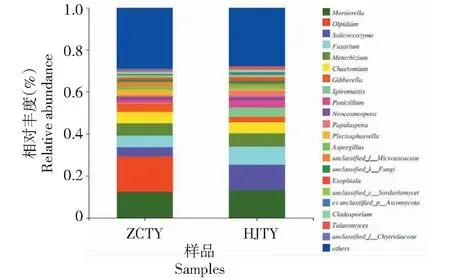
图4 各样品中真菌的相对丰度(属水平)
使用R软件,选取感黑胫病与健康烟株根际土壤真菌菌群丰度排名前30的菌属进行聚类分析,并根据聚类分析结果对分类单元进行排序绘制真菌群落分布热图。如图5所示,排名前30的菌属可聚类为两大类,而后又进一步聚类成4类,从上至下依次为菌群Ⅰ、菌群Ⅱ、菌群Ⅲ、菌群Ⅳ,30个属中有2个壶菌门、1个油壶菌门、1个被孢霉门、1个担子菌门和1个未知真菌,其余皆为子囊菌门。菌群Ⅰ在HJTY中相对丰度较高,主要聚集了Spiromastix、青霉属和Plectosphaerella;菌群Ⅱ聚类的菌属较多,其中Plectosphaerella和unclassified_f_Microascaeae在ZCTY相对丰度较高,而新赤壳属、Aspergillus、篮状菌属和外瓶柄霉属在HJTY中的相对丰度较高;菌群Ⅲ只有油壶菌属1种,主要集中分布在ZCTY中;菌群Ⅳ中主要聚集了被孢霉属、Soilcoccozyma、赤霉菌属、毛壳菌属、镰刀菌属和绿僵菌属,其中Soilcoccozyma、镰刀菌属和绿僵菌属在HJTY中的丰度较高,而赤霉菌属、毛壳菌属则在ZCTY中丰度较高。
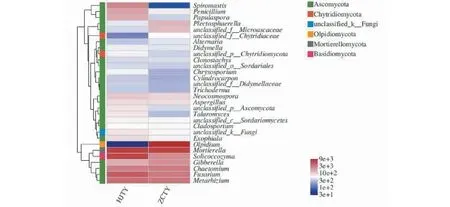
图5 属水平上根际土壤真菌群落分布热图
2.4.3 真菌物种差异分析 为找出感黑胫病与健康烟株根际土壤之间的差异真菌物种,以两根际土壤属水平上的物种注释结果及相对丰度数据为基础,选取相对丰度前10的菌属,运用Student’s T检验对其相对丰度数据进行假设检验,以此评估排名前10各菌属的相对丰度差异显著性水平。如图6所示,在排名前10的菌属中除油壶菌属、毛壳菌属和赤霉菌属外,其余菌属在感黑胫病根际土中的相对丰度均高于健康烟株根际土,且Soilcoccozyma、镰刀菌属、Spiromastix、青霉属在二者间差异显著,其中Soilcoccozyma在感黑胫病与健康烟株根际土壤之间差异最为明显,该菌在感黑胫根际土中的相对丰度是健康根际土的2.77倍,可能是影响烟草黑胫病发病的重要属;此外,镰刀菌属是植物相关病害的主要菌属,在感黑胫病根际土中的丰度显著高于健康根际土。
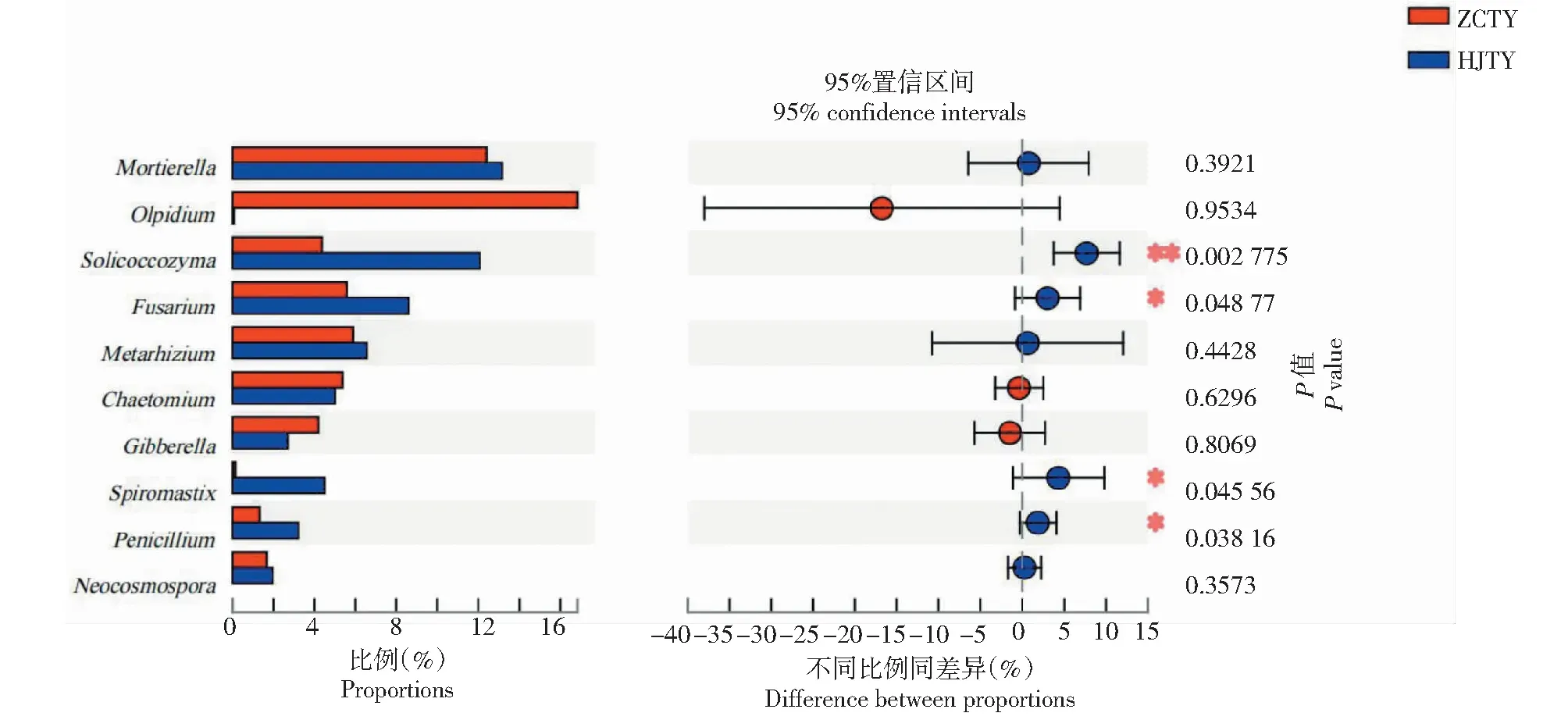
