
元江普通野生稻后代镉分布特点及镉积累动态变化规律
2022-05-24 03:28曾民陈佳李娥贤殷富有王玲仙曾黎琼郭蓉
生态环境学报 2022年3期
曾民,陈佳,李娥贤,殷富有,王玲仙,曾黎琼,郭蓉
1. 云南农业科学院生物技术与种质资源研究所/云南省农业生物技术重点实验室/农业部西南作物基因资源与种质创制重点实验室,云南 昆明 650223;
2. 云南农业科学院经济作物研究所,云南 昆明 650205
水稻是中国种植面积最大、食用人口最多的粮食作物,水稻易吸收并在籽粒内富集镉(Cd),这已成为中国人体镉摄入的主要途径(文典等,2021)。在中国中南及西南部分地区,稻米中的镉含量能高达1—2 mg·kg-1,远高于食品安全国家标准0.2 mg·kg-1(GB 2762—2012)(甄燕红等,2008)。目前,中国土壤镉污染面积约为1.4×104hm2,农田土壤超标位点约为7%(文典等,2020)。如何保证镉污染农田的水稻生产安全是急需解决的难题,而种植籽粒低积累镉的水稻品种是保障水稻食用安全最直接、最有效的方法(Zhao et al.,2015)。
水稻遗传基因不同是造成水稻品种籽粒镉含量差异的主要原因(Ueno et al.,2010;Sun et al.,2016)。水稻通过根系吸收镉离子后,装载到木质部并随蒸腾作用转运至地上部,然后再分配到茎叶中,最后通过韧皮部运输进入籽粒(贺慧等,2014)。水稻在吸收运输镉的同时,还能通过各组织器官内的物质与镉结合,例如纤维素、半纤维素、果胶、非蛋白质巯基及谷胱甘肽等,并将其大量储存在细胞壁或液泡内,降低镉活性,减少其进入籽粒的数量(严勋等,2019)。因此,水稻籽粒镉含量不仅与水稻对Cd吸收及运送到籽粒的能力相关,还与限制并降低镉活性相关(史静等,2015)。因此,通过研究低/高镉株系各器官内的镉含量、亚细胞分布及生育期内各组织中镉的积累动态变化,可以更好地了解不同水稻材料对镉的吸收、转运、积累存在差异的原因,以及如何限制镉向籽粒转运。
普通野生稻具有丰富的遗传多样性,拥有许多栽培稻消失或者不具备的优良性状(杨庆文等,2004)。元江普通野生稻根系发达、枝叶茂盛、生命力强、穗型散、种子小(黄兴奇等,2005)。这些性状表明,元江普通野生稻具有很强的根系吸收、光合代谢及物质转运能力。同时,其种子少,籽粒小,繁殖后代所消耗能量低,元江普通野生稻能储存大量物质来应对恶劣的环境,并进化出多种生物及非生物抗性来提高其自身生存能力(黄兴奇等,2005)。与现代栽培稻相比,野生稻植株体内富含更多的有益矿质元素和更少的有害重金属(徐玲玲等,2005;徐靖等,2014)。研究还发现,水稻对镉的吸收和转运是依靠其他矿质元素的离子通道,尚未发现吸收和转运镉的专用通道(刘仲齐等,2019)。因此从生物学性状的角度分析,元江普通野生稻根系应具有较强的镉吸收能力,同时也应该具有很强的抵抗镉毒害能力。元江普通野生稻渗入系是具有普通野生稻基因和性状的后代群体,是创新性水稻种质资源,是进行基因挖掘和理论研究的优良群体材料(柯学等,2018)。元江普通野生稻渗入系在籽粒镉含量方面存在极显著差异,可从中筛选出多份镉低积累株系及多份高积累株系(曾民等,2021)。目前对元江普通野生稻渗入系后代株系对镉吸收、转运和积累特征均不清楚,因此本文以元江普通野生稻渗入系群体中籽粒镉含量低的株系(低镉株系)和籽粒镉含量高的株系(高镉株系)为研究材料,分析镉在这两类株系各器官内的分配和亚细胞分布差异以及镉积累的动态变化特点,从而揭示元江普通野生稻渗入系后代中低镉株系与高镉株系间对镉的吸收和分配差异,为阐明元江普通野生稻渗入系镉积累的机理提供了理论基础。
