
玉米谷氨酰胺酶基因在毕赤酵母中表达条件的优化
2022-05-25 07:20韩秀娥秦兰霞任浩威
中国食品学报 2022年4期
韩秀娥,秦兰霞,邵 红,任浩威
(东北农业大学食品学院 哈尔滨 150030)
谷氨酰胺转胺酶(Transglutaminase.EC2.3.2.13,TGase),又称转谷氨酰胺酶,广泛存在于哺乳动物、鱼类、植物、细菌、真菌和绿藻等生物中[1-9],具有催化蛋白质分子内和分子间交联,蛋白质和氨基酸间连接,以及蛋白质分子内谷氨酰胺酰胺基的水解功能。通过TGase 催化反应能提高蛋白质的弹性、发泡性、溶解性、乳化性、持水性和热稳定性,从而改善蛋白质功能特性,在食品工业中具有很好的应用前景[9-19]。
国内使用的产谷氨酰胺转胺酶菌株的产酶量低,酶活低。为解决目前谷氨酰胺转胺酶的生产成本过高,产量不能满足市场需求等问题,需要获得一株高产谷氨酰胺转胺酶的菌株。随着基因工程的飞速发展,利用基因工程手段生产酶制剂和生物制药得到广泛应用[20-25]。采用基因工程的方法获得具有自主知识产权的产谷氨酰胺转胺酶菌株,是解决目前问题的一个重要手段。
毕赤酵母是一种单细胞的真核生物,在小量培养基和大规模生物反应器中都能快速生长。同其它表达系统一样,在实际应用和生产中,毕赤酵母高效生产外源蛋白受许多因素的影响[26-30]。本课题的主要目的是将前期构建的重组表达质粒pPIC9K-TGZ,经Sac I 线性化处理,电转入巴斯德毕赤酵母GS115,优选出最佳表达条件,为下游蛋白提纯及酶的生物功能特性研究奠定基础。
1 材料与方法
1.1 菌种与试剂
巴斯德毕赤酵母 (Pichia pastoris)GS115、质粒酵母表达载体pPIC9K-TGZ,东北农业大学乳品重点实验室。各种限制性内切酶,TaKaRa 公司;酵母基因组DNA 提取试剂盒,TIANGEN。
异丙基硫代半乳糖苷(IPTG),上海华舜生物工程有限公司;胰蛋白胨、酵母提取物,OXOID 公司;丙烯酰胺、N,N′-亚甲基双丙烯酰胺、十二烷基磺酸钠(SDS)、二硫苏糖醇(DTT)、Tris-Base、四甲基已二铵(TEMED)、过硫酸铵(APS)、甘氨酸、酵母氮源培养基(YNB),GIBCO 公司;蛋白分子量标准,Fermentas;TritonX-100、EB (溴化乙锭),美国Sigma 公司。
