
乌梁素海冰封期分层与混合特征及对氧代谢速率的影响*
2022-05-17 08:13:06杨惠杰黄文峰李志军林战举
湖泊科学 2022年3期
杨惠杰,黄文峰,张 程,李志军,林战举
(1:长安大学,旱区地下水文与生态效应教育部重点实验室,西安 710054) (2:长安大学,水利与环境学院,西安 710054) (3:大连理工大学,海岸与近海工程国家重点实验室,大连 116024) (4:中国科学院西北生态环境资源研究院冻土工程国家重点实验室,兰州 730000)
全球超过一半的湖泊位于高纬度或高海拔寒区[1],普遍存在湖冰层生长与消融过程. 气候变暖在寒区则更为显著,导致湖泊稳定性增加,甚至改变混合机制[2],湖底缺氧区域与延续时长因分层期的延长而扩大、延续[3],威胁湖泊水生态安全. 湖冰过程对气候变暖较为敏感,北半球湖泊冰封期变短、冰厚变薄被广泛报道[4-5],也是引起高纬度湖泊环境与生态系统演变的关键驱动[6]. 然而,冰封条件下湖泊环境与生态特征及其驱动过程仍不清晰. 国内外学者通过结冰前、结冰中、融冰后大量采样,测试湖泊理化指标、营养盐、浮游植物等的变化,证实了湖冰过程对湖泊环境与生态的显著影响[7-9].
湖泊分层与混合过程是湖泊系统中最基本的物理过程之一,也是湖泊分类的依据之一,对湖泊水温、溶解氧结构,叶绿素、营养盐分布等具有显著影响[10-11]. 冰封后,冰层切断了风应力驱动的湖泊混合过程,但冰下水体仍存在弱水动力过程,最为显著的便是太阳透射辐射驱动的垂向混合过程. Bengtsson等[12]、Volkov等[13]在Vendyurskoe湖的观测揭示了透射辐射驱动的冰下水体湍动混合过程. Bouffard等[14]、 Bogdanov等[15]指出白天较强的辐射会驱动较深的混合层出现;Yang等的观测也指出晚冬辐射驱动的对流会导致混合层加深,且冰下溶解氧结构与混合强度密切相关[16]. 但是,相较于水面开放期,冰下水体混合强度较弱,其能否或能在多大程度上驱动水环境发生改变仍需进一步研究.
水温和溶解氧是评价水环境优劣的基础指标,其变化过程对湖泊生态系统有重要的影响. Golosov等[17]研究俄罗斯和北美5个冰封湖泊缺氧层的形成与发展时就发现冰封湖泊底部容易形成缺氧区域. Bai等[18]研究了芬兰Valkea-Kotinen湖冰封期深层溶解氧特征,发现溶解氧浓度随着冰封延续而下降,且在一定深度出现持续缺氧区域. Yang等[16]在加拿大Simcoe湖观测到溶解氧随深度衰减且在底部存在氧衰竭现象. 上述研究基本揭示北欧、北美等地典型结冰期湖泊溶解氧的变化动态. 然而,我国结冰湖泊多位于干旱半干旱区(如蒙新高原湖区、青藏高原湖区),冬季降雪少、辐射强,不同的气候、气象、天气条件决定了其不同于北欧、中欧、北美等地的湖泊[19],亟需明确冰封条件下其温氧环境的演化特征及其驱动过程.
乌梁素海是我国干旱半干旱地区典型的浅水湖泊,是河套灌区生活污水、农田灌溉排水、退水的承泄场所,对于缓解黄河下游污染、水土保持、流域环境改善都有极其重要的作用. 近年来,冰封期乌梁素海环境与生态不断引起重视,不但认识到冰、雪过程对乌梁素海环境与生态的重要影响[20-21],也开始深入探索冰封期水温、溶解氧、新陈代谢动态变化与内在机理,如翟佳伦等[22]用小波分析对乌梁素海水温与溶解氧变化趋势进行研究得到了其不同周期特征,Song等[23]通过乌梁素海冰封期新陈代谢速率计算指出冰、雪过程对湖泊新陈代谢的影响.
