
宁南山区植被恢复模式对土壤主要酶活性、微生物多样性及土壤养分的影响
2022-05-24 07:29:32陶吉杨谭军利郑飞龙王西娜张维江
干旱地区农业研究 2022年3期
陶吉杨,谭军利,2,3,郑飞龙,王西娜,张维江,2,3
(1.宁夏大学土木与水利工程学院,宁夏 银川 750021;2.旱区现代农业水资源高效利用教育部工程研究中心,宁夏 银川 750021;3.宁夏节水灌溉与水资源调控工程技术研究中心,宁夏 银川 750021;4.宁夏大学农学院,宁夏 银川 750021)
宁南山区水土流失和土壤退化问题严重,适宜的植被恢复措施可以保持水土、改善生态环境[1],建设水源涵养林是当地生态恢复重建的重要途径之一。在生态恢复过程中,植物与土壤相互影响、互相协调,逐渐形成了生态功能侧重不同的土壤生态系统[2]。一方面植被的生长离不开土壤中的养分,另一方面不同植被类型所携带的化学成分、土壤微生物群落结构、微生物代谢活动匀不同,推动着土壤养分状况朝不同的方向发展[3]。土壤中各种生物化学反应的发生都离不开土壤水解酶,通过酶的催化作用,土壤养分有效性增强,从而更容易被植物吸收利用[4]。刘飞渡等[5]在对亚热带红壤丘陵区不同人工林型与自然恢复地的对比研究中发现,人工林土壤脲酶、酸性磷酸酶活性高于自然恢复地。此外,杨君珑等[6]通过对六盘山移民迁出区7种水源涵养林的研究发现,在不同林地中土壤酶活性也存在差异,具有乔木针阔混交的林地中磷酸酶和脲酶活性最高。土壤酶作为土壤微生物活动的产物,其差异的产生主要是由于不同植被类型下土壤微生物量、区系组成以及代谢过程改变而引起[7]。良好的土壤微生物群落结构不仅有利于土壤结构的形成与改良,还可以促进土壤有毒物质降解等,而其多样性下降对土壤的可持续利用有负面影响[8-9]。胡婵娟等[10]研究发现,刺槐作为一种固氮树种其土壤中的微生物多样性十分丰富。除此之外,随土层加深土壤微生物数量减少,在微生物活动频繁的表层土壤养分含量更为丰富[11]。不同植被恢复模式对土壤酶活性、土壤微生物与土壤养分产生了不同的影响,呈现出不同的土壤生态功能。
由于特殊的地理位置以及气候条件,宁南山区进行植被恢复的初衷是涵养更多的水源,大多数学者关注其水文方面的功能,忽视了植被恢复对土壤生态功能的潜在影响。尽管也有学者专注于森林土壤酶活性、微生物多样性以及土壤养分方面的研究,但对于宁南山区植被恢复模式对土壤性质影响的定向研究较少,不同植被恢复对土壤肥力以及土壤微生物的影响还不明确。本研究采用 Illumina-Miseq高通量测序并结合生物信息学方法,研究不同植被恢复模式下土壤主要酶(脲酶、蔗糖酶、碱性磷酸酶、过氧化氢酶)活性、细菌和真菌群落组成、土壤养分的差异,探究植被恢复对土壤性质的影响,揭示不同植被恢复模式下土壤生态功能的差异,为宁南山区通过人工造林进行生态恢复建设提供理论依据[12]。