1.2 方法
1.2.1 毕赤酵母的转化及筛选 毕赤酵母转化采用电转化法。重组质粒pPIC9K-TGZ 经Sac I 线性化处理电转化至感受态细胞毕赤酵母GS115。以空质粒pPIC9K 作为阴性对照。电转化参数为:电压2 000 V,脉冲5 ms,温度0 ℃。电击结束后,立即加入1 mL 预冷的1 mol/L 山梨醇。将混合物转移到灭菌离心管中,30 ℃静置培养1.5 h,取转化物均匀涂布于含100 μg/mL 博来霉素的酵母浸出粉胨葡萄糖选择平板上。
1.2.2 酵母重组子的聚合酶链式反应(Polymerase chain reaction,PCR)鉴定 以重组酵母菌GS115/pPIC9K-TGZ DNA 作为模板,以α-因子引物:5′-TACTATTGCCAGCATTGCTGC-3′;3′AOX1 基因家族:5′-GCAAATGGCATTCTGACATCC-3′作 为引物进行PCR 鉴定。挑取一个空白表达载体pPIC9K 转化的重组菌GS115/pPIC9K 作为对照,进行相同处理。PCR 反应程序为:94 ℃预变性1 min;94 ℃变性1 min,56 ℃退火1 min,72 ℃延伸1 min,共30 个循环;72 ℃延伸10 min。2%琼脂糖凝胶电泳检测PCR 产物。
1.2.3 玉米TGase 基因在酵母中的表达 Mut+型胞内或分泌表达(空载体作对照):(1)接种一单菌落到25 mL 的BMGY 培养基中,28~30 ℃培养(250~300 r/min),使OD600nm=2~6(约16~18 h),细胞处于对数生长期;(2)1 500×g 室温离心5 min,收集细胞,移入100 mL BMMY 培养基中,用3~4层灭菌纱布封口,置于28 ℃遥床培养;(3)每24 h向培养基中添加100%甲醇,使其添加量为0.5%(体积分数),维持诱导;(4)按下面时间间隔每隔一定时间取样1 mL 来分析表达水平,以确定最佳诱导时间。以最大转速在室温条件下离心2~3 min 收集菌体;(5) 取样时间点为(h):0,24,48,72,96;(6)对分泌型表达来说,应将上清转移至单独的管中,并将其与细胞沉淀同时贮于-80 ℃直至检测。可快速冻于液氮或干冰中。对胞内表达而言,弃上清,仅需将细胞沉淀贮于-80 ℃直至检测。沉淀中加入100 μL ddH2O,涡旋振荡混匀后加入25 μL 5×SDS 凝胶加样缓冲液,继续混合20 s。样品经沸水浴处理5 min 后进行SDS-PAGE 电泳分析。