图6 属水平上真菌群落差异
2.5 真菌群落组成的层级聚类及PCoA分析
在OTU水平上对感黑胫病土壤和正常健康根际土壤真菌群落进行样本层级聚类。如图7所示,ZCTY的所有样品均能聚成1支,说明ZCTY中3个土壤样品中微生物结构相似,而HJTY中XW-RS-P2与XW-RS-P3能聚成1支,但XW-RS-P1独自聚成1支,说明其与XW-RS-P2、XW-RS-P3的微生物结构存在一定差异。

图7 OTU水平上样本层级聚类分析
另外,在OTU水平上对ZCTY和HJTY进行PCoA分析,如图8所示,PC1和PC2对样品解释度依次为49.15%和21.97%,二者合计解释度为71.12%,图中可以看出第二、三象限主要聚集了ZCTY中的真菌群落,而HJTY中的真菌群落除XW-RS-P1外均聚集在第一象限,且ZCTY中的3个样品间的距离较小,表明组间真菌群落组成差异较小;并在PC1的作用下,将ZCTY分在PC1的负值区域而将HJTY分在正值区域,进一步阐明了健康土样与患黑胫病根际土壤中真菌群落组成及分布差异显著。

图8 OTU水平样品PCoA分析
2.6 真菌群落组成与环境因子的相关性分析
将感黑胫病和健康植株根际土壤的pH、有机质、碱解氮、有效磷、速效钾作为土壤理化因子与土壤属水平上的真菌群落结构进行冗余分析,如图9所示,RDA1和RDA2对各样品的解释度分别为76.55%和12.45%,两轴共计对样品的解释度达到89%。有机质(OC)、碱解氮(N)、有效磷(EP)、pH对烟株根际土壤真菌的影响程度大于速效钾(AP),且ZCTY与有机质、碱解氮、有效磷、速效钾间的夹角均为钝角呈负相关,与pH间的夹角为锐角呈正相关,而HJTY反之。
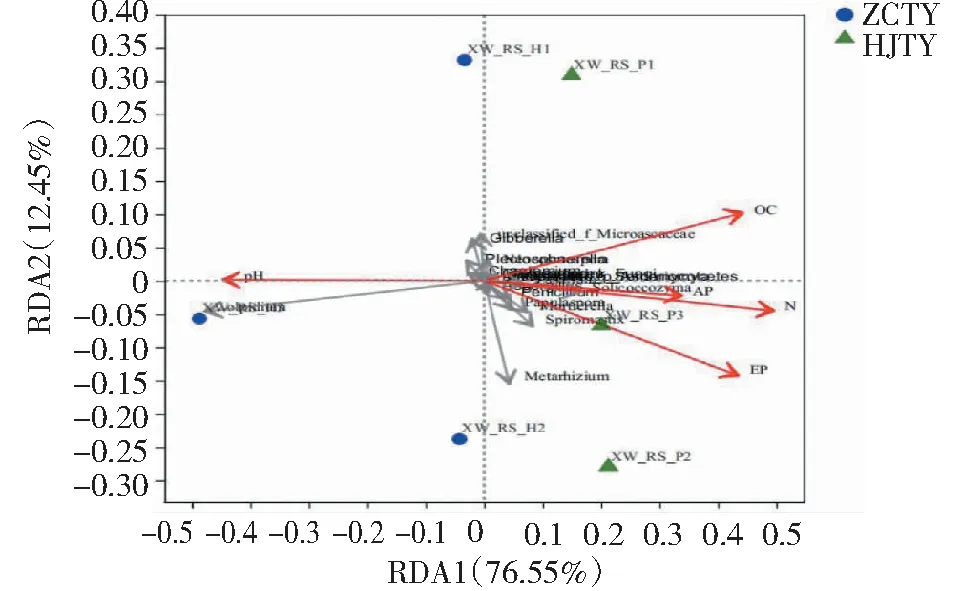
图9 不同处理根际土壤细菌组成与环境因子间的RDA冗余分析
与此同时,利用Spearman相关性系数进一步对Top 20真菌菌属进行分析并构建Spearman相关性热图,各土壤环境因子对土壤Top 20真菌菌属分布存在一定的影响。