1 材料与方法
1.1 研究区域概况
研究区域位于宁夏回族自治区固原市泾源县六盘山镇海子流域(106°14′24.4″E,35°36′31.0″N),面积2.3 km2,属于黄土高原沟壑区第二副区。地势西北高东南低,海拔高度2 186~2 835 m。多年平均气温5℃,多年平均降水量602 mm,夏季温热多雨,冬季寒冷干燥,属于温带大陆性季风气候。地形沟壑纵横,土壤以黑垆土为主,腐殖质较多,抗侵蚀能力差。自2000年退耕还林以来,植被盖度明显提高,分布有大面积的落叶松、人工油松、天然沙棘等。
1.2 采样点设置与样品采集
于2019年7月在研究区域选择7 a生油松(PinustabuliformisCarr.)、落叶松(Larixgmelinii(Rupr.) Kuzen.)、刺槐(Robiniapseadoacacia)和相同退耕年限的荒地,不同植被恢复模式样地基本情况如表1所示。在每种植被恢复模式的样地中设3个10 m×10 m的样方,每个样方中随机选取3处具有代表性的取样点,刨开表面枯落物与草本植物,用土钻分别钻取0~15 cm、15~30 cm土层根际土壤,将同一土层3个取样点的土混合均匀作为1个样品,去掉石砾与动植物残体,装入自封袋带回实验室平均分成两份,一份自然风干用于土壤养分、酶活性的测定,另一份立即过2 mm筛用于土壤微生物多样性测定,每个样品测3次。
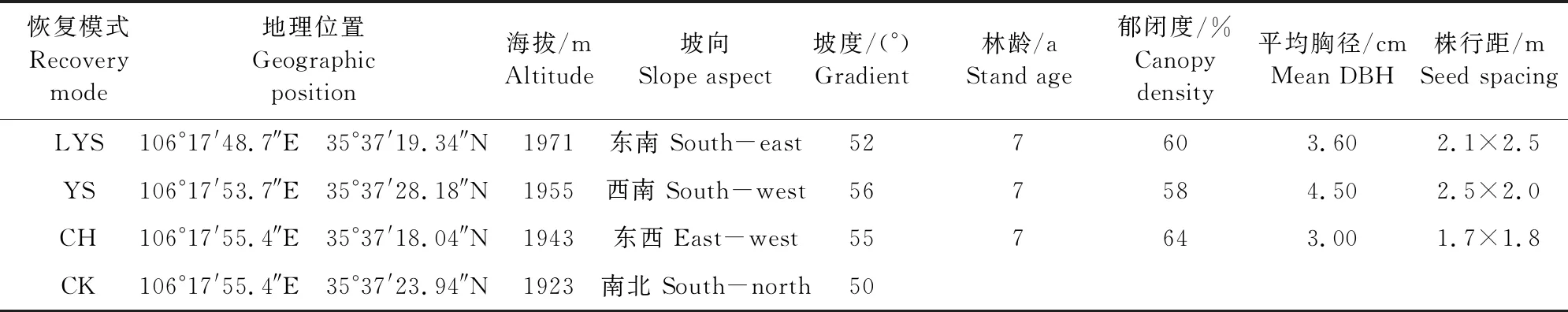
表1 不同植被恢复模式样地概况表
1.3 测定指标及方法
1.3.1 土壤养分 全氮-凯氏定氮法、全磷-钼锑比色法、碱解氮-扩散法、速效磷-NaHCO3提取法、速效钾-NH4AC-火焰光度法、有机质-K2Cr2O7容量法等具体方法参照《土壤农化分析》[13],pH采用意大利HANNA公司HI2221型pH/ORP测定仪测定,电导率采用上海仪电科学仪器股份有限公司雷磁DDS-307A型电导率仪测定。
1.3.2 土壤酶活性(在不同的培养时间、温度、基质下进行土样培养测定) 土壤脲酶-苯酚钠-次氯酸钠比色法、蔗糖酶-3,5二硝基水杨酸比色法、碱性磷酸酶-磷酸苯二钠比色法、过氧化氢酶-高锰酸钾滴定法具体方法参见《土壤与环境微生物研究法》[14]。
1.3.3 土壤微生物生物量(在真空干燥器中进行氯仿熏蒸测定) 土壤微生物生物量碳-氯仿熏蒸-FeSo4滴定法、微生物生物量氮-氯仿熏蒸-凯氏定氮法具体步骤参见《土壤与环境微生物研究法》[14]。