1.2.4 重组TGase 表达条件的优化
1)最佳诱导表达时间的确定 接种一酵母工程菌GS115/pPIC9k-TGZ 单菌落到25 mL 的BMGY 培养基中,28 ℃摇床培养(250~300 r/min),使OD600nm=2~6(约16~18 h),细胞处于对数生长期,1 500×g 室温离心5 min 收集细胞,移入100 mL BMMY 培养基中,用3~4 层灭菌纱布封口,置于28 ℃摇床培养,每24 h 加入培养基体积0.5%的甲醇,维持诱导;按0,24,48,72,96,120,144,168 h 的时间间隔取样,以12 000×g 离心2~3 min 收集上清,测酶活,确定诱导表达的最佳时间。
2)菌体起始诱导密度对目的蛋白表达的影响 按(1)的方法诱导表达,当菌体生长24 h 后,离心收集细胞,按不同光密度值的菌种量转入表达培养基BMMY(pH 6.0)中培养96 h,每隔24 h加入培养基体积0.5%的甲醇,以12 000×g,离心2~3 min 收集上清,测酶活,确定菌体起始诱导密度对目的蛋白表达的影响。
3)起始pH 值对目的蛋白表达水平的影响按(1)的方法培养细胞、诱导表达,接种的表达培养基pH 值分别为5.0,5.25,5.5,5.75,6.0,6.25,6.5,6.75,7.0,确定不同起始pH 值对目的蛋白表达水平的影响。
4)不同氮源对目的蛋白表达水平的影响 按(1)的方法培养细胞、诱导表达,分别接入BMMY、BMM、BMM+AA (分别添加20 种氨基酸,20 mg/种)表达培养基中培养,检测不同氮源对目的蛋白表达水平的影响。
5)甲醇浓度对目的蛋白表达的影响 按(1)的方法培养细胞、诱导表达,每隔24 h 加入甲醇使其占培养基体积分别为:0.2%,0.4%,0.6%,0.8%,1.0%,1.2%。以12 000×g,离心2~3 min 收集上清,测酶活,确定不同甲醇浓度对目的蛋白表达的影响。
6)诱导温度对目的蛋白表达量的影响 按(1)的方法培养细胞、诱导表达,诱导温度分别从15~30 ℃,确定表达量。