本文以寒旱区典型湖泊乌梁素海为例,开展了冰封期水温、溶解氧、冰/雪过程和气象要素的现场监测与分析,并以2017-2018年冬季观测结果为例,介绍冻结期和融化期乌梁素海水温、溶解氧分层结构、混合过程演化特征和氧代谢速率,着重分析冰下混合层的形成与发展及其对代谢速率的影响,基于4个冬季的观测对比分析乌梁素海分层与混合结构的年际变化和冰下混合形成机理,最后给出重要结论.
1 数据获取与处理
1.1 观测地点
乌梁素海(40°47′~41°03′N,108°43′~108°57′E)位于内蒙古自治区巴彦淖尔市乌拉特前旗境内(图1a). 为中国八大淡水湖之一,也是全球荒漠及半荒漠地区极为少见的大型草原型湖泊、地球同纬度最大的湿地,为干旱及半干旱地区具有代表性的浅水湖泊. 其面积约为310 km2,平均水深1.5 m,整体南北长(35~40 km),东西宽(5~10 km). 湖岸线长度为130 km,储水量约为3×108m3. 湖泊呈富营养化,N、P元素严重超标[24].
乌梁素海属温带大陆性气候,年降水量仅215 mm,超过70%集中在7-9月[25]. 年蒸发量达2200 mm,夏季湖面蒸发占全年蒸发量的50%. 湖区pH约为8.1,多年平均气温为5.6~7.4℃[21],年内平均气温7月最高(25℃),1月最低(-10℃). 每年11月中下旬开始结冰,最大冰厚为0.3~0.7 m,出现在2月底至3月初;翌年3月开始消融,冰封期约4~5个月[20]. 冬季偶有降雪,冰/雪面常有沙尘沉降.
1.2 数据获取
观测点在乌梁素海东部远离芦苇的开阔区(图1b),水深1.42 m. 2016-2019年4个冬季冰封期连续观测气象要素、冰雪条件与冰下水体环境. 以2017-2018冬季为例,在冰面架设自动气象站实时监测2 m和6 m处气温、气湿、风、太阳辐射等;布设温度链监测冰面以下0、5、10、15、20、25、30、35、40、44、50、54、59、64、69、79、89、99.3、109、119、129、144、152、160、173 cm处冰、水、底泥温度;利用CSI CS511监测冰面以下50、70、90、110、130 cm处水体溶解氧(DO)浓度;同时,利用Trios辐照度探头测量冰面以下65、90、120 cm处的光合有效辐射(PAR)入射和98 cm处PAR反射. 数据观测与可用时段见表1,现场布置见图1c,其他详见文献[25].

表1 数据观测与可用时段
1.3 混合层的判定
混合层是对“等温层”或“等密度层”的一个大体表述[26],特点是湍流作用显著,温度、密度以及水平流场在垂直方向上基本均匀分布[27]. 垂向水温梯度被广泛用于湖泊混合层的判定. 当水温梯度小于某临界值,则可认为该水层为混合层. 不同湖泊,该临界值大小存在差异(0.02~4℃/m)[28-29]. 据监测温度(T)的探头所在深度(z)将乌梁素海垂向划分成层,相邻两层深度和水温分别记为Zi、Zi-1和Ti、Ti-1;考虑到该湖水深浅,混合层由式(1)判定:
(1)

1.4 代谢速率计算
冰封湖泊与大气之间氧气交换被冰层隔断,则溶解氧变化动态仅由冰下水体光合产氧、水体生化耗氧与底泥耗氧过程收支平衡控制. 若将水体和底泥生化耗氧过程统一看作湖泊生态系统的呼吸作用,则冰封湖泊溶解氧变化是由光合产氧和呼吸耗氧造成[30],即:
(2)
夜间无光照,水体溶解氧浓度降低仅由呼吸作用控制,夜间呼吸耗氧速率(Rnight)可由实测溶解氧的降低速率计算(式(3)). 假设白天的呼吸耗氧速率与夜间平均呼吸耗氧速率相等[23](式(4)),则光合产氧速率(GPP)可由式(5)计算:
Rnight=NEPnight=(DOt+1-DOt)/Δt
(3)
(4)
GPP=Rdaytime+NEPdaytime
(5)
式中,DOt和DOt+1为时间间隔Δt的两个时刻的溶解氧浓度;NEP、GPP、R分别指净光合产氧速率、光合产氧速率与呼吸耗氧速率,mg/(L·h),下标night、daytime分别表示各项于夜晚、白天的值;为去除溶解氧垂向扩散运移的影响,将上述各式在整个水柱上积分后取水深平均值.
