
冷季补饲对放牧后备母羊生产性能和血液生化指标的影响
2022-05-23 11:09:10张志军郭同军阿不夏合曼穆巴拉克古再丽努尔艾麦提秦荣艳巴音巴特王承敏王文奇
草业科学 2022年5期
张志军,郭同军,阿不夏合曼·穆巴拉克,古再丽努尔·艾麦提,秦荣艳,巴音巴特,王承敏,叶 军,王文奇
(1. 新疆畜牧科学院饲料研究所, 新疆 乌鲁木齐 830011;2. 哈密市摆盛牧业有限责任公司, 新疆 哈密 839000)
新疆是全国五大牧区之一,牧区养羊业是新疆畜牧业重要组成部分,5 725.88万hm2的天然草地面积,是放牧绵羊的主要营养来源[1]。但天然草地从丰盛到枯萎的四季更迭规律,使得牧区绵羊“夏活、秋肥、冬瘦、春死”现象屡见不鲜,直接影响牧区养殖的产出和养殖户的经济效益[2-3]。冬春两季枯草期对放牧绵羊进行补饲,调控好家畜摄入的营养水平,可实现放牧家畜营养均衡,进而降低牧民因草场营养水平差导致的养殖效益损失。作为反刍动物小肠氨基酸合成重要来源的瘤胃微生物蛋白(microbial protein, MCP),其含量与为瘤胃微生物提供的能量和蛋白水平有关,且两者的同步供给和释放很大程度上影响瘤胃微生物的生长。对放牧绵羊补饲,需综合考虑摄入的可利用有机物和瘤胃可降解氮的水平。
近年来,学者们针对牧区绵羊冬春季补饲进行的研究结果表明,为牧区羔羊补饲能氮比为73.25 MJ·kg-1的精料,羔羊生长性能最好[4]。为牧区妊娠期和泌乳期母羊补饲精料,分别以日粮瘤胃可降解氮(RDN):可发酵有机物(FOM)为25和35的补饲效果最佳,且羔羊生长性能都良好[5]。后备母羊培育水平的高低,直接影响到后期的生产性能和繁殖性能。通过监测乌珠穆沁后备母羊四季放牧采食量,发现无法满足营养需求,且冬季蛋白质摄入量严重不足,还需要补充粗蛋白(crude protein, CP)、锌(zinc, Zn)、铜(copper, Cu)和硒(selenium, Se)[6-7],说明放牧后备母羊补饲也非常重要,但目前,针对放牧后备母羊补饲的营养调控研究报道较少,还需进一步深入开展。针对冷季后备母羊放牧条件下体损耗过大、生长发育受阻的问题,本研究设计科学合理的“放牧加补饲”方案,为有效缓解冷季放牧后备母羊体损耗,提高后备母羊培育质量,减轻草地压力,进一步提高牧民养殖效益提供数据支持。
1 材料与方法
1.1 试验地点
新疆富蕴县冬牧场位于阿勒泰富蕴县吐尔洪乡,草原类型为温性荒漠类。试验区海拔约为1 400 m,昼夜温差大,冬季寒冷漫长,枯草期逾7个月以上,试验期间平均气温-14 ℃左右。
1.2 试验动物与试验设计
1.2.1试验动物分组及管理