7)甲醇流加方式对目的蛋白表达量的影响按(1)的方法培养细胞、诱导表达,分别设定每隔8,12,16,20,24 h 流加甲醇,至其占培养基体积的0.6%,24 ℃诱导,确定添加甲醇的最佳时间。
8)诱导后阶段添加适量甘油对表达的影响按(1)的方法培养细胞诱导表达,在诱导的48 h开始每24 h 添加甘油,甘油的添加量分别为0,0.5,1,2,3,4,5 g/L,确定甘油的最佳添加量。
9)酵母工程菌表达条件的正交优化 将甲醇添加量、诱导时间、菌体起始密度及起始pH 值按照4 因素5 水平L25(54)设计正交试验,因素水平设计见表1。
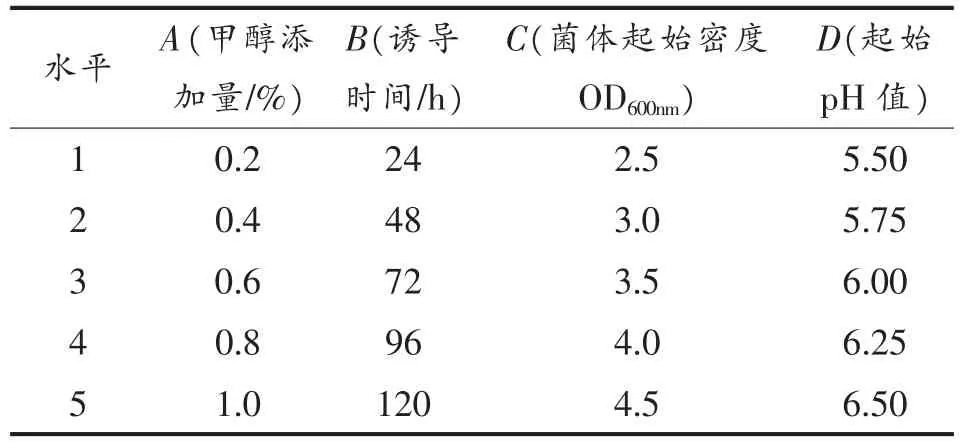
表1 毕赤酵母基因工程菌表达条件正交设计Table 1 The orthogonal design of expression condition of Pichi yeast
2 结果与分析
2.1 重组酵母菌株的PCR 鉴定
由图1可以看出,利用TGase 特异性引物,从GS115/pPIC9K-TGase 转化酵母菌的基因组中PCR 得到目的片段,而从pPIC9K 空载体转化酵母菌基因组中,并没有扩增到相应的目的带,说明目的基因已很好的整合到酵母染色体中。
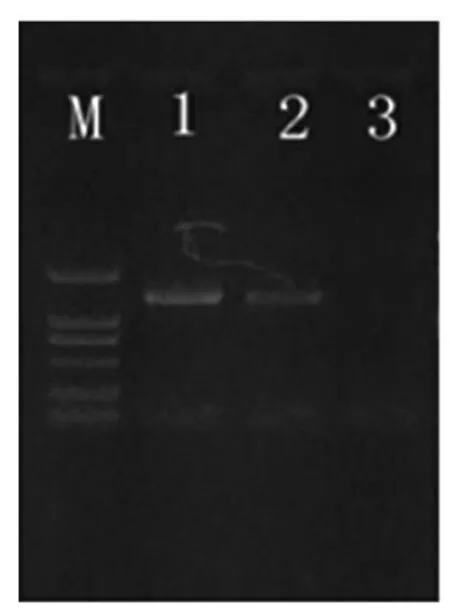
图1 重组菌株的PCR 鉴定Fig.1 Identification of GS115/pPIC9k-TGZ by PCR
2.2 重组蛋白的表达
挑取pPIC9K-TGZ 酵母单菌落,用1%甲醇诱导表达,以转化pPIC9K 空质粒的菌株同样条件培养作对照,诱导96 h 后取上清进行SDS-PAGE 电泳,与对照组比较,重组菌诱导表达上清液在分子质量约60 ku 处有明显特异性条带出现(图2箭头所示),与预期的TGase 分子质量大小相符,表明TGase 蛋白在酵母中得到了表达。通过对培养基中总蛋白浓度的测定以及对SDS-PAGE 电泳薄层扫描,可知表达蛋白的量占培养基上清液中可溶性蛋白的2%,培养基中的目的蛋白的浓度为85 mg/L,酶活为6.5 U/mL。
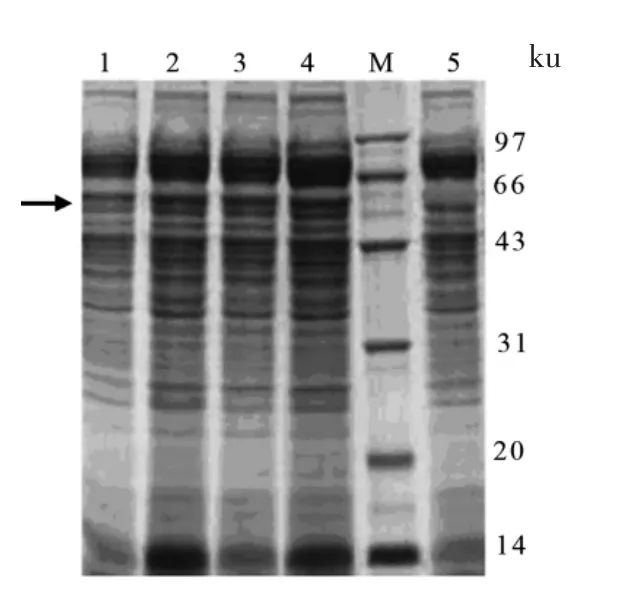
图2 酵母转化子表达上清SDS-PAGE 及Western blotting 检测Fig.2 SDS-PAGE of TGase expression supernatant and Western blotting