如图10 所示,油壶菌属与有机质、碱解氮均呈显著负相关,相关系数分别为-0.886、-0.943,镰刀菌属与碱解氮呈显著正相关,相关系数为0.886;Soilcoccozyma与pH呈显著负相关,而与速效钾呈显著正相关,相关系数分别为-0.841、0.829;Plectosphaerella与碱解氮、有效磷呈显著负相关,相关系数分别为-0.829、-0.943,而青霉菌属与碱解氮、有效磷呈显著正相关,相关系数为0.943、0.829;外瓶柄霉属与有机质呈显著正相关,相关性指数为0.829。

图10 属水平上不同土壤环境因子与细菌群落组成相关性分析
3 讨 论
大量研究指出,土壤质量的优劣与其微生物群落的丰富度、多样性息息相关[12],多样性、丰富度越高土壤稳定性越高,抵御病原菌侵染的能力也就越强,反之则会引起土传病害的增加[13-14]。本研究结果显示,ZCTY中ACE、Chao1和Shannon指数均比感黑胫病根际土壤要高,而Simpson指数要低于感黑胫病根际土壤,表明烟株感染黑胫病后降低了其根际土壤真菌的多样性与丰富度;此外,其真菌群落OTU数降低了18.24%,进一步说明了烟株感染黑胫病后,其根际土壤微生物数量有所降低,这与向立刚等人研究烟株感染黑胫病后其根际土壤真菌中丰富度与多样性的变化规律相一致[15]。土壤微生物的多样性对维持土壤质量和抵御病害的发生具有重要影响,多样性的降低可能是病害发生的重要因子;同时植物病害发生后随着病原菌的大量积累,有益菌群的数量逐渐降低,土壤微生物逐渐单一化,最终导致群落整体水平的降低[9, 16-17],这与本研究中感染烟草黑胫病后根际土壤真菌的种类与数量显著低于ZCTY的结论一致。
根际土壤微生物分布在植物根系的周围,其土壤微生物的群落结构与组成易受到作物种类、根系分泌物质、施肥方式、耕作方式等因素的影响[18-19]。植株感染病害后会对其根际土壤微生物及优势菌群产生一定的影响。陈巧环等[20]研究表明,菊花枯萎病导致根际土壤中大量富集了镰刀菌属致病菌和积累菊迪基氏菌属(Dickeya)、果胶杆菌属(Pectobacterium)致病细菌。伍文宪等[21]研究发现,根肿病能够引起十字花科作物根际土壤微生物中拟杆菌门的丰度显著升高,放线菌门的显著降低。本研究分析发现两根际土壤微生物的群落结构发生了明显变化,也进一步证实了烟株感染烟草黑胫病后会引起土壤微生物群落组成发生改变。在门分类水平上,子囊菌门、担子菌门、被孢霉门、壶菌门为两根际土壤中的优势菌门,前人研究表明,上述菌门是根际土壤中主要的优势菌门,广泛分布于各根际土壤中[22-24],这与本研究的结论一致。