1 材料与方法
1. 1 试验材料
以前期开展的元江普通野生稻渗入系低镉积累材料筛选试验中获得的籽粒镉含量低的株系(GJ71、GJ91)及籽粒镉含量高的株系(GJ11、GJ110)为试验材料,并选取籽粒镉含量居中的株系(GJ38、GJ114)作为对照材料。
1.2 试验方法
大田试验:试验地为个旧市倘甸乡水旱轮作农田,前茬作物为蚕豆,农田存在Cd 污染,主要是由污水灌溉导致,现周边已无污染源(曾民等,2019)。采用随机区组排列,3次重复,在水稻播种前平整土地、划定小区,对小区地块进行多次深耕深翻处理,每个小区面积30 m2(6 m×5 m),株距为15 cm,行距为20 cm。四周设置2 m的保护行,相邻小区间隔40 cm。于2020年3月下旬育秧,4月移栽,10月收获。土壤为褐色土,其基本理化性质为:pH值6.52,有机质21.97 g·kg-1,碱解氮1.24 g·kg-1,有效磷 12.2 mg·kg-1,速效钾 92.7 mg·kg-1,土壤总Cd质量分数为1.01 mg·kg-1。田间管理按当地大田常规方式进行管理。
1.3 样品采集、处理与分析
1.3.1 样品采集与处理
在不同生育期(分蘖期、孕穗期、抽穗期、灌浆期、乳熟期及完熟期)采集田间水稻样品各 10株,用不锈钢剪刀将根、茎、叶分离,先用自来水小心清洗水稻植株的根系、茎、叶,然后再用蒸馏水清洗,自然风干,粉碎过0.15 mm筛网,保存于自封袋中备用;待水稻成熟后,进行测产并采集植株样品,每个小区随机采集 10株植株样,将根、茎、叶分离,先用自来水小心清洗水稻植株的根系及其茎、叶,然后再用蒸馏水清洗,稻谷风干后用砻谷机脱壳,再用粉碎机将籽粒和谷壳粉碎,过0.15 mm筛网,保存于自封袋中备用。
1.3.2 Cd的亚细胞组分提取与消解
在分蘖期和灌浆期,分别采集田间种植的不同水稻株系植株各10 株,用去离子水清洗3次,各称取根、茎、叶鲜样2.0 g用于亚细胞组分提取(付铄岚等,2017)。提取步骤如下:将样品剪成约 2—3 mm的小段,在预冷研钵内加入液氮研磨,研磨成粉末后置4 ℃冰箱保存。采用Tris-HCl缓冲液(0.154 g·L-1DTT,0.25 mol·L-1蔗糖溶液,pH 7.4)作为亚细胞提取液,样品和提取剂以1∶10的比例充分混合,用漏斗和尼龙布过滤到离心管中,将尼龙布上的残渣(细胞壁)装入三角瓶中,滤液装入离心管,在15000 r·min-1下离心40 min,将上清液和沉淀分别倒入三角瓶中,分别为可溶性部分和细胞器部分。最后分别向所用三角瓶中加4∶1的硝酸和高氯酸的混合酸10 mL(可溶性部分和细胞器)和20 mL(细胞壁),待充分消化,消煮至澄清,蒸馏水定容后放入4 ℃冰箱保存待用。采用石墨炉原子吸收光谱仪(美国Thermo Elemental公司生产)测定Cd含量。