2.3 重组蛋白表达条件的优化
2.3.1 最佳诱导表达时间的确定 从图3可见,TGase 酵母工程菌经甲醇诱导表达后,表达上清的酶活逐渐上升至96 h 达到最高,在培养96 h后,目的蛋白的含量略有上升,而幅度很小。确定96 h 为最佳诱导表达时间。
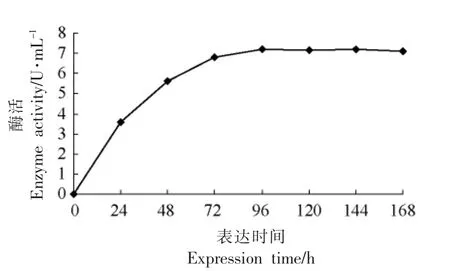
图3 表达时间对产酶的影响Fig.3 The effect of enzyme amount at different time
2.3.2 菌体起始诱导密度对目的蛋白表达的影响 由图4可见,从表达开始至发酵结束时,细胞光密度OD600nm值随着接种量的增加而呈上升趋势。当OD600nm值在2~4 范围内,随着诱导起始菌体浓度的增加,目的蛋白表达量逐渐增加,而OD600nm值超过4 后,蛋白表达量开始下降。
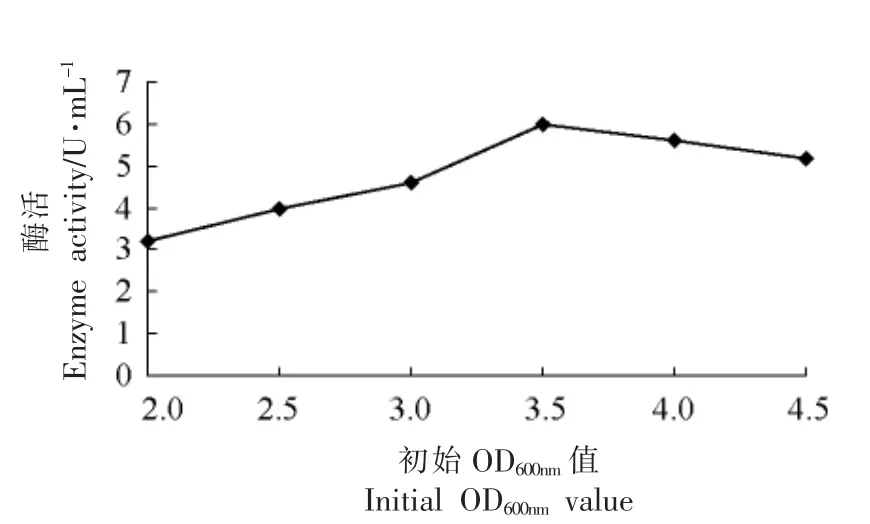
图4 诱导初始菌体浓度对产酶的影响Fig.4 The effect of enzyme with different primary yeast concentration
2.3.3 不同起始pH 值对目的蛋白表达水平的影响 选择一个合适的pH 值,使目的蛋白的表达量最高。由图5可见,pH 值为6.0 时,酶活最高,表明蛋白表达量最高。
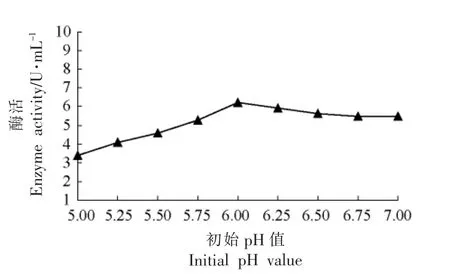
图5 初始pH 值对产酶的影响Fig.5 The effect of enzyme with different primary pH value
2.3.4 不同氮源对目的蛋白表达水平的影响 由图6可见,选择的3 种氮源,其中BMM+AA 对基因工程菌表达TGase 表达量最大。而三者差异很小,为节约成本,选用BMMY 培养基。
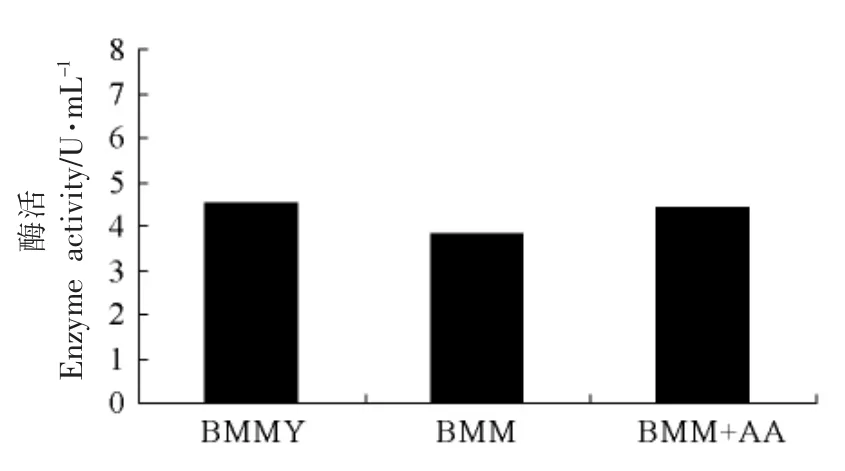
图6 不同氮源对产酶的影响Fig.6 The effect of enzyme with different nitrogen sources