2 水温与溶解氧动态
2.1 气象与冰雪过程
现场监测覆盖了冰生长后期以及冰消融前期(图2). 日均风速在2.0~4.5 m/s之间,最大风速超过10 m/s. 监测期内平均气温-12℃,1与2月平均气温分别为-13.61、-9.77℃. 开始观测时冰厚已有31 cm,在1月9日-2月15日期间,冰以0.25 cm/d的生长速率缓慢生长,随后,以0.53 cm/d的消融速率消融(图2e),偶有少量降雪.
1月9日-21日,气温上升,相对湿度下降,雪因吸收辐射消融与风吹蚀而变薄,冰生长被抑制;随后,气温降低,冰生长略加快(图2e①箭头),相对湿度较大;1月27日发生降雪,冰生长速度变缓(图2e②箭头). 雪因吸收辐射消融和风吹漂移作用,至2月7日变为裸冰面. 2月13日后日最高气温在0℃上下波动,冰层停止生长,并于2月16日以0.53 cm/d的速率消融. 入射与反射PAR均具有明显的日变化过程,在12:00-13:00之间达到峰值. 入射PAR呈增大趋势,平均值为51 W/m2;反射PAR呈减少趋势,平均值为28 W/m2.

图2 气象与冰雪过程(a:风速;b:气温;c:相对湿度;d:雪厚;e:冰厚;f:总入射、65 cm处入射及总反射PAR)Fig.2 Meteorology and ice and snow process (a: wind speed; b: air temperature; c: relative humidity; d: snow thickness; e: ice thickness; f: incident and reflected photosynthetically active radiation)
2.2 水温与溶解氧动态变化
冰下各层水温与溶解氧呈现明显的日变化特征,其日变幅随深度衰减且对辐射与气温的响应逐渐滞后(图3a,b). 无雪层覆盖时,水柱平均水温最小值出现在8:00左右,最大值出现在17:00左右,日变幅约为1℃;溶解氧最小值出现在7:00左右,最大值出现在18:00左右,日变幅可达3 mg/L. 有雪层覆盖时,水温降温更为平缓且最小值的出现时间因雪厚不同表现出不同程度的延迟,日变幅降低(约0.6℃),这与雪层保温效应有关.
但根据图3c,相比于无雪条件下,并没观察到水温最大值出现的时间显著变化或仅是稍微延迟(与雪层保温效应有关). 这主要是因为:尽管经历多次降雪,但每次降雪量很小,又因多强风,冰面堆积厚度很薄(不足3 cm),且呈斑块状分布;其存在会在一定程度上削弱太阳辐射的透射量,但仍有少部分太阳辐射能够穿透冰雪层直接加热上层水体,引起水温小幅度的日变化(白天的升温);这一升温过程应与太阳投射持续累积加热同步,不会存在明显的时间差异.
有雪覆盖时溶解氧浓度变幅较小且降低的持续时间长,1月15日直到12:00左右才开始升高(图3d). 这是因为冬季湖泊溶解氧浓度日变化主要受控于生物过程[31],而影响生物过程的太阳辐射和水温在水体中的分布受冰、雪过程的影响. 整体来看,深层日变幅小于浅层,极值出现时间也推迟,比如69.2 cm处水温比39.8 cm处极小值滞后约2 h.