采用完全随机分组试验设计,选取8月龄40只体重为(38.50 ± 2.84) kg的阿勒泰后备母羊,经驱虫后,随机分成3个补饲组,补饲组每只羊每日补饲精料350 g,设置全价日粮瘤胃可降解氮(RDN):可发酵有机物(FOM)分别为35、30和25共3组,记作R:F35组、R:F30组和R:F25组,以及1个纯放牧组(CON),每组10只羊,试验期45 d,其中预试期10 d,正试期35 d。试验期每天10:00出牧前和18:30归牧后,采用补饲袋,对补饲组的每只试验羊进行精准补饲(精饲料饲喂量0.35 kg·d-1,早晚各半),保证试验羊精饲料采食完全,对照组不补饲。
1.2.2日粮组成及营养水平
在试验前期,通过链烷烃法准确测定纯放牧情况下后备母羊的牧草采食结构为:羽状三芒草(Aristida pennata)、芦苇(Phragmites australis)和沙生麻黄草(Ephedra sinica),其采食比例分别为22.52%、14.53%和62.93%。根据冯仰廉[8]提出的能氮平衡理论,选用RDN:FOM作为瘤胃能量与蛋白同步释放的匹配指标,参考各种饲料原料的有机物和蛋白质在瘤胃内降解特性参数设计后备母羊全价日粮配方RDN:FOM分别为35、30、25,以此为基础扣除放牧采食营养后确定补饲组的精饲料配方(表1)。其 中,全 价 日 粮RDN(g·kg-1) = 17.08 ×CP(%) +18.466 × (NDF/OM) - 162.029 (n= 24,R2= 0.876,P<0.01)。式中:NDF为中性洗涤纤维(neutral detergent fibers),OM为有机物(organic matter)。全价日粮FOM/OM(%) = 65.690 - 60.049 × (NDF/OM) (r= -0.912,n= 12,P< 0.01)[9]。精饲料加工成颗粒状(直径6 mm),其精饲料组成及营养水平如表1所列。
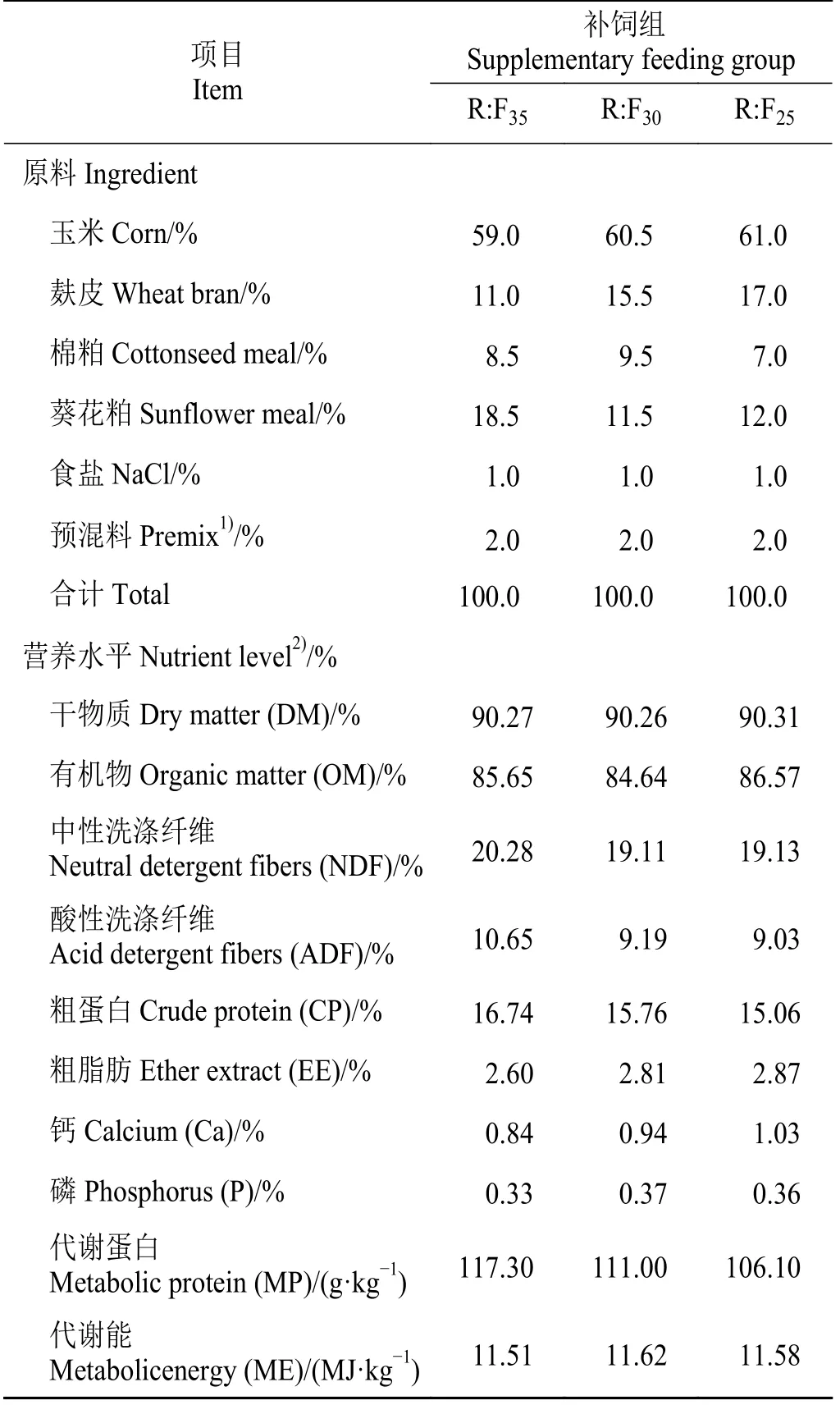
表1 精料组成及营养水平Table 1 Composition and nutrient levels of concentrates%
1.3 生长性能
正试期开始、结束日晨饲前,空腹称量每只母羊体重,计算日增重(average daily gain, ADG)。
1.4 血清生化指标
采用一次性真空采血管采集血液。在试验期结束前1 d,后备母羊每组10只羊,早上空腹前腔静脉采血。采血后3 500 r·min-1离心,之后使用l mL移液枪吸取血清到1.5 mL EP管,置于冰箱中(-20℃)保存。