1.3.4 土壤微生物多样性 基于Illumina-HiSeq测序平台(北京百迈克生物科技有限公司),利用双末端测序(Paired-End)方法,构建小片段文库进行测序。根据对不同土壤样本的OUT进行分析,得到土壤微生物在界、门、纲、目、科、属、种分类水平上的分析结果以及基于Alpha多样性分析下各样品在97%相似度水平下的物种多样性指数。相关指数含义如下:
度量菌群丰度的指标:ACE和Chaol
度量菌群多样性的指标:
Shannon指数:
H=-∑PilnPi
(1)
Simpson指数:
(2)
其中,Pi为第i个分类单元中个体占微生物总个体数量的比例。
1.4 数据处理与分析
采用Excel 2010和SPSS 22.0进行数据分析,用OriginPro 8绘图。采用单因素方差分析(Duncan法)评价植被恢复模式间土壤养分、酶活性以及微生物多样性的差异(P<0.05),采用Pearson相关分析研究土壤微生物多样性与土壤养分、酶活性的相关性。
2 结果与分析
2.1 不同植被恢复模式下主要土壤酶活性
土壤酶对土壤中各种生物化学反应具有催化效应,在土壤C、N、P等养分的转化中土壤脲酶、蔗糖酶、碱性磷酸酶起着举足轻重的作用[15]。如图1所示,在0~15 cm土层落叶松与油松的脲酶、蔗糖酶、过氧化氢酶活性均无显著差异,两者的蔗糖酶活性显著低于刺槐,分别仅占刺槐的38%和42%,碱性磷酸酶和过氧化氢酶活性较高,其中油松的碱性磷酸酶活性为刺槐的4.5倍,过氧化氢酶活性为荒地的3.1倍。刺槐的酶活性最接近荒地,蔗糖酶活性及其离散程度均最高。除脲酶和蔗糖酶活性外,荒地其他两种酶活性均较低,说明在以草本为主的自然恢复模式中,草本植物的固氮作用强于木本植物,氮素周转速度更快。
注:不同字母表示不同植被恢复模式间差异显著(P<0.05),下同。
在15~30 cm土层,落叶松与油松的4种酶活性均无显著性差异,荒地的过氧化氢酶活性显著低于其他3种人工植被恢复模式。与0~15 cm土层酶活性相比,刺槐的脲酶活性显著提高了167%,油松的碱性磷酸酶活性降低了43.6%。土壤酶活性基本随土层加深而降低,其中碱性磷酸酶的变化最为显著,变化范围由原来的1.03~4.63 mg·g-1·d-1变为1.21~2.61 mg·g-1·d-1,减小了61%。
2.2 不同植被恢复模式下土壤微生物生物量碳氮与微生物多样性
微生物生物量与有机质和氮素周转密切相关,表征着土壤肥力以及人类活动对生态系统的影响,在有关土壤微环境的研究中通常作为一种生态学指标[16-17]。如表2所示,荒地的微生物生物量C最低,随土层加深,4种植被恢复模式的微生物生物量C减小,其中油松减小幅度最大为32.73%。油松的微生物生物量N最低,随土层加深,落叶松的微生物生物量N显著增高,含量达到了油松的6.16倍,而其他植被恢复模式的微生物生物量N都在减小。