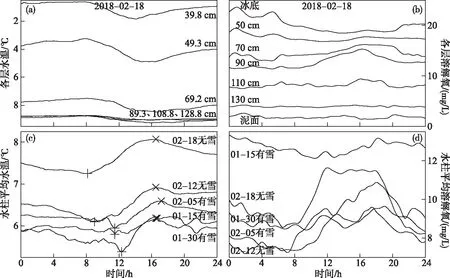
图3 水温及溶解氧浓度的日变化(a:2018年2月18日各层水温,b:2018年2月18日各层溶解氧浓度, c:水柱平均水温,+为最低值,×为最高值,d:水柱平均溶解氧浓度)Fig.3 Daily changes of water temperature and dissolved oxygen (a: water temperature of each layer on February 18, 2018;b: dissolved oxygen concentration of each layer on February 18, 2018; c: bulk water temperature, + is the lowest value, × is the highest value; d: bulk dissolved oxygen concentration)
水温在1月21日前呈上升趋势,主要是因为逐渐变薄的雪盖层使得更多的辐射进入水体,随后的降雪又使其呈现下降趋势直到2月初,之后缓慢上升;水体溶解氧浓度则先下降,再波动升高(图4). 1月27日-2月7日,冰生长几乎停滞且存在降雪及融雪过程,雪层的存在能削弱进入水体的太阳辐射,减少气温以热传导方式传递给水体的热量,但较薄的雪层影响有限,水温仍受气温影响而升高,湖底缺氧水层迅速发展(图5b).
水温结构最初表现为上部为逆温层(水温梯度为19.2℃/m)、中间为弱正温层(-4.7℃/m)、下部为弱逆温层(1.4℃/m)(图4c黑实线). 这种水温结构的出现与该时段存在冰、雪过程,尤其是雪过程有关. 积雪较大的反照率使得进入水体的太阳辐射变小,而湖底由于沉积物释热保持较高的温度,导致在中间部分接受能量较少而出现局部正温层;2月7日起,雪消融后,太阳辐射能够到达的深度逐渐变深,上部逆温层接收的辐射最多,其下边界随之变深. 当水温升至约7℃后,水层失稳,产生混合,混合层不断加深;过渡至上部为逆温层(18.2℃/m)、下部为近等温层(0.04℃/m)结构(图4c红实线).
溶解氧浓度的垂向结构不同于水温,由上至下逐渐衰减(-17 mg/(L·h),甚至在水层底部出现缺氧(<2 mg/L[32])状态(图5b). 主要是因为水层顶部接受PAR更多,光合更强烈,而呼吸则因水温低较缓慢,再者冰层冻结排氧会增加表层附近溶解氧;底部水温较高,呼吸速率快,底泥耗氧明显加快而出现缺氧状态.

图4 观测期内水温及溶解氧浓度的变化与垂向结构(a:观测期内水温及溶解氧浓度的变化趋势; b:溶解氧浓度的垂向变化;c:水温的垂向变化)Fig.4 Changes and vertical structure during the observation period of water temperature and dissolved oxygen concentration (a: trend during the observation period of water temperature and dissolved oxygen concentration; b: vertical change of dissolved oxygen concentration; c: vertical change of water temperature)
2.3 湖泊代谢速率
光合产氧速率1月18日前呈升高趋势,随后波动降低,至1月27日后又呈升高趋势,2月24日达到0.55 mg/(L·h)的最大值(图6a). 观测期光合产氧速率平均值为0.23 mg/(L·h). 呼吸耗氧速率趋势与光合产氧速率变化过程相似,相关性较高(R2=0.73),观测期间平均呼吸耗氧速率为-0.091 mg/(L·h),最大耗氧速率出现在2月18日,值为-0.252 mg/(L·h). 净生产力是每天光合产氧与呼吸耗氧平衡作用的结果,围绕0上下波动,观测期间平均为-0.006 mg/(L·h).