测定血清总蛋白(total protein, TP)、白蛋白(albumin, ALB)、球蛋白(globulin, GLB)、尿素氮(urea nitrogen, UN)、葡萄糖(glucose, GLU)、甘油三酯(total triglyceride, TG)、总胆固醇(total cholesterol,TC)、谷丙转氨酶(alanine aminotransferase, ALT)、谷草转氨酶(aspartate aminotransferase, AST)、碱性磷酸酶(alkaline phosphatase, ALP)、肌酐(creatinine)、钾(kalium, K)、钠(natrium, Na)、氯(chlorine, Cl)、钙(calcium, Ca)、镁(magnesium, Mg)、无机磷(inorganic phosphorus, IP)等指标。
1.5 试验数据处理与统计分析
试验数据采用Excel 2007进行初步分析整理,利用SPSS 19.1统计软件进行单因素方差分析和显著性检验,采用one-way ANOVA中的LSD法进行多重比较。试验结果以平均值 ± 标准差表示,以P<0.05表示差异显著,P< 0.01表示差异极显著。
2 结果与分析
2.1 补饲精料对放牧后备母羊生长性能的影响
试验末期,纯放牧组的体重每只减少了6.33 kg,3个补饲组变化不大,其中R:F35组增加了0.58 g(表2)。R:F35组、R:F30组和R:F25组的末体重较对照组分别极显著提高了20.48%、19.24%和17.90%(P< 0.01)。
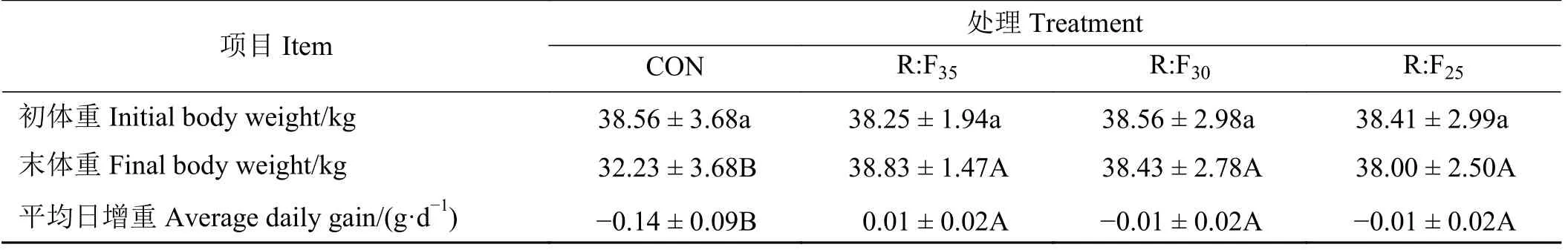
表2 补饲精料对放牧后备母羊生长性能的影响Table 2 Effects of supplementation with concentrates on the growth of grazing reserved ewes
2.2 补饲精料对放牧后备母羊血清GLU的影响
R:F35组和R:F30组放牧绵羊血清GLU较对照组分别极显著提高了121.98%和106.60% (P<0.01)。R:F25组放牧绵羊较对照显著提高了63.74%(P< 0.05) (图1)。

图1 补饲精料对放牧后备母羊血清葡萄糖含量的影响Figure 1 Effects of supplementation of concentrates on content of serum glucose (GLU) in reserved ewe
2.3 补饲精料对放牧后备母羊血清中蛋白代谢指标的影响
