
半干旱区草地土壤碳呼吸不同组分的初步区分
2022-05-16 09:08张琳焓张雪腾左大鹏
智慧农业导刊 2022年9期
张琳焓 ,张雪腾 ,左大鹏
(1.兰州大学 大气科学学院,甘肃 兰州730000;2.甘肃省半干旱气候与环境野外科学观测研究站,甘肃 兰州 730000)
干旱半干旱区占全球陆地面积的41%,该区域降水稀少,蒸发量大,植被覆盖稀疏,生态系统十分脆弱[1],是全球陆地表面增温最显著的区域[2],主导着全球碳循环的年际变化。研究表明,虽然热带雨林是全球陆地生态系统中最重要的碳汇,但是其具有较大的降水和气温年变率;半干旱区草地系统对全球陆地碳汇年际变异的贡献约为39%,显著高于热带雨林的19%和农田的27%[3-4]。因此,全面阐明半干旱地区不同草地类型土壤呼吸作用的变化规律及机理,是准确评估全球尺度碳平衡收支的重要基础。
土壤呼吸通常是指未扰动土壤中产生二氧化碳(CO2)的所有代谢作用,包括根系呼吸、土壤微生物和动物呼吸以及含碳矿物质的化学氧化作用[5-6]。Schimel等[7]指出,植物根系的自养呼吸和土壤微生物及动物的异氧呼吸占生态系统总呼吸的60%~90%。我国学者对内蒙古温带草原[8-10]、东北平原羊草草原[11-12]和青藏高原高寒草甸[13-14]不同组分土壤呼吸的日变化、季节变化及年际变化特征及其影响开展了大量的研究工作,刘立新[15]等研究表明,内蒙古锡林河流域草地根系呼吸速率占土壤总呼吸的25%~45%,平均值为35.66%。贾子毅[16]研究发现,内蒙古荒漠生态系统中白刺叶片净光合速率、土壤温度和水分的三因素模型可以解释土壤呼吸的58%。而目前对于黄土高原草地生态系统不同组分土壤呼吸的关注较少,Li等[17]结合LI-6400型动态气室和挖壕沟法区分了根系和土壤微生物呼吸对黄土高原半干旱区草地系统土壤呼吸的相对贡献,指出前者的日变化由植物的光合作用主导,而后者主要受土壤温度的影响。以往的研究主要集中于根系和土壤微生物的呼吸作用,而忽略了其他组分如叶片的贡献,Wang等[18]发现黄土高原半干旱区在夏、秋季自然草地植被的高度最大可达20~30 cm,如果忽略了这部分呼吸作用将对准确评估草地系统碳源汇产生一定影响。因此,定量区分根系、叶片冠层和土壤微生物呼吸等不同组分的贡献对我国草原生态系统碳平衡过程的全面理解具有重要的意义。
本文对比分析了黄土高原半干旱区原生草地生态系统总呼吸、根系呼吸、叶片冠层呼吸和土壤微生物呼吸的日变化特征,探究了降水、土壤温度和含水量对不同组分呼吸速率日动态的影响,讨论了不同组分呼吸对草地生态系统总呼吸的贡献率,为定量评估我国黄土高原地区草地系统的碳源汇收支及其对全球气候变化的响应提供重要数据基础。
1 研究区域概况
研究区域设置在兰州大学半干旱气候与环境观测站(下文中简称“SACOL”),占地面积约为120亩,位于甘肃省榆中县兰州大学榆中校区萃英山顶(35.946°N,104.137°E,海拔1 965.8 m),距兰州市东南方向约48 km。气候类型属于大陆性半干旱气候,年平均气温6.7℃,多年平均降水量382 mm,主要集中在7-9月,年变率比较大,年蒸发量高达1 340 mm,日照时数约2 600 h[19]。实验场地原为耕作农田,从1986年被弃耕后已恢复成天然草地,自2005年围封以来已连续16年禁止放牧,受人类活动干扰较少,可代表周边几百公里内半干旱草地的气候状况。土壤类型为栗钙土,0~10 cm土壤层的有机碳、全氮、pH值和土壤容重分别为8.96 g/kg、0.75 g/kg、8.45 和 1.14 g/cm3,10 cm~20 cm 土层的对应值分别为 7.42 g/kg、0.69 g/kg、8.49 和 1.14 g/cm3[20]。植被类型为半干旱地区典型的天然草地,主要有铁杆蒿(Tripolium vulgare)、冷蒿(Artemisia frigida)、赖草(Leymus secalinus)和本氏针茅(Stipa bungeana)等[21],其生长主要受降水量和气温的控制;总体而言,在夏秋季节植被生长较茂盛,而在冬春季节草地植被逐渐演变枯黄。