3 混合层的发展及其对代谢速率的影响
观测后期,自2月8日水体出现稳定而持续的等温层,0.9 m下水温随后整体从6.7℃逐渐升高至9.9℃(图4c,5a). 若以该层水温白天升高0.5℃所需的太阳辐射强度作为白天日均辐射的有效影响深度,则2月17日以后,太阳辐射可照射至湖底(图5a红实线). 自此之后,因太阳辐射在水体中传输满足指数衰减规律,该等温层内从上至下吸收的太阳辐射逐渐降低,则其升温幅度也应出现从上至下逐渐变小的趋势,即上部升温快而大、下部升温慢而小;然而实测的水温廓线从次日后仍维持上下等温状态(图4c, 5a),所以若未出现混合过程,该层水体无法维持稳定而连续的等温状态.
以水温梯度(式(1))绝对值等值线图方式表现混合层形成与发展过程更为直观(图6b). 受雪盖层影响,观测初期(1月9日-2月3日)未形成明显而稳定的混合层. 随积雪融化变薄,透射增加,水温升高至约7℃以后,连续的水体混合从中层水体(110 cm)开始出现,形成局部混合;之后其不断向上、下扩展,至2月14日以后混合层几乎占据整个下层水体(90 cm以下).
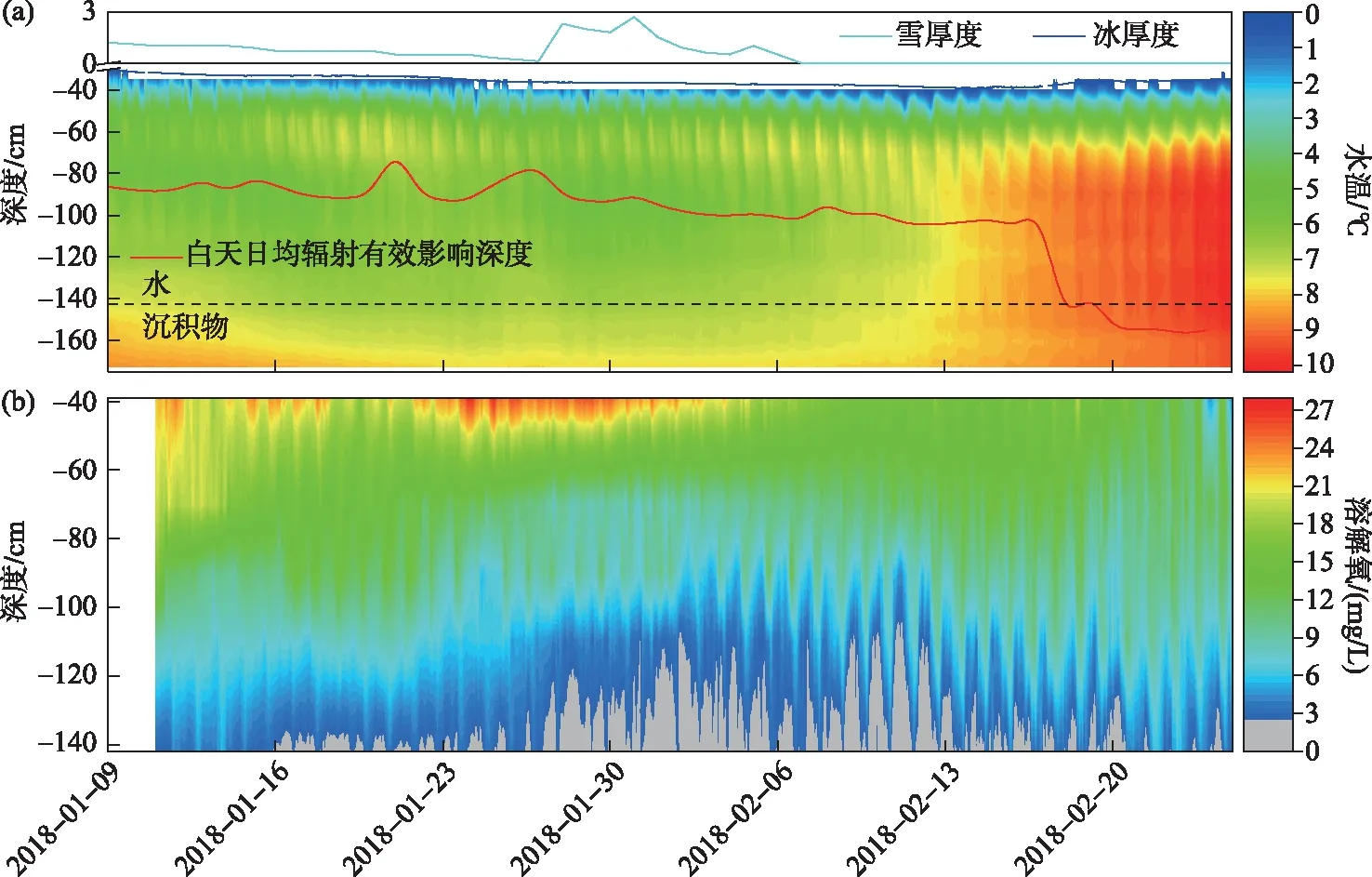
图5 观测期内水温与溶解氧浓度的变化(a:水温等值线图;b:溶解氧等值线图)Fig.5 Changes in water temperature and dissolved oxygen concentration during the observation period (a: water temperature contour map; b: dissolved oxygen contour map)

图6 新陈代谢速率变化与混合层发展过程(a:新陈代谢速率;b:混合层发展过程)Fig.6 Metabolic rate change and mixed layer development process (a: metabolic rate; b: mixed layer development process)