试验组TP与对照组无显著差异(P> 0.05),R:F30较R:F35组显著降低(P <0.05);各组间ALB和GLB无显著差异(P >0.05) (表3);补饲组的血清UN含量较对照组分别提高了22.19%、27.80%和12.33%,其中R:F35和R:F30组显著高于对照组(P<0.05);R:F35组的血清ALT最高,R:F30组最低,两组差异极显著(P< 0.01),但R:F30组较对照组无显著差异(P >0.05);试验组的血清AST活性均低于对照组,且随着RDN水平的降低而降低,R:F25组显著低于对照组(P< 0.05)。
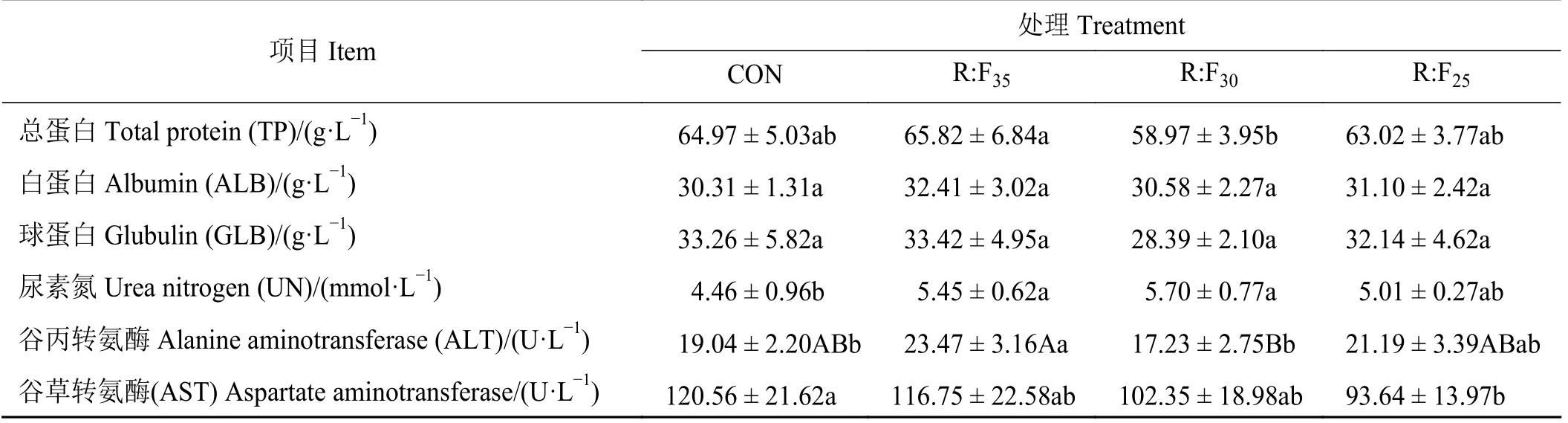
表3 补饲精料对放牧后备母羊血清中蛋白质代谢指标的影响Table 3 Effects of supplementation with concentrates on the serum protein metabolism of grazing reserved ewes
2.4 补饲精料对放牧后备母羊血清中脂肪代谢指标的影响
与对照组相比,R:F35组的血清TG较对照组增加了100% (表4),差异显著(P< 0.05),R:F30和R:F25组也增加了50%和71.43%;补饲各组的血清TC较对照组差异不显著,R:F30组最低,R:F25组最高,两组相差25.43% (P< 0.05)。

表4 补饲精料对放牧后备母羊血清脂肪代谢指标的影响Table 4 Effects of supplementation with concentrates on the serum fat metabolism of grazing reserved ewes
2.5 补饲精料对放牧后备母羊血清中矿物元素指标的影响
补饲精料对放牧后备母羊血清中K+、Na+、Cl-、Ca2+、Mg2+的含量无显著影响(P< 0.05) (表5);IP的含量较对照组分别提高了16.67%、1.31%和23.25%,其中R:F25组较对照组差异显著(P< 0.05)。补饲组血清ALP活性分别和R:F30组较对照组差异极显著(P< 0.01)。

表5 补饲精料对放牧后备母羊血清矿物元素代谢指标的影响Table 5 Effects of supplementation with concentrates on the serum mineral elements in grazing reserved ewes
3 讨论
3.1 补饲精料对牧区放牧后备母羊体重的影响
与其他季节相比,冬春季放牧后备母羊因牧草干枯,适口性差,且多冰雪覆盖导致采食量降到最低[6]。本研究对照组后备母羊的体重因营养物质摄入不足造成体重下降,较初期每只减少了6.33 kg,而补饲后,R:F35、R:F30、R:F253个试验组后备母羊体重变化较小,该结果与靳继鹏等[7]为甘肃高原细毛羊补饲高能量高蛋白水平日粮的结果一致,说明通过补饲可防止放牧后备母羊冬春季因摄入能量蛋白不足产生体损耗。