研究表明微生物生物量C/N比表征了单位重量土壤有机质中微生物生物量碳与氮的比值,影响着微生物的群落结构[18-19]。油松土壤微生物生物量C/N在0~30cm土层最高,说明这一恢复模式下土壤微生物生物量碳含量在微生物生物量中占比较大,微生物利用碳源的能力较强,也能更好地促进N的矿化,有利于微生物多样性发展。随土层加深,油松和刺槐的C/N比显著增大,落叶松、荒地有所减小。总的来说,人工植被恢复模式较自然恢复为微生物多样性发展创造了更有利的条件。
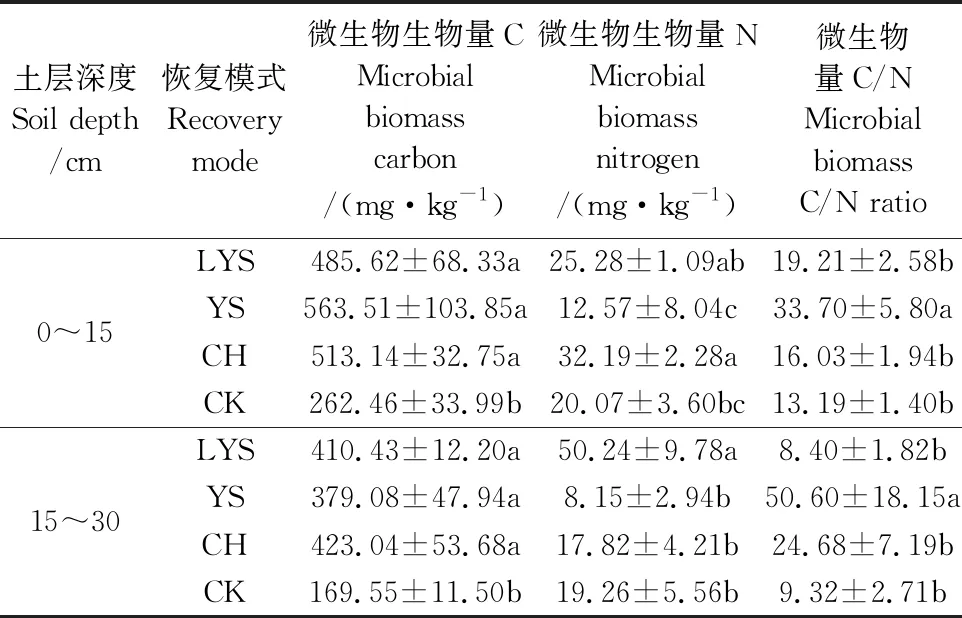
表2 不同植被恢复模式下土壤微生物生物量C、N
如表3所示,在0~15 cm土层4种植被恢复模式土壤真菌的ACE和Chaol指数均表现为荒地>油松>落叶松>刺槐,其中刺槐的上述两个指数均显著低于荒地。Simpson指数作为逆向指数,其值越高表明土壤真菌群落中某种或某几种真菌占明显优势,对资源的利用程度强于其他真菌群落。落叶松的Simpson指数显著高于刺槐和荒地,分别为刺槐和荒地的4.2倍和3.1倍,并不利于土壤真菌多样性的发展,Shannon指数最低,不到刺槐的2/3。在15~30 cm土层4种植被恢复模式间土壤真菌的ACE和Chaol指数大小规律与0~15 cm土层相同,荒地的ACE指数显著高于落叶松和刺槐,Chaol指数达刺槐的1.5倍以上,Simpson和Shannon指数在4种植被恢复模式间均未出现显著性差异。随土层加深,ACE和Chaol指数减小,其中刺槐减小幅度最大,分别为24.63%和19.2%。除落叶松外其他3种植被恢复模式的Simpson指数均随土层加深而增大,Shannon指数随土层加深而减小。
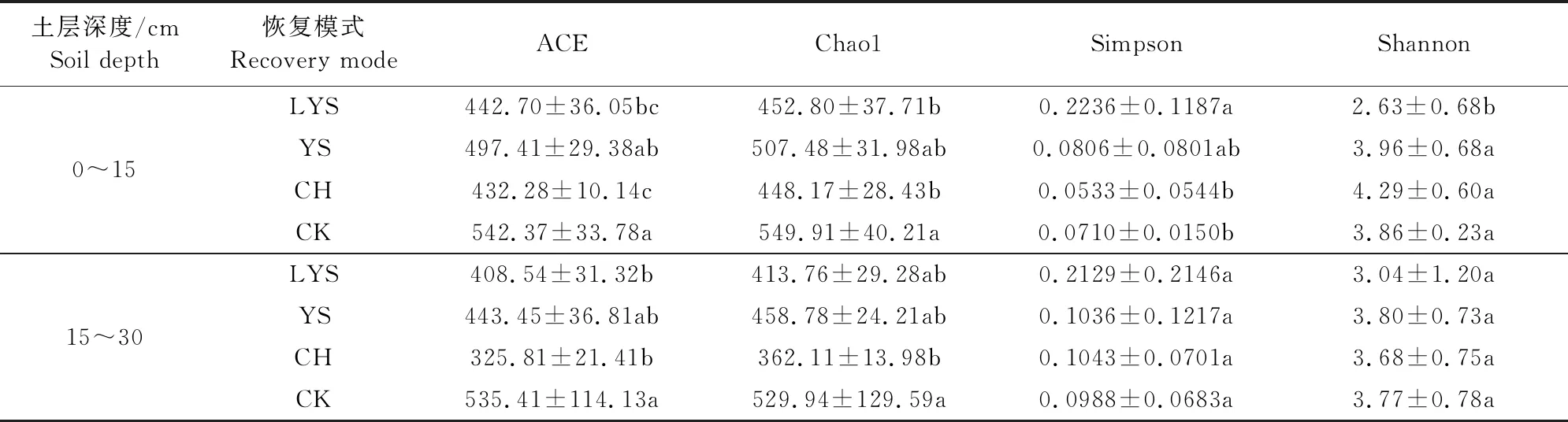
表3 不同植被恢复模式下土壤真菌多样性
如表4所示,4种植被恢复模式土壤细菌各项指数均未出现显著性差异,荒地各项指数的离散程度在两个土层中均为最大,这直接导致了荒地与其他3种人工植被恢复模式差异不显著。油松的ACE、Chaol指数随土层加深而增大,Simpson指数随土层加深而减小,呈现出与其他3种植被恢复模式不同的规律。4种植被恢复模式的Shannon指数随土层的加深减小12%左右,除Simpson指数外土壤细菌的其他3种指数均高于土壤真菌,ACE和Chaol指数更是高出2倍以上。
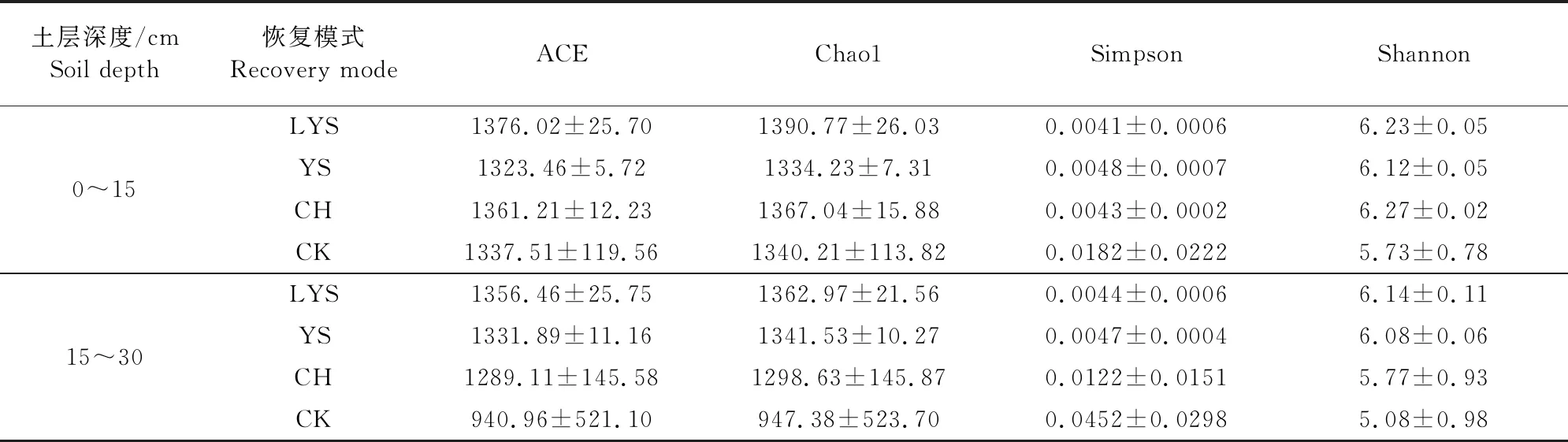
表4 不同植被恢复模式下土壤细菌多样性