进一步分析发现,烟草黑胫侵染的烟株根际土壤中子囊菌门、被孢霉门、担子菌门的相对丰度较ZCTY分别增加了14.66%、1.61%、137.52%。子囊菌门和担子菌门分布着众多的腐生菌群,其中大部分腐生菌能够分解有机质,对土壤肥力的提高具有一定的促进作用,进而对作物的生长产生着重要的影响[25]。而被孢霉菌门中较多的属为植物病害的主要属,其丰度的增加打破了土壤微生物的平衡为黑胫病菌的侵入提供了有利条件,增加了植株患病的几率[26]。属水平上,感染黑胫病后,被孢霉属、Soilcoccozyma、镰刀菌属、绿僵菌属、青霉属、新赤壳属、Aspergillus、枝孢属、篮状菌属相对丰度有所增加,此外Student’s T检验结果表明,Soilcoccozyma、镰刀菌属、Spiromastix、青霉属在健康与感黑胫病烟株二者间具有显著差异,而Soilcoccozyma差异极显著(P<0.01),其相对丰度是ZCTY中2.77倍,可能是引起烟株患黑胫病的重要微生物因素;而镰刀菌为自然界中常见的病原菌与作物的生长密切相关[27]。其中,尖孢镰刀菌(F.oxysporium)和茄病镰刀菌侵染烟株能引起烟草根腐病发生[28],本研究中发现感染黑胫病后其根际土壤中镰刀菌丰度大幅升高,可能是由于田间根腐病与黑胫病混合发生造成该菌相对丰度的上升,同时感病植株凋萎死亡腐烂也会产生较多的腐生型镰刀菌;此外,青霉属是土壤真菌群落的主要菌群,广泛存在于根际土壤中,能够在干旱、贫瘠的土地中,且多数青霉菌为有益真菌,分泌活性物质,降低土壤中有毒物质的危害,促进作物生长,本研究该菌在感黑胫根际土壤中丰度大幅升高,可能是患病烟株为了防止黑胫病菌的进一步侵染其根际微生态做出的防御响应[29]。而油壶菌属、毛壳菌属、赤霉菌属、Plectosphaerella在ZCTY中的相对丰度明显高于HJTY,其中油壶菌属是一种重要的病毒病传播介体,在烟草上能够传播烟草坏死病毒、矮化病毒和坏死卫星病毒等病毒,其丰度的增高可能与周围患病毒病的烟株有关;而毛壳菌属对致病疫霉、辣椒疫霉、尖孢镰刀等病原菌具有较好的抑制效果[30],本研究在ZCTY中发现该菌丰度显著高于HJTY,可能是由于其对烟草疫霉具有一定的抑制作用,进而抑制了烟草黑胫病的发生。