2.3.5 甲醇添加量对目的蛋白表达的影响 如果甲醇浓度过低,不足以完全启动AOX1 的转录,当甲醇浓度过高,会抑制AOX1 的转录水平,甲醇主要作为表达阶段菌体生长的唯一碳源,浓度过高会造成毒性,抑制细胞生长和外源蛋白的表达,由图7可见,甲醇添加量为0.6%(体积分数)时酶活最高。
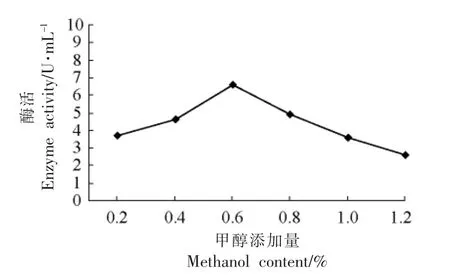
图7 不同甲醇添加量对产酶的影响Fig.7 The effect of enzyme with different methanol content
2.3.6 诱导温度对目的蛋白表达量的影响 由图8可见,低温可以降低目标蛋白的降解,提高表达量,而低温条件需要消耗更多的能量,进而提高成本,因此确定诱导的最佳温度为24 ℃。
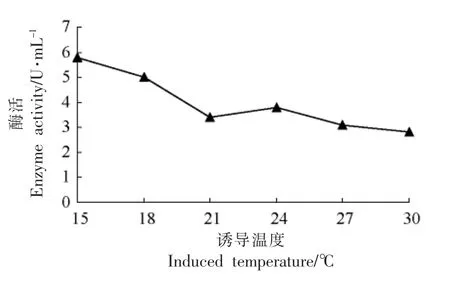
图8 不同诱导温度对产酶蛋白的影响Fig.8 The effect of enzyme with different induced temperature
2.3.7 甲醇流加方式对目的蛋白表达量的影响 由图9可见,每12 h 添加培养基体积分数0.6%的甲醇,TGase 的表达量最大。
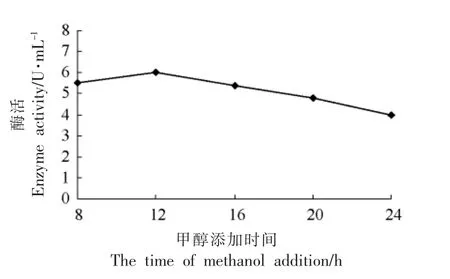
图9 不同甲醇添加时间间隔对表达目的蛋白的影响Fig.9 The effect of expression protein with different time adding methanol
2.3.8 在诱导后阶段甘油添加量对表达的影响由图10可见,在诱导48 h 后,每隔24 h 添加甘油,甘油质量浓度过高对表达有抑制作用,可知甘油添加量为1 g/L 最佳。
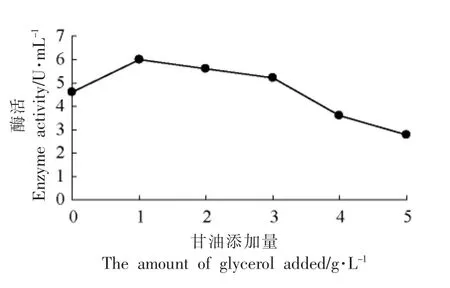
图10 诱导后阶段甘油添加量对产酶的影响Fig.10 Effect of glycerin addition on enzyme production in post-induction stage
2.3.9 工程菌表达条件的正交优化 从表2结果的极差分析可知,对毕赤酵母基因工程菌表达条件的影响因素由强至弱依次为:诱导时间>初始pH 值>甲醇添加量>菌体起始密度。从方差分析结果可知,诱导时间、甲醇添加量均达到显著差异水平(α=0.1),而菌体起始密度、初始pH 值则为差异不显著。最终确定毕赤酵母基因工程菌摇瓶最佳条件为A3B4C4D3,即诱导时间96 h,初始pH 6.0,甲醇添加量为0.6%(体积分数),菌体起始密度OD600nm=4.0。在此最优条件下测得酶活为7.51 U/mL。