依混合层发展过程,将观测期划分为两个阶段:1月9日-2月3日的间断性小范围弱混合或不混合阶段和2月3日到观测结束的混合快速发展与持续稳定混合阶段. 从第1阶段至第2阶段整体水温从6.0℃升至7.3℃,仅升温1.3℃;冰下总PAR由14.8 W/m2升至22.8 W/m2,增加54%. 而光合产氧速率第2阶段是第1阶段的3.2倍,呼吸耗氧速率是第1阶段的2.5倍(图6a). 显然,除了水温升高、PAR增大促进光合和呼吸作用外,水体混合伴随的溶解氧再分配、底部浮游植物和营养物质悬浮必然也会促进代谢速率加快. 例如,溶解氧下移和底部营养物上移能够增强底部缺氧层和底泥的耗氧作用,致使湖泊总呼吸速率大幅度升高;垂向混合使得底部营养盐、浮游植物向上输移至上层,光照强度更大,促进光合作用产氧速率.
4 讨论
4.1 水温、溶解氧浓度的年际差异及同其他湖泊的比较
本文观测处于乌梁素海冰层冻结后期向融化期转变的过程,冻结期水温结构主要由稳定的上部逆温层和下部弱逆温层构成,并逐渐过渡为融化期的上部逆温层-下部混合层结构. 这种水温结构在结冰的淡水湖泊中较为常见[33-34]. 但乌梁素海作为中纬度寒旱区浅湖的典型代表,其冰封期水温结构也呈现出不同之处:1)上部逆温层和冻结期的下部弱逆温层的温度梯度分别为19.2和1.4℃/m,远高于北欧、中欧、北美等湖泊,主要原因是水浅[35]、底泥温度高等;2)整体水温高,下部弱逆温层或混合层温度达到9~10℃,而高纬度湖泊多接近冰点或不高于4℃[36-37],主要是因为纬度低辐射更强、干旱半干旱气候决定冬季降雪少,裸冰面使得穿透辐射更多而加热水体;3)冻结末期和融化初期的下部混合层(等温层)并非每个冬季都会出现. 2016-2019年4个冬季中(图7),仅有2015-2016、2017-2018年两个冬季出现大范围混合层(图7a、c红实线),且发现只有当中下层水温升高至6~7℃以后才会出现明显的混合过程,这与乌梁素海的盐度垂向结构有关.