此外,R:F35组的日增重较R:F30、R:F25组高,但差异不显著。以本研究日粮水平,日增重随着RDN水平的增加而提高,这可能与日粮RDN:FOM值逐渐接近零平衡,瘤胃微生物对RDN的利用率和瘤胃MCP的合成效率逐渐提高有关。张勇等[10]研究结果表明,补饲的饲料中能量蛋白越高,试验母羊的日增重越高,并有利于成年繁殖期的配种。本研究条件下,日粮RDN:FOM为35时,机体体重越高,有利于后期动物生产性能的提高。
3.2 补饲精料对放牧后备母羊血清中葡萄糖的影响
饲料中易降解的碳水化合物含量越高,饲料越容易被消化吸收。王文奇等[11]为放牧后备母羊补饲精料补充料,补饲后的表观消化率显著高于纯放牧组。血清GLU的来源主要为动物对饲料中的碳水化合物的消化吸收,维持着动物机体能量平衡。在甘肃高原泌乳的细毛羊放牧后补饲精粗比7 : 3颗粒饲料,其血清GLU含量显著高于纯放牧组,6 : 4组和5 : 5组血清葡萄糖含量则差异不显著[10]。本研究为放牧后备母羊补饲精料,随着RDN水平的增加,血清GLU浓度逐渐增加,且显著高于对照组,表明随着RDN水平的增加,促进了瘤胃微生物对FOM的消化降解,增加瘤胃液总挥发性脂肪酸(total volatile fatty acids, TVFA)含量,提高了血清葡萄糖水平;在本研究条件下,冬春季后备母羊补饲精料后,尽管日粮RDN:FOM为35时未达到蛋白水平的阈值,但放牧后备母羊可保持较好的能量代谢水平。
3.3 补饲精料对放牧后备母羊血清中蛋白质代谢的影响
血清中TP、ALB和GLB含量反映机体蛋白质的吸收、代谢以及免疫等情况,与日粮中蛋白水平正相关[12-13]。本研究中,R:F35后备母羊血清中TP、ALB和GLB浓度最高,提示日粮可降解氮的增加有提高机体对蛋白质的消化吸收、增加血清蛋白含量的趋势。血清尿素氮是动物机体摄入蛋白质、氨基酸代谢后的最终产物,含量升高不利于机体氮沉积和蛋白质平衡;本研究结果显示R:F35组和R:F30组的SUN显著高于对照组,表明补饲后,摄入蛋白含量增加,SUN含量也显著增加。该结果与张勇等[10]为泌乳母羊补饲不同能氮水平的精饲料研究结果相似,饲料可降解蛋白的增加对提高血清蛋白含量无显著影响,氮的利用率未得到显著提高。
血清ALT和AST通过转氨基的作用参与氨基酸代谢,本研究补饲精料后,R:F35组和R:F25组的ALT活性有所增加,可能与丙氨酸代谢增强有关;血清AST活性随着RDN水平的增加而增加,AST通过催化生成α-酮戊二酸和天冬氨酸进入三羧酸循环,可间接提高能量利用效率。本研究条件下,谷草转氨酶以及谷丙转氨酶都在正常范围内,且与甘肃高山细毛羊的补饲试验羊血清ALT和AST结果相近,说明补饲日粮对试验羊肝脏无不良影响,活性提高表明氨基酸代谢活动加强,有利于蛋白质的正常代谢[14-15]。