1.3.3 样品消解与测定
植物样品的消解与测定:称取0.2 g处理好的样品于50 mL离心管中,加入2 mL优级纯硝酸,混合均匀,室温下放置一夜后,用微波消煮炉消煮,程序是55 ℃ 10 min;75 ℃ 10 min;95 ℃30 min,采用标准样品(GBW 08502,大米粉标准物质)同样消煮。消煮后的样品用2%的硝酸溶液稀释至40 mL,过0.45 μm 滤膜,4 ℃保存待测。消解液中的重金属浓度用石墨炉原子吸收光谱仪测定。
土壤样品的消解与测定:称取0.2 g土壤样品于石英玻璃管中,加入 5 mL 王水(HCl∶HNO3=3∶1),混匀后室温下平衡过夜,再将土壤样品采用开放式消煮炉消解,消解程序是100 ℃ 1 h,120 ℃ 1 h;保持温度在140 ℃,直到溶液清澈,将消解后的样品置于通风橱挥发酸,然后用 2%的硝酸溶液定容到20 mL,过0.45 μm滤膜后备用。空白和标准样品(GBW 07405,土壤标准物质)同时消煮后用石墨炉原子吸收光谱仪测定Cd含量。
1.4 数据分析与统计
试验数据利用Microsoft Excel 2007进行均值、标准差计算和绘图,利用SPSS 19.0进行方差分析和LSD多重比较,采用Pearson相关系数法进行相关性分析,采用非参数检验中K独立样本检验不同株系差异性。
2 结果与分析
2.1 元江普通野生稻渗入系株系Cd 含量
由表1可知,不同渗入系株系各组织Cd含量为根>茎>叶>谷壳>籽粒,Cd主要集中在渗入系株系根部。高镉株系(GJ11、GJ110)根部 Cd质量分数分别为24.2、22.4 mg·kg-1,几乎是低镉株系(GJ71、GJ91)的 2倍。高镉株系籽粒和谷壳的Cd含量也显著高于低镉株系。低镉株系GJ91茎、叶中的镉质量分数分别为1.77、1.75 mg·kg-1,显著高于其他株系,表明GJ91能将地上部的镉更多的转运并储存在茎、叶内。低镉株系GJ71 与GJ91在茎、叶、谷壳中的镉含量均存在显著差异性,这表明 2份低镉株系对转运到地上部的镉的分配方式存在差异。
高镉株系总Cd积累量显著高于其他株系(见表 1),其中 GJ11 积累量最多,为 268.01 µg·kg-1,GJ71 Cd积累量最少,为93.15 µg·kg-1,两者相差2.88倍。高镉株系的籽粒和根中的Cd积累量显著高于低镉株系,其中GJ110籽粒Cd积累量最多,为 18.94 µg·kg-1,GJ71 籽粒 Cd 积累量最少,为 0.47µg·kg-1,而GJ91茎部Cd积累量最多。通常,以地上部与地下部 Cd含量比值(S∶R)表示水稻对 Cd的转运能力(李江遐等,2017),本文采用水稻籽粒与根部的镉含量的比值(G∶R)表征水稻将 Cd转运到籽粒的能力,比值越小表示积累能力越强。高镉株系(GJ11、GJ110)根/籽粒比值分别是 1∶43.21和 1∶32.94,远低于低镉株系(GJ71、GJ91)的根/籽粒的 1∶67.5和 1∶471,这表明高镉株系将镉运输到籽粒的能力远高于低镉株系,其中GJ110的运输能力最强,GJ71最弱。