图7 2016-2019年水-泥温度廓线图Fig.7 Water-sediment temperature profile of 2016-2019
乌梁素海溶解氧浓度的垂向结构形态与芬兰、俄罗斯等国家湖泊的监测结果[37]基本一致,但值比其高,主要是因为乌梁素海纬度较高,太阳辐射强,覆雪薄且水浅,故冰下一定深度仍能进行光合作用. 融化期形成的低氧融水也可使冰下方附近溶解氧浓度降低,这与Meding和Jackson在北美温带湖泊的观测结果一致[38]. 乌梁素海底层水体缺氧区域的发展与持续时间与冰封时间长短及是否存在雪盖层关系密切,如2月8日前连续的冰面积雪存在使得湖泊底部缺氧现象逐渐加重(图5). 较少的光合产氧可能会对湖底沉积物呼吸耗氧造成胁迫作用,这是辐射通过光合作用对呼吸作用的间接控制[23],也是光合作用产氧与呼吸作用耗氧存在相近趋势的可能原因之一.
观测期光合平均产氧速率为0.23 mg/(L·h),最大值为0.55 mg/(L·h). 呼吸耗氧速率平均值为-0.091 mg/(L·h),最大值为-0.252 mg/(L·h). 光合产氧速率高于Obertegger等在意大利Tovel湖研究测得的值(最大约为0.3 mg/(L·h)[39]),这是因为Tovel湖为贫营养湖,无法支持高的代谢速率. 第1阶段代谢速率(平均GPP为0.112 mg/(L·h),平均R为-0.054 mg/(L·h))与Song等在该区域于2016-2017年所测得的结果(GPP最大值约为0.2 mg/(L·h),R最大约为-0.1 mg/(L·h))基本一致. 但第2阶段(即混合阶段)代谢速率(平均GPP为0.356 mg/(L·h)、平均R为-0.133 mg/(L·h)),高于Song等[23]于2016-2017年冬季测得的结果. 这一年际差别主要是由混合层的贡献导致,2017-2018年冬季出现了大深度的水体混合过程,促进了代谢过程;而2016-2017年冬季并未形成明显的混合过程.
前人监测或调查结果显示,非冰封期(尤其是夏季)乌梁素海水温较高(均大于20℃)[40-41],冰封期温度低,主要是因为冰雪层极大削弱穿透的太阳辐射,并且水体持续向冰层放热[33];但溶解氧浓度高于乌兰等[42]于2014-2015年7和9月的观测值(分别为6.1和5.8 mg/L),一方面是因为冰封期较低的水温降低了耗氧速率,另一方面是由于冰盖层的存在阻隔了溶解氧的逸散. 除此以外,还可能与逐年上升的溶解氧浓度趋势有关[43].
4.2 冰封期乌梁素海混合层的形成机制
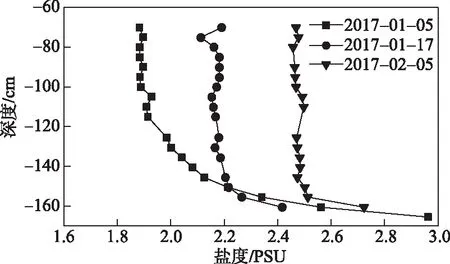
图8 2016-2017年盐度廓线图Fig.8 Salinity profile of 2016-2017
冰封湖泊混合层的发展过程与透射辐射密切相关[44]. 一般而言,对结冰的淡水湖泊,因结冰前的风动力混合和冻结期水体不断向冰层释放热量,开始融化前水体温度多低于4℃,甚至整体接近冰点. 融化期,表面积雪融化消失和增强的太阳辐射促使更多辐射穿透冰层,对水层形成垂向差异化加热(从上至下加热升温速率逐渐降低),上部升温快而大,更接近最大密度温度(4℃),引起水体上下失稳混合[34,44]. 然而乌梁素海却呈现不同的水温结构和混合形成特征,冻结期整个水层维持着逆温结构,上层水体温度低于4℃而中下水层温度通常高于4℃,但此时并未在接近4℃时发生明显的混合现象(图7),这是因为自由混合过程是由水温和盐度共同控制的. 乌梁素海实际是微咸水湖,水样测试表明,其盐度在上、中水层垂向差异较小,而近底部分层较稳定(图8). 根据水状态方程[45],盐度分层对水密度的影响比温度更为显著. 根据2016-2017的采样盐度廓线数据,只有当中下部水温接近或超过6~8℃(超过4℃)时,水体密度结构才能出现失稳反转,这与2015-2016、2017-2018年冬季的观测结果一致. 但是由于沉积物表面附近的高盐度,混合层无法深入到底泥表面,无法进行全深度混合.