为了更好地探究土壤微生物的物种多样性,使土壤养分差异的成因具体化,分别绘制4种不同植被恢复模式在真菌和细菌门水平上排名前9和前11的微生物群落。如图2所示,4种不同植被恢复模式土壤微生物群落组成随土层加深没有发生太大变化,在0~15 cm土层土壤真菌优势群落主要是子囊菌门(35.32%~60.11%)和担子菌门(15.22%~47.24%),此外还有被孢菌门(1.28%~9.84%)、球囊菌门(0.06%~0.65%)、壶菌门(0.03%~0.31%)、油壶菌门(0.08%~0.22%)、罗兹菌门(0.06%~0.14%)和游动菌门(0.01%~0.05%)。与荒地相比,刺槐的子囊菌门的相对丰度有所提高,落叶松和油松的相对丰度较荒地分别降低了39.32%和30.60%,原因可能是子囊菌大都与植物根系形成共生或与动植物残体形成腐生,刺槐和荒地中根系分布广泛且枯落物更多,更适宜子囊菌的生存与繁殖。刺槐的担子菌门相对丰度较荒地有所减小,落叶松和油松分别较荒地增长2.03倍和1.46倍,3种人工植被恢复模式的被孢菌门相对丰度较荒地都有所降低。土壤细菌中相对丰度超过10%的微生物群落主要是变形菌门(29.83%~38.42%)、酸杆菌门(17.38%~20.19%)、放线菌门(11%~12.89%)、拟杆菌门(6.57%~11.04%)和厚壁菌门(0.68%~11.04%),3种人工植被恢复模式的变形菌门和放线菌门相对丰度较荒地均有提高,拟杆菌门和厚壁菌门相对丰度较荒地分别降低34.51%和89.04%以上,酸酐菌门在4种植被恢复模式间变化不大。

图2 不同植被恢复模式下土壤微生物群落组成
15~30 cm土层土壤真菌和细菌群落组成较0~15 cm土层没有明显变化,4种植被恢复模式土壤真菌中子囊菌门和担子菌门相对丰度总占比均达到了80%以上,担子菌门与子囊菌门作为分布最为广泛的两类菌种,在资源利用上往往具有趋同性,这就导致子囊菌门相对丰度越高的植被恢复模式,其担子菌门相对丰度越低。其中,除落叶松子囊菌门相对丰度较0~15 cm土层有所降低外,其他3种植被恢复模式子囊菌门相对丰度都有所提高,油松增幅最大为26.63%,此外,4种植被恢复模式被孢菌门的变化范围较0~15 cm土层缩小了将近1/2。土壤细菌中相对丰度超过10%的微生物群落中除拟杆菌门和厚壁菌门外,其他菌门的相对丰度在3种人工植被恢复模式下较荒地均有所提高。随土层加深,4种植被恢复模式变形菌门的相对丰度降低,其中荒地变化最为显著,降低了一半以上,原因可能是随土层加深变形菌利用光合作用储能受到抑制,导致菌落减小;厚壁菌门的相对丰度提高,其中落叶松和刺槐分别达到了0~15 cm土层的9.3倍和9.1倍,此外,荒地拟杆菌门相对丰度的增长也较为显著。
2.3 不同植被恢复模式下土壤养分特征