表1 水稻完熟期不同器官Cd质量分数Table 1 The Cd content in different organs of six strains during maturing stage
2.2 元江普通野生稻渗入系株系籽粒Cd相关性分析
对渗入系株系籽粒镉含量与根、茎、叶、稻壳镉含量进行相关性分析(图 1),结果发现,籽粒镉含量与谷壳、根、茎中的镉含量存在正相关性,相关系数分别为0.737、0.739、0.519,其相关性顺序为谷壳>根>茎。
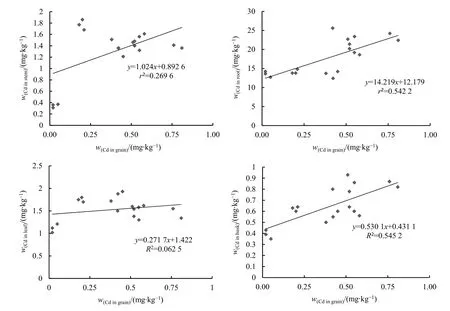
图1 籽粒镉含量与其他器官镉含量的相关性分析Figure 1 Correlation analysis of Cd content in grains and other organs
2.3 元江普通野生稻渗入系株系Cd含量动态变化规律分析
从图2可知:生育期内,各株系根部Cd含量变化趋势一致,均表现为缓慢上升,且高镉株系镉含量均高于低镉株系。各株系茎部的镉含量在抽穗期前高镉株系高于低镉株系,抽穗期后低镉株系高于高镉株系,其中 GJ71镉含量在抽穗期后快速上升,到乳熟期后又快速下降,GJ91则是在乳熟期后快速上升。从各株系叶部镉含量来看,低镉株系GJ91在乳熟期后镉含量快速上升,高于高镉株系。

图2 不同生育期水稻各器官内Cd积累动态变化Figure 2 Dynamic changes of Cd accumulation in different organs of six rice strains at different growth stages
2.4 元江普通野生稻渗入系株系Cd含量的亚细胞分布
不同株系各器官镉含量亚细胞分布(见表 2)均是细胞壁>可溶部分>细胞器,Cd在细胞壁分布最多,约占总量的39.47%—60.39%。分蘖期细胞壁Cd含量顺序为根>茎>叶,灌浆期细胞壁Cd含量为根>叶>茎。从分蘖期到灌浆期,不同株系各器官内分布在细胞壁、细胞器、可溶部分的镉含量均增加,镉含量增加量为细胞壁>可溶部分>细胞器。
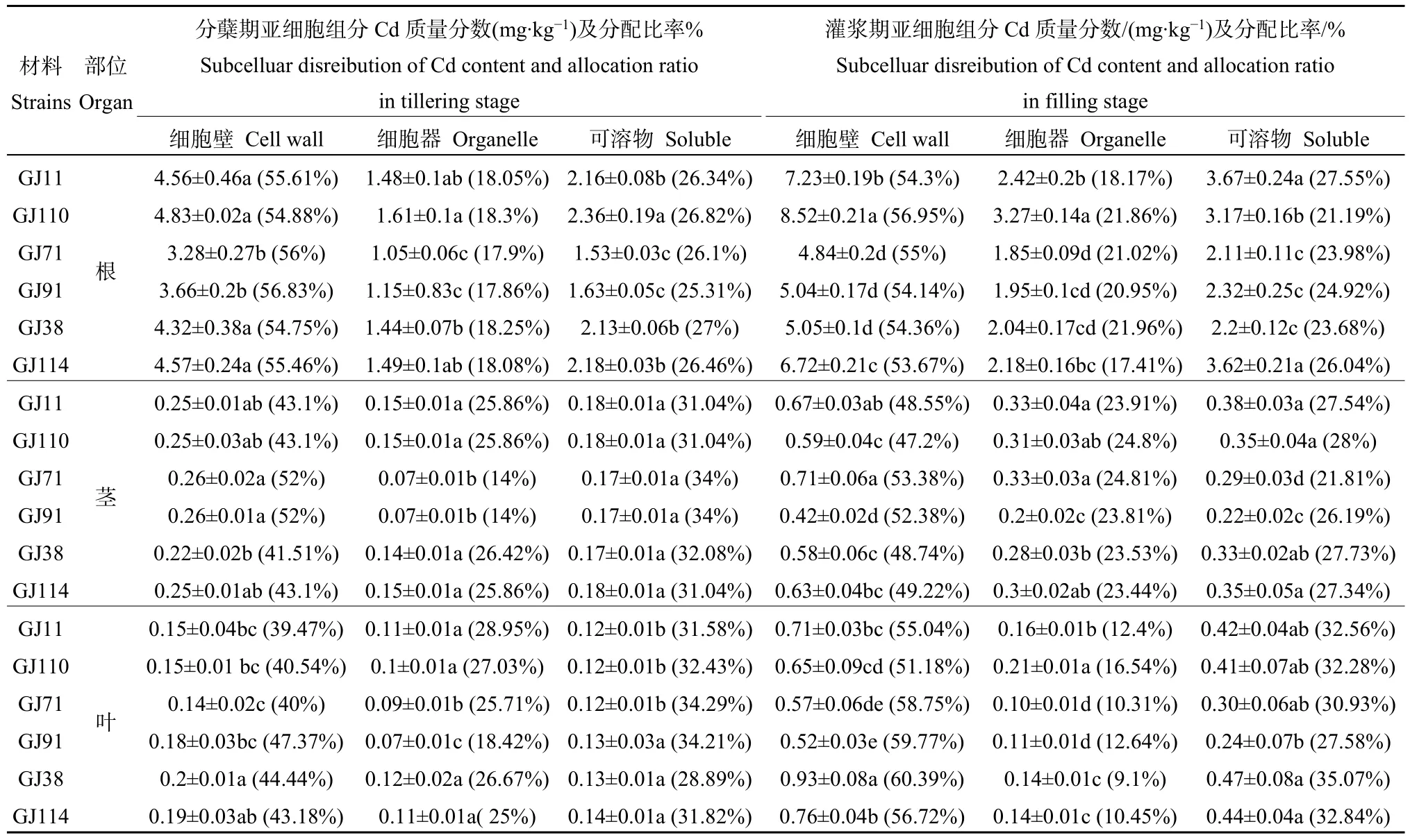
表2 渗入系水稻材料的不同时期亚细胞分布Table 2 Subcelluar disreibution of Cd content in introgression lines rice at different stage
低镉株系与高镉株系根部细胞壁、细胞器及可溶部分的Cd含量均存在显著差异。分蘖期,低镉株系与高镉株系的茎和叶部中细胞器的Cd含量存在显著差异;灌浆期,低镉株系与高镉株系的茎部可溶部分的Cd含量和叶部细胞壁的Cd含量均存在显著差异。
3 讨论
渗入系株系中存在叶部镉含量超过了茎部的现象,这与以往研究得出的水稻茎部镉含量高于叶部结论不同(李坤权等,2003),并且不同株系籽粒镉含量与叶片镉含量之间没有明显的相关性,也与以往研究得出的高积累水稻品种根系、茎和叶Cd含量均高于低积累水稻品种(付铄岚等,2017)不一致,这可能与元江普通野生稻渗入系株系保留野生稻遗传特性有关。当各渗入系株系籽粒成熟时,其植株生长依然旺盛,叶片未见枯萎,光合作用和新陈代谢旺盛,依然能将镉运输到叶片组织,所以渗入系株系叶部镉含量普遍偏高。