混合层对水温结构的影响最为直观,使水温结构成为判断混合层是否存在的标志之一[26]. 以2016-2017和2017-2018年为例,前者未出现混合而以逆温形式存在,后者后期出现混合层而在冰下冷界面层下有以近等温形式存在的对流混合层(图7b,c). 水温结构的改变必然会改变以热导方式进行的气-冰-水-泥体系热传递,影响能量在其中的分布. 对流混合对溶解氧结构的影响包括直接和间接两个过程,一是水体垂向混合会直接引起溶解氧的垂向混合,趋向于等氧状态,如,2017-2018年冬季混合前的分层状态下,湖底缺氧层随分层结构的延续由湖底扩展到冰下1.1 m(2月13日),混合后湖底缺氧状态得到极大的缓解(图5);二是垂向混合可以使微藻、营养盐上移接收更多的光照而促进光合产氧[46-47],还可以使得溶解氧向下迁移至低氧区域甚至底泥附近而促进底层生化耗氧,共同造成上部氧多、下部氧少的趋势,这也解释了为什么混合并未引起上下等氧状态(图4b).
5 结论
对乌梁素海结冰条件下水体温度、溶解氧的分层结构和混合动态开展原位监测与理论计算分析,取得以下结论:
1)乌梁素海在冻结中后期仍保持较高的水温(接近10℃),冻结期水温垂向结构主要由稳定的上部逆温层和下部弱逆温层构成,并可逐渐过渡为融化期的上部逆温-下部混合层结构. 混合过程并非每年都会出现或出现日期不同,取决于冰雪条件、底泥温度和盐度特征.
2)结冰期溶解氧浓度较高,由上到下降低,出现湖底缺氧层,其发展与持续时间与冰封时间长短及是否存在雪盖层关系密切. 溶解氧动态由光合产氧和呼吸耗氧平衡决定,而光合产氧、呼吸耗氧速率的变化动态主要受太阳辐射强度、水温和混合过程影响. 混合层的形成与发展受水体盐度与温度共同控制,其能极大地促进光合产氧和呼吸耗氧作用、调控水温和溶解氧结构.
辐射强、降雪(冰面积雪)少是中纬度寒旱区湖泊区别于其他地区的显著特征,其对冰封期水温、溶解氧、代谢速率和分层动态的影响作用在寒旱区具有普遍性. 但冰下水体分层与混合特征的年际差异也说明不同时期或不同湖泊具有明显个体差异,未来冰雪、温(含辐射)、盐同步观测将能更细致地揭示冰封期不同湖泊混合过程及冰雪条件的影响.
致谢:感谢内蒙古农业大学乌梁素海湿地生态系统国家定位观测研究站提供的技术支持和后勤保障.
猜你喜欢
百科探秘·海底世界(2022年6期)2022-06-24 09:37:51
疯狂英语·新读写(2021年8期)2021-11-05 08:44:24
奇闻怪事(2021年2期)2021-04-20 12:12:01
家庭科学·新健康(2020年7期)2020-08-06 08:15:35
南方农业·中旬(2020年2期)2020-05-28 13:47:28
少儿美术(快乐历史地理)(2019年11期)2019-04-20 12:33:24
中国三峡(2017年4期)2017-06-06 10:44:22
中国海洋大学学报(自然科学版)(2017年7期)2017-06-05 15:08:52
水利科技与经济(2016年8期)2016-04-22 03:41:12
资源节约与环保(2015年10期)2015-03-17 08:36:30