2 数据资料和研究方法
采用 LI-8100A型土壤碳通量仪(LI-COR,Lincoln,USA)全天时连续测定土壤呼吸碳通量速率,基本原理是利用动态密闭气室红外二氧化碳方法测量室内CO2浓度的增加速率来推算室外土壤CO2扩散到空气中的速度[22]。初始安装时,在SACOL站内选择了一块大小为20 m×20 m的平坦样地,整体而言,实验场地内植被分布较均匀,但在局地尺度上,有多种类型的植被和裸地的分布,为此我们选择了5个不同植被覆盖度的点安装了长期测量气室,分别标记1~5号,每个气室之间相隔4 m。为了减少安装对观测的干扰,提前1 d将各气室的底座PVC环(直径21 cm,高11 cm)埋入土壤中,嵌入深度6 cm,土壤环露出地面4 cm。不同气室中的植被覆盖如图1所示,其中1号和2号气室内分别为常年干枯植被和无根裸地,而3~5号气室为不同覆盖密度的茂盛草植被。为了区分根系、叶片冠层和土壤微生物对草地生态系统总呼吸的相对贡献,我们于2020年5月16日剪除了3号和4号气室内地上的叶片和茎,只保留其地下的茂盛根系。为了尽可能减少剪草试验操作对观测的干扰,我们选定剪草后的第6天开始分析。

图1 LI-8100A型土壤碳通量仪5个气室内的植被分布(拍摄于SACOL站,2020年5月16日剪草前)
本文的数据时段为2020年5月21-31日。实验期间,每30 min对5个气室依次完成一轮测量,即每天每个气室会进行48次土壤呼吸测定,同时仪器自带的土壤温度探针(EC-5型)和土壤水分传感器(GS-1型)可实时探测5 cm的土壤温度及含水量。此外SACOL站内同期还测量了5层土壤含水量(5~80 cm;CS616-L型)和土壤温度(2~80 cm;STP01-l50型),太阳总辐射(CM21型)、光合有效辐射(PAR-Lite型)和降雨量(TE525MM-L型)数据,分辨率均为30 min。
3 结果分析
3.1 剪草后土壤呼吸速率变化规律
3.1.1 降水、土壤参数和太阳辐射的时间变化序列
剪草后的5月下旬出现了3次明显的降雨过程(图2a),其中,23 日的降水从 8:00 开始一直持续到 20:00,小时最大降水量达3.2 mm,总降水量为8.5 mm。26日和30日的小时最大降水量分别为2.9 mm和2.0 mm,总降水量分别为9.1 mm和8.8 mm。降水过程造成的雨水下渗对不同深度土壤水分产生不同程度的影响(图2b),5 cm土壤含水量的增幅最显著,从0.12 m3m-3增加到0.17 m3m-3,10 cm和20 cm的含水量分别从0.124 m3m-3增加到 0.149 m3m-3和从 0.104 m3m-3增加到0.114 m3m-3,40 cm和80 cm层土壤含水量的增幅较小,本区域内植被根系主要分布在0~20 cm土层,各层土壤含水量的增加会促进根系呼吸和土壤微生物的活性,进而间接影响到生态系统的碳呼吸作用。受降水过程的影响,太阳总辐射和光合有效辐射的最大值分别从晴天的1 100 W/m2降低到阴雨天的100 W/m2、从2 200 μmolm-2s-1降低到200 μmolm-2s-1(图2c),对应不同深层土壤温度也有所降低(图2d),比如,5 cm层土壤温度最大值从27.5℃降低到18.0℃,这些要素的变化也将改变不同组分土壤的呼吸速率。

图2 2020年5月21-31日SACOL站主要环境因子的连续变化情况
3.1.2 剪草后土壤呼吸速率的时间变化序列