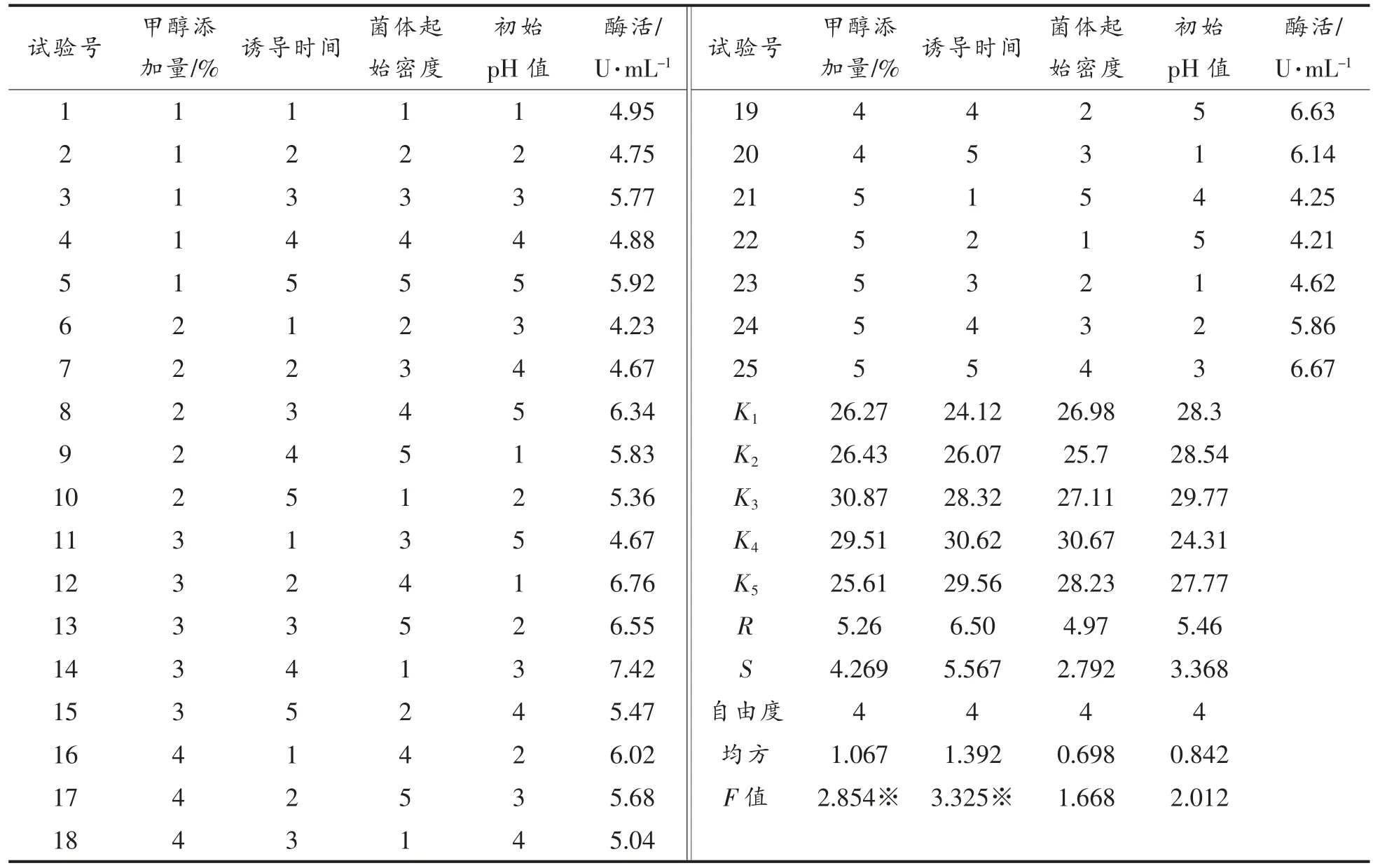
表2 毕赤酵母基因工程菌表达条件正交优化试验结果Table 2 Experimental results of orthogonal optimization of expression condition of Pichia pastoriss gene engineering strain
3 结论
毕赤酵母表达系统在表达异源蛋白上有诸多优点。本试验对最佳诱导表达时间、菌体起始诱导密度、不同起始pH 值、不同氮源、甲醇添加量、诱导温度、甲醇流加方式、在诱导后阶段甘油添加量表达条件进行优化。确定了工程菌最佳表达条件为:表达培养基为BMMY,诱导表达时间96 h,初始表达pH 值6.0,菌体起始诱导OD600nm值达4.0,诱导温度24 ℃,甲醇添加量为0.6%(体积分数),甲醇流加12 h/次、诱导48 h 后每24 h 添加1 g/L的甘油。采用优化条件后培养基上清的最高酶活可达7.51 U/mL。
由此可见,本研究获得了玉米谷氨酰胺转氨酶的最佳表达条件,为下游蛋白提纯及酶生物功能特性研究奠定基础。本研究为新型食品的应用开发奠定了理论和实践基础,也为下一步利用计算机模拟技术找出酶的活性中心,利用蛋白质工程技术对酶进行修饰,以提高酶的催化效率,提高酶的稳定性,以及酶分子改造奠定基础,从而建立模型,为酶制品的生产和应用提供思路。
猜你喜欢
现代妇产科进展(2022年9期)2022-10-07
中国调味品(2022年9期)2022-08-30
当代水产(2022年1期)2022-04-26
天津医科大学学报(2021年2期)2021-03-29
岭南现代临床外科(2020年5期)2020-12-13
食品界(2019年2期)2019-03-10
名人传记·财富人物(2017年9期)2017-11-02
名人传记·财富人物(2017年9期)2017-11-02
Coco薇(2016年8期)2016-10-09
医学研究杂志(2015年12期)2015-06-10