土传病害的病原菌主要存活于土壤中,并从土壤环境获取营养物质满足生长繁殖需要,因此土壤中环境理化因子含量的变化对植物病害的发生以及土壤微生物群落的分布产生重要的影响[14]。本研究中健康烟株与患病烟株两者间的环境理化因子含量存在一定差异,且根际土壤中的主要菌属与环境理化因子均有一定的正负相关性。其中,患病土壤中的有机质和碱解氮和有效磷的含量高于健康土壤。有研究指出在土壤养分充足的环境下,病原菌更容易获取营养物质进行生长繁殖,并以此对抗具有拮抗作用的根际微生物,加剧病原菌的侵染进程[31]。而有机质含量在20~40 g/kg时有利于烤烟的生长和品质的形成,陈海念[32]在探究患青枯病和健康烟株土壤理化性质时发现土壤中有机质含量偏高可能会加剧青枯病病情的发展,这与本研究结果类似。而患病土壤中pH和速效钾的含量则低于健康土壤,土壤中的钾能够诱导植株根系分泌抑制病原菌生长的分泌物增加植株抗病性,减少病害的发生[33-34]。其次,适宜的土壤酸碱度有利于植株根系养分的吸收和抗逆性的强化,作物长期生长在酸性的土壤中会加剧病害的发生[35],而本研究中患病土壤的pH显著低于健康土壤,在一定程度上表明烟株患病后土壤酸性增加,在农业实践中可通过移栽前进行土地翻耕、合理施肥和使用生石灰等土壤改良剂进行改善土壤质量。
4 结 论
本研究表明,烟株根际土壤真菌群落结构的改变是烟草黑胫病感染烟株后造成根际土壤微生态环境变化的重要指征之一,镰刀菌属、被孢霉属、青霉属等相关病原菌及腐生菌在患黑胫病土壤中显著增加,而毛壳菌属等有益菌属显著减少;此外,健康烟株与患病烟株两者间的环境理化因子含量存在一定差异,且根际土壤中的主要菌属与环境理化因子具有一定的正负相关性。
猜你喜欢
科学技术与工程(2022年26期)2022-11-01
南方农业学报(2022年6期)2022-09-05
农业工程学报(2022年8期)2022-08-08
中国土壤与肥料(2022年4期)2022-06-14
中国农学通报(2022年14期)2022-06-01
草地学报(2022年4期)2022-04-25
土壤学报(2022年1期)2022-03-08
干旱地区农业研究(2022年1期)2022-01-28
湖北农业科学(2019年22期)2019-12-23
安徽农业科学(2019年19期)2019-12-14