剪草后5个不同气室土壤呼吸速率的连续变化情况如图3所示,1号气室的测量结果可以大致代表土壤微生物呼吸与微生物对干根际有机碳的分解;2号气室内的测量值可大致代表土壤微生物的呼吸;3~4号气室的观测值包含了活根系呼吸、土壤微生物呼吸及活根际有机碳的分解;而5号气室代表草地生态系统总的呼吸作用,包含根系和叶片冠层的呼吸、土壤微生物呼吸和根际有机碳的分解等。剪除地上植被冠层后的3号、4号气室之间的土壤呼吸速率差异较小(平均标准差为0.12 μmolm-2s-1)且变化规律非常一致。21-31日不同气室的土壤呼吸速率连续变化趋势较为一致,都是在每日的 12:00-14:00 出现最大值(3.30~5.0 μmolm-2s-1),而在日出前或夜间出现极小值(0.23~0.68 μmolm-2s-1),这与我国其他地区生长季草原土壤呼吸的日变化特征较为一致[8-12]。不同气室之间对比可得,5号茂盛草植被的土壤呼吸速率最大,只保留活根系的3号、4号次之,裸地最小,即意味着草地生态系统总呼吸>(土壤根系呼吸+微生物呼吸)>(土壤微生物呼吸+干根际分解)>土壤微生物呼吸。降水后(5月27-31日)5个气室的平均呼吸速率分别比降水前(21-22日)增加了0.37 μmolm-2s-1、0.39 μmolm-2s-1、0.66 μmolm-2s-1、0.79 μmolm-2s-1和 0.82 μmolm-2s-1。结果再次表明,当前期土壤干旱时,降雨过程引起不同土壤层水分的增加对茂盛植被的根系呼吸、叶片冠层呼吸和土壤微生物的种群数量、土壤的养分供给及分解活性具有显著的增强作用,对干枯植被和裸地的呼吸速率虽然有一定的促进作用,但影响较小。
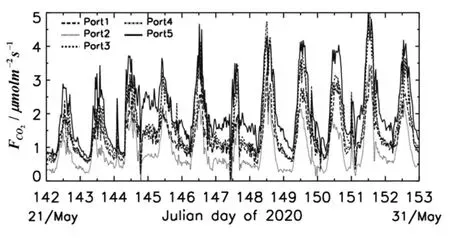
图3 2020年5月21-31日SACOL站各气室土壤呼吸速率的日变化
为更加深入地讨论降水对不同组分土壤呼吸的影响,我们选取5月21日和28日作为个例,并通过求差值的方式将不同组分初步区分出来,其中土壤微生物的呼吸可用2号气室的测量值大致代表,根系呼吸用3号气室与2号气室之间的差值来估算,叶片冠层呼吸用5号和3号气室之间的差值代表。如图4所示降水前(图4a)和降水后(图4c)土壤微生物的呼吸速率均为单峰型日变化,夜间时段(20:00-06:00)为较小值(0.20~0.40 μmolm-2s-1),从 07:00 开始迅速增加,到 12:00 达极大值,然后又逐渐减小,最低和最高值分别为0.20/0.25 μmolm-2s-1、1.35/2.10 μmolm-2s-1,对应出现在 06:00 时和12:00时。造成上述现象的原因是夜间土壤温度较低且光合作用较弱,使得土壤微生物活性也相对微弱,通常在凌晨出现最低值。而白天正午12:00太阳辐射较强、光合作用和土壤温度均较高,适宜的温度条件使土壤微生物呼吸作用达到极大值,但随着温度的进一步升高,超过微生物活动的温度临界值,此时土壤微生物活性受到抑制,呼吸作用逐渐减弱。茂盛根系和叶片冠层的呼吸速率均呈现“多峰型”日变化,其中,茂盛根系的极值出现的时间比土壤微生物呼吸的滞后2~3 h,即在降水前后最低和最高值分别出现在9:00/10:00时和14:00/17:00 时,对应为-0.25/0.20 μmolm-2s-1、1.0/1.80 μmolm-2s-1。此外,根系呼吸作用在夜间(22:00-10:00)较弱,变化范围在-0.25~0.50 μmolm-2s-1,而白天(11:00-21:00)较强,保持在 0.50~1.80 μmolm-2s-1。叶片冠层的呼吸作用主要受太阳辐射和光合作用的影响,其呼吸速率在夜间(20:00-08:00)很微弱(0.0~0.30 μmolm-2s-1),而在白天(09:00-19:00)变化较大(0.20~1.0 μmolm-2s-1)。最低和最高值分别为 0.07 μmolm-2s-1和 1.0 μmolm-2s-1,对应出现在08:00时和 12:00时。图4(c)中,5月 28日12:00-15:00的多云天气使得到达地面的太阳总辐射和光合有效辐射严重衰减,进而影响了植物的光合作用,因此冠层的呼吸无明显的极大值。总的来说,降水通过影响土壤水热条件、根系生长需水量及微生物活性,有效促进了根系和土壤微生物的呼吸作用,但对叶片冠层的呼吸影响较小。降水前后,根系、土壤微生物和叶片冠层呼吸极大值的增幅分别为83.2%、60.9%和26.0%。