土壤养分主要来源于土壤本身、植物凋落物以及植物根系分泌物,在不同植被恢复模式下存在显著差异[20]。如图3所示,0~15 cm土层落叶松与油松的各项养分指标均未出现显著性差异,其中落叶松的碱解氮含量最高,可达115.73 g·kg-1,为荒地的1.63倍。刺槐的速效钾和有机质含量显著高于其他3种植被恢复模式,除土壤全氮、速效钾含量外,荒地的其他养分含量均显著低于其他3种人工植被恢复模式,有机质、速效磷、全磷含量分别为刺槐的1/5、2/5、1/2。4种植被恢复模式土壤全磷和速效磷含量大小基本一致性,均为刺槐>油松>落叶松>荒地,3种人工植被恢复模式这两个指标的含量均达到了荒地的1.5倍以上。
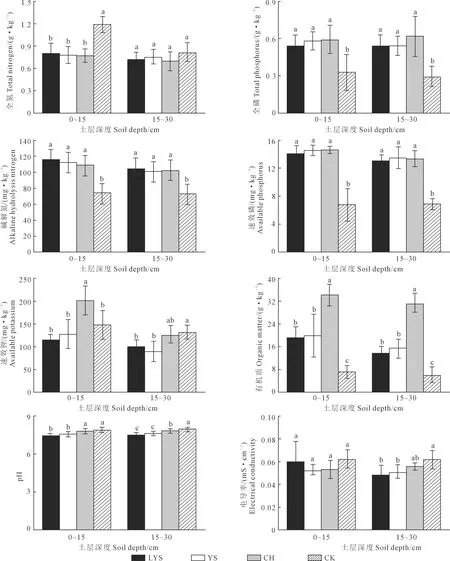
图3 不同植被恢复模式下土壤养分
随土层加深,除pH和电导率以外,4种植被恢复模式的养分含量均有减小的趋势,且差异性基本不变。其中荒地的全氮含量减小幅度最大为31.88%,其他3种人工植被恢复模式的变幅维持在11%以内,刺槐的速效钾含量减小幅度最大,超过了其他3种植被恢复模式变幅的1.25倍以上。0~15 cm和15~30 cm土层pH值的范围维持在7.5~8.0之间,土壤呈弱碱性,不同植被恢复模式间电导率的差异性随土层加深而增加。
2.4 土壤主要酶活性、微生物多样性与土壤养分相关性分析
土壤中的养分、酶活性与土壤微生物是相辅相承的关系,一方面土壤养分为土壤微生物的生长提供食物,为土壤胶结提供腐殖质改变微生物的活动场所,另一方面土壤养分的积累与消耗离不开一系列的酶促反应,而微生物则是各种水解酶的来源[21-22]。如表5所示,脲酶能够加快氮的周转,在0~15 cm土层中与全氮极显著正相关(P<0.01),与碱解氮、速效磷极显著负相关(P<0.01),过氧化氢酶同样与上述3种土壤养分极显著相关(P<0.01),且相关性与脲酶相反。蔗糖酶与土壤速效钾、pH显著正相关(P<0.05),碱性磷酸酶与土壤养分无显著相关性。真菌ACE、Chaol指数与土壤养分的相关性较Simpson、Shannon指数更强,均与碱解氮、速效磷显著负相关 (P<0.05),分别与土壤有机质极显著正相关(P<0.01)和显著正相关(P<0.05),其中ACE指数还与全氮具有显著的正相关关系(P<0.05)。说明真菌丰度与土壤养分密切相关,土壤有机质为微生物提供生存的食物,是其中极为重要的因素。土壤细菌ACE、Chaol指数与土壤养分无显著相关性,Simpson指数与土壤全氮显著正相关(P<0.05),与全磷极显著负相关(P<0.01),Shannon指数与上述两种养分的相关关系显著性与Simpson指数相同,相关关系与Simpson指数相反。
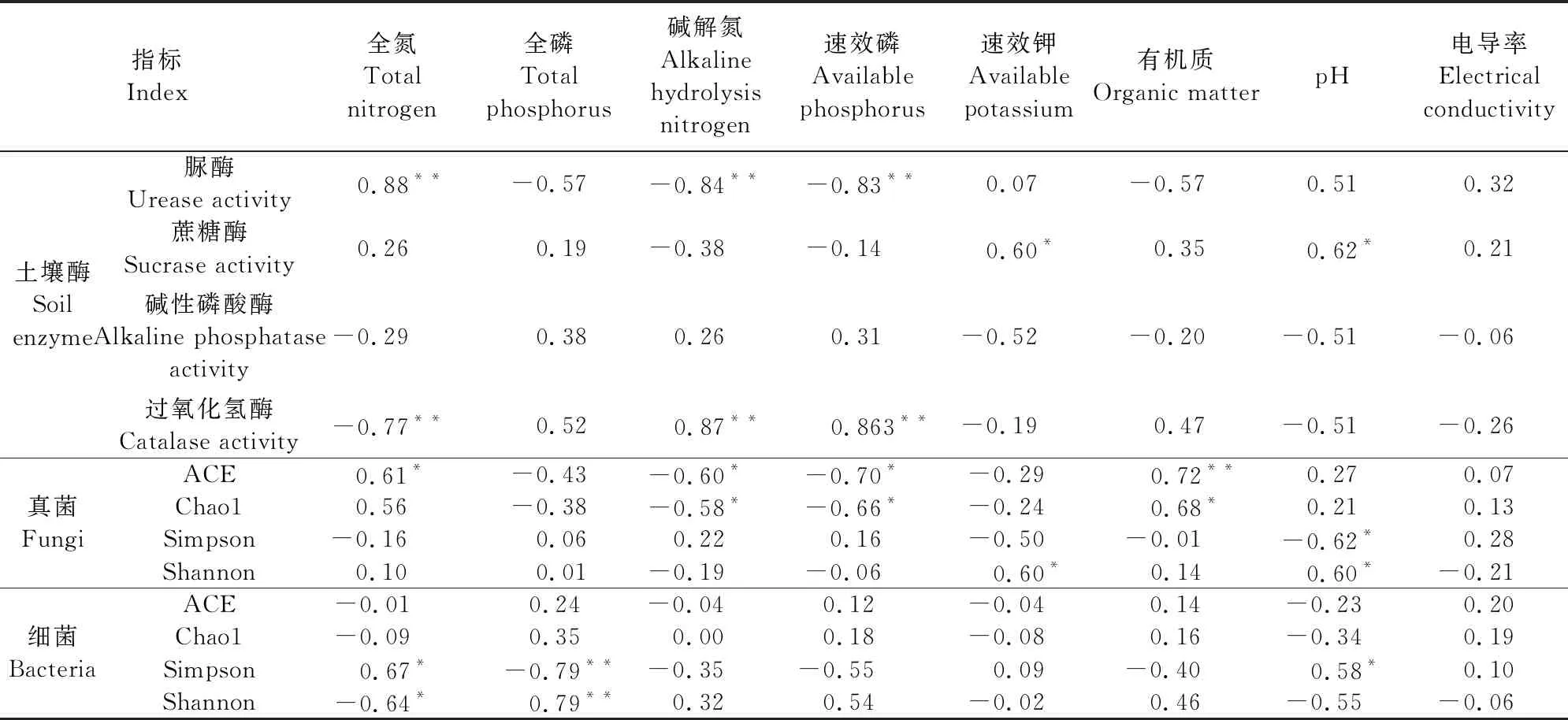
表5 0~15 cm土层土壤酶活性、微生物多样性与土壤养分相关性