3.4 补饲精料对放牧后备母羊血清中脂肪代谢的影响
血清中胆固醇含量高低可反映动物机体脂肪代谢情况,胆固醇可与载脂蛋白结合,形成可溶性脂蛋白以维持细胞形态功能[16]。甘油三酯主要在肝脏和脂肪组织中合成,也可经小肠黏膜从食物中吸收合成,作为脂类代谢的中间代谢产物,其含量高低一定程度上反映脂肪代谢情况[17]。大鼠饲喂高糖日粮,可以增加内源性甘油三酯的合成[18]。低蛋白日粮补充合成氨基酸较常规日粮的甘油三酯和极低密度脂蛋白水平显著升高[19]。本研究中,补饲组血清TG升高,这可能与后备母羊摄入更多的瘤胃可降解有机物和可降解氮,使之转换为脂肪存贮于体内有关。
3.5 补饲精料对放牧后备母羊血清中矿物元素代谢的影响
李金朋[20]、李庆[21]和王永东[22]发现,血液中ALP与骨骼中的钙化密切相关,成骨细胞中的ALP作用产生磷酸,与钙结合生成磷酸钙存于骨中,且ALP还与营养物质的吸收转运过程相关。研究表明,ALP可有预防和减少肠道炎症、增加有益菌数量、调节钙吸收等功能[23]。较高浓度的无机磷可竞争性地抑制ALP的活性,Hoshino等[24]对大鼠的研究表明,低磷水平日粮升高了骨骼ALP活性。王凤来等[25]对小型猪的研究也表明,随磷水平升高血清ALP活性成下降趋势。本研究中,与对照组相比,R:F25组的血清IP浓度显著增加,但其ALP活性无显著差异;R:F30组和R:F35组的血清IP浓度较对照组差异不显著,但ALP活性显著提高,血清IP浓度与血清ALP活性未表现出相关性,且试验各组的血清IP和ALP均在正常范围内,Na+、K+、Cl-、Ca2+、Mg2+离子浓度也均无显著差异,表明试验各组日粮可基本满足放牧后备母羊的矿物元素代谢。
4 结论
综上所述,放牧后备母羊补饲不同能量水平的精料补充料可降低体损耗,维持体况水平,血清能量、蛋白、脂肪代谢水平显著提高,血清微量元素除无机磷外都无显著变化。
猜你喜欢
今日农业(2022年2期)2022-11-16 12:29:47
汽车工程师(2021年12期)2022-01-17 02:29:50
现代畜牧科技(2021年8期)2021-10-13 07:21:52
现代畜牧科技(2021年7期)2021-07-28 06:40:22
现代畜牧科技(2021年4期)2021-07-21 06:12:56
现代畜牧科技(2021年6期)2021-07-16 05:50:02
现代畜牧科技(2021年2期)2021-03-19 07:48:42
冰雪运动(2020年2期)2020-08-24 08:34:22
河南畜牧兽医(2020年1期)2020-01-11 05:00:25
河南畜牧兽医(2020年3期)2020-01-10 14:45:01