分蘖期后是水稻干物质积累的主要时期,灌浆期是干物质从茎叶向稻谷转移的重要时期,也是水稻籽粒蛋白质与氨基酸合成的关键时期(吴文革等,2007)。此时,茎叶中蛋白组和其他有机质将被分解成氨基酸等小分子物质,并作为合成蛋白质的原料运输到籽粒部位,而Cd可能也会随OsLCT1和OsNRAMP1等转运蛋白转运至籽粒(Uraguchi et al.,2011;Tiwari et al.,2014)。本研究中,高镉株系灌浆期后茎叶中镉含量出现下降,这可能是由于镉随氨基酸等活性物质向籽粒运输,而低镉株系茎部镉含量上升及 GJ91叶镉含量上升,可能是由于镉的积累速度大于镉向籽粒转运的速度,也可能是存在某种机制阻碍茎叶部镉向籽粒转运。
水稻中Cd主要与有活性基团的物质结合:这些物质包括细胞壁上纤维素、半纤维素、果胶、金属硫蛋白和植物螯合肽等,细胞可溶部分中的有机酸和氨基酸等,以及细胞器膜上的活性物质(Siebers et al.,2013)。研究表明,在镉胁迫下,植物首先是通过细胞壁上的纤维素、半纤维素和果胶等大分子物质与镉结合,当植物细胞壁上的重金属结合位点饱和后,镉才会与大量的有机配位体结合形成稳定态的螯合物,并被转运至液泡中储藏,从而降低细胞质中自由重金属离子的浓度(Wang et al.,2015)。本研究发现,从分蘖期到灌浆期,各器官分布在细胞壁、细胞器、可溶部分的镉含量均有所增加,由此推测镉可能是按照一定的比例分布在细胞壁、细胞器和可溶部分上,而不是与细胞壁上的大分子物质先结合至饱和后,再转运到可溶物和细胞器上。因为水稻的根系在分蘖期后已发育完全,如果根系已经饱和,那么灌浆期细胞壁上的镉含量就不应该再增加。
4 结论
(1)高镉株系对镉的吸收及转运镉到籽粒的能力均显著高于低镉株系。低镉株系GJ71与GJ91间地上部对镉的分配存在显著差异。低积累株系GJ91茎叶的镉含量显著高于其他株系,它能将更多镉分配并储存在茎叶中,从而降低籽粒镉含量。高镉株系总Cd积累量显著高于低镉株系,低镉株系GJ71总Cd积累量显著低于其他株系。
(2)低/高镉株系根部Cd 含量积累动态变化趋势一致,茎和叶的Cd含量积累的动态变化分别从孕穗期和抽穗期开始有所不同,低镉株系的镉含量在灌浆期后上升,而高镉株系的镉含量在灌浆期后下降或维持不变。
(3)低/高镉株系各组织器官镉含量的亚细胞分布为细胞壁>可溶部分>细胞器,细胞壁Cd含量约占总量的40%—60%。从分蘖期到灌浆期,各组织细胞壁、细胞器和可溶部分的镉含量均增加。
猜你喜欢
中国农业科学(2022年8期)2022-05-16
科学24小时(2021年4期)2021-03-22
科学导报(2020年70期)2020-11-09
三农资讯半月报(2020年18期)2020-10-14
数码世界(2018年1期)2018-12-23
浙江工业大学学报(2017年5期)2018-01-22
吉林农业·下半月(2017年7期)2017-07-15
农民致富之友(2017年6期)2017-05-10
饮食科学(2016年9期)2016-11-18
江苏农业科学(2014年8期)2014-10-23