如表6所示,在15~30 cm土层,土壤脲酶与速效钾、pH值极显著正相关(P<0.01),碱性磷酸酶和过氧化氢酶均与全磷、碱解氮、速效磷显著正相关(P<0.05),与速效钾、pH值显著负相关(P<0.05),其中与速效钾的负相关关系达到了极显著水平(P<0.01)。这是因为土壤钾含量过高易造成土壤板结,不利于微生物活动分泌土壤酶,且碱性条件下会抑制碱性磷酸酶与过氧化氢酶活性。土壤真菌ACE和Chaol指数与土壤养分的相关性随土层加深基本呈增强趋势,Simpson和Shannon指数与土壤养分无显著相关性。土壤细菌Simpson指数与全氮、全磷、pH值的相关性随土层加深而减弱,与碱解氮、速效磷的相关性达到了极显著负相关(P<0.01)。

表6 15~30 cm土层土壤酶活性、微生物多样性与土壤养分相关性
3 讨 论
3.1 植被恢复模式对土壤酶活性、微生物多样性的影响
植被恢复模式对土壤养分、酶活性等有着较大影响。荒地作为一种自然生态恢复方式,其土壤状况更符合自然演替规律[23-24]。土壤酶使土壤的各种养分有效化,酶活性越大催化作用越强,各种酶协同作用维护了生态系统的稳定性[25-26]。土壤中脲酶活性能够反映氮素周转速率,由于草本植物的固氮作用强于木本植物,因此荒地的脲酶活性较强[27]。凋落物化学组成和根系分泌物影响了土壤有机质的输入,蔗糖酶在植物碳水化合物代谢中发挥着重要作用[28],因此刺槐恢复模式土壤有机质含量最高,蔗糖酶活性也最强。油松作为一种油料植物,其叶片中含有大量磷素,根系分泌产生磷酸酶,故土壤中碱性磷酸酶活性最强。过氧化氢酶是一种还原酶,其稳定性较强,随土层加深酶活性变化不大,这与黄懿梅等[29]研究结果一致,但其活性的大小与土壤pH的变化呈现一定的规律,即土壤pH值越高,过氧化氢酶活性越低,这可能是过氧化氢酶活性在碱性条件下受到抑制所导致[30]。
本研究中植被恢复模式对土壤细菌多样性无显著影响,而对真菌影响较大,说明了土壤细菌对于环境的适应能力更强,土壤真菌对于植被类型的选择较细菌更为严苛。3种人工植被恢复模式并未显著提高土壤微生物的多样性,其中刺槐还降低了土壤真菌的ACE、Chaol指数,这可能是由于土壤孔隙度较小,不利于土壤微生物的活动,抑制了微生物的代谢过程和群落结构的发展。荒地土壤真菌多样性最为丰富,这可能与土壤水分有关,在适宜的水分条件下真菌种类及数量最多,且荒地的氮素储量和周转速率最快,这都为土壤真菌的生存提供了必要条件[31]。此外,土壤真菌多样性还与有机质含量密切相关,有机质含量越高可供微生物利用的碳源越多,但在本研究中有机质含量最高的刺槐土壤微生物多样性并非最大,原因可能在于人工种植刺槐对土壤中原本的微生物群落结构造成了扰动,新的微生物群落不成熟,旧的群落遭到破坏,且由于植物特性土壤中固氮微生物较多,制约了利用其他资源的微生物的发展,所以土壤微生物多样性不及荒地。土壤真菌以子囊菌门和担子菌门为主要优势菌种,子囊菌门作为全球土壤生态系统的优势类群,能够利用多种养分供其自身发展,而担子菌门的一些类群对环境扰动十分敏感,因此可以作为土壤扰动强弱的指示真菌[32]。两种针叶林的担子菌门较荒地均有所提高,说明人工植被恢复在一定程度上减小了外界环境对土壤生态系统的扰动。
3.2 植被恢复模式对土壤养分的影响
不同植被恢复模式下生物量、植被盖度、根系分泌物与枯落物的化学组成,以及根系周转速率均存在差异[33],土壤酶活性以及微生物种类不同,这些因素增强了工植被恢复对土壤养分的改善效果[34]。其中刺槐是固氮植物在改善土壤养分上表现最好,蔗糖酶活性强,加快了土壤中碳的运转,加上刺槐凋落物产量大,阔叶分解较针叶更快,有机质积累迅速,继而增加了土壤中其他元素的有效性。荒地的全氮含量最高,因为荒地草本植物居多且根系多分布在0~15 cm土层,细菌中的拟杆菌门常与根系形成共生固氮,且大量植被在一年内衰败很快归于土壤,周而复始。在荒地中土壤脲酶活性强加速了氮的转化,碱解氮常常供不应求,因此积累较少[35],且荒地没有冠层遮蔽,降雨对养分的淋溶更严重,导致荒地的养分含量较低[36]。落叶松、油松两种针叶林属于油料植物,其枯落物化学组成中磷素含量较为丰富,枯落物分解营养元素回归土壤,全磷和速效磷含量升高,再加上植被根系分泌物以及根际微生物作用,碱性磷酸酶活性强,促进了土壤磷素有效性。
4 结 论
人工植被恢复模式对于养分的改善效果较自然恢复更为显著,其中刺槐模式对提高土壤养分的作用明显。荒地虽然提高了土壤微生物多样性,但土壤养分条件较差。综合土壤养分、主要酶活性及微生物多样三个方面看,在今后营造水源涵养林时,刺槐类的固氮阔叶树种可以作为宁南山区营造水源涵养林的优先选择的树种。
猜你喜欢
当代作家(2023年3期)2023-04-23 21:26:58
现代园艺(2020年2期)2020-12-22 05:57:04
甘肃林业科技(2020年1期)2020-06-23 03:38:22
中国水土保持科学(2019年6期)2019-04-26 05:14:34
小溪流(故事作文)(2018年10期)2018-12-28 10:45:08
中国诗歌(2018年6期)2018-11-14 13:24:12
现代园艺(2018年2期)2018-03-15 08:00:12
山东林业科技(2016年5期)2016-07-05 00:43:00
湖南农业(2016年3期)2016-06-05 09:37:34
电脑迷(2015年3期)2015-04-29 21:59:57