图4 降水前后(a、c)根系呼吸(Port3-Port2)、土壤微生物呼吸(Port2)和叶片冠层呼吸(Port5-Port3)的动态日变化;降水前后(b、d)各组分占草地生态系统总呼吸(Port5)的比例
本文又将不同组分的呼吸速率除以草地生态系统总呼吸(Port5),用来定量研究各组分对总呼吸的相对贡献。图4中,降水前(图4b)和降水后(图4d)土壤微生物呼吸在总呼吸中的占比呈现较一致的日变化特征,都是夜间 20:00-06:00为较小值(18%~40%),白天 07:00-19:00为较大值(32%~78%),最低和最高值分别为30%/18%、65%/78%,分别出现在 23:00-24:00时和 10:00时,降水在一定程度上促进了土壤微生物呼吸对总呼吸的贡献。茂盛根系呼吸的占比在夜间(20:00-08:00)变化为 35%~80%,显著高于白天(09:00-19:00)的 10%~45%,极小和极大值分别出现在10:00时和23:00时,这与Li等[17]利用挖壕沟法分析SACOL站2008-2009年根系呼吸占比的14.5%~63.6%较为一致,直接证实了本文使用的估算方法具有一定的可靠性。而叶片冠层呼吸在土壤总呼吸中的占比波动变化较大,范围在0%~40%,其中夜间(22:00-06:00)保持较大的值(20%~36%)。晴天较强的光合作用能有效增强冠层呼吸的占比,而阴天条件下植被光合作用和土壤温度都有较大的减弱,使冠层呼吸的贡献显著降低。
3.1.3 降水前不同组分土壤呼吸速率与水热因子的关系
土壤温度和水分可通过改变植被根系、叶片和茎生理代谢和微生物的活性,影响土壤呼吸作用。如图5所示,干旱条件下(土壤含水量<0.05 m3m-3),在一定温度范围内(10~40℃)茂盛根系和土壤微生物呼吸与5 cm土壤温度均呈现正相关关系,即它们的呼吸速率会随着土壤温度的升高(或降低)而增加(或减小),采用线性或指数模型能较好地拟合土壤温度对其影响,见表1。土壤微生物的呼吸速率与土壤温度表现出明显的顺时针环状结构,即日出后随着温度的升高而增加,到中午微生物的呼吸速率达最大值(1.40 μmolm-2s-1),对应的土壤温度为24~26℃,午后随着土壤温度的继续上升,微生物的呼吸速率呈现出逐渐降低的趋势,这就是时滞现象。土壤微生物呼吸实际是一种酶促生化反应过程,微生物体内酶的活性随着温度变化而改变,当温度超过一定的生理阈值时,某些酶的活性可能降低,甚至失活,从而导致微生物呼吸作用的温度敏感性降低[23-24]。而叶片冠层与土壤温度的关系呈现较大的散点变化,从拟合结果来看,其受土壤温度的影响程度明显低于根系和微生物呼吸。

表1 2020年5月21日各组分的呼吸速率与5 cm土壤温度的拟合方程
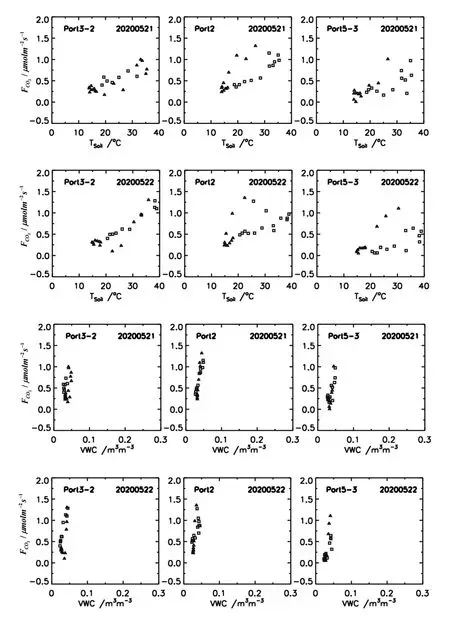
图5 干旱条件下不同组分的呼吸速率与土壤温度、含水量的关系(三角和正方形分别代表上午与下午时段)
3.1.4 降水后不同组分土壤呼吸速率与水热因子的关系
降水过程能增加不同深度土层的含水量,为植被的光合作用、叶片和根系的生理代谢、微生物的活性分解等提供必要的水分,同时降水的冲刷和淋溶作用可促进地上的生物量残体向地下输送,为土壤呼吸提供重要的基质[25]。如图6所示,降水后5 cm土壤含水量显著增加,变化范围在0.12~0.25 m3m-3,有效缓解了干旱土壤的状况。此时根系呼吸与5 cm土壤温度仍具有明显的线性关系,但散点较多,主要受土壤温度和含水量的共同影响。降水后,土壤微生物的呼吸速率与土壤温度的时滞效应更为明显。微生物酶活性的温度生理阈值由干旱时的26℃增加到湿润条件的28℃,即湿润土壤能提高微生物体内某些酶活性的温度阈值,5月28日上午和下午环状结构的指数拟合方程分别为Fco2=0.146×e0.11Tsoil和 Fco2=0.096×e0.086Tsoil,相关系数为 0.649 和 0.885,见表2。而降水后叶片冠层的碳呼吸速率与土壤温度无明显的线性关系。

图6 降水后不同组分的呼吸速率与土壤温度、含水量的关系(三角和正方形分别代表上午与下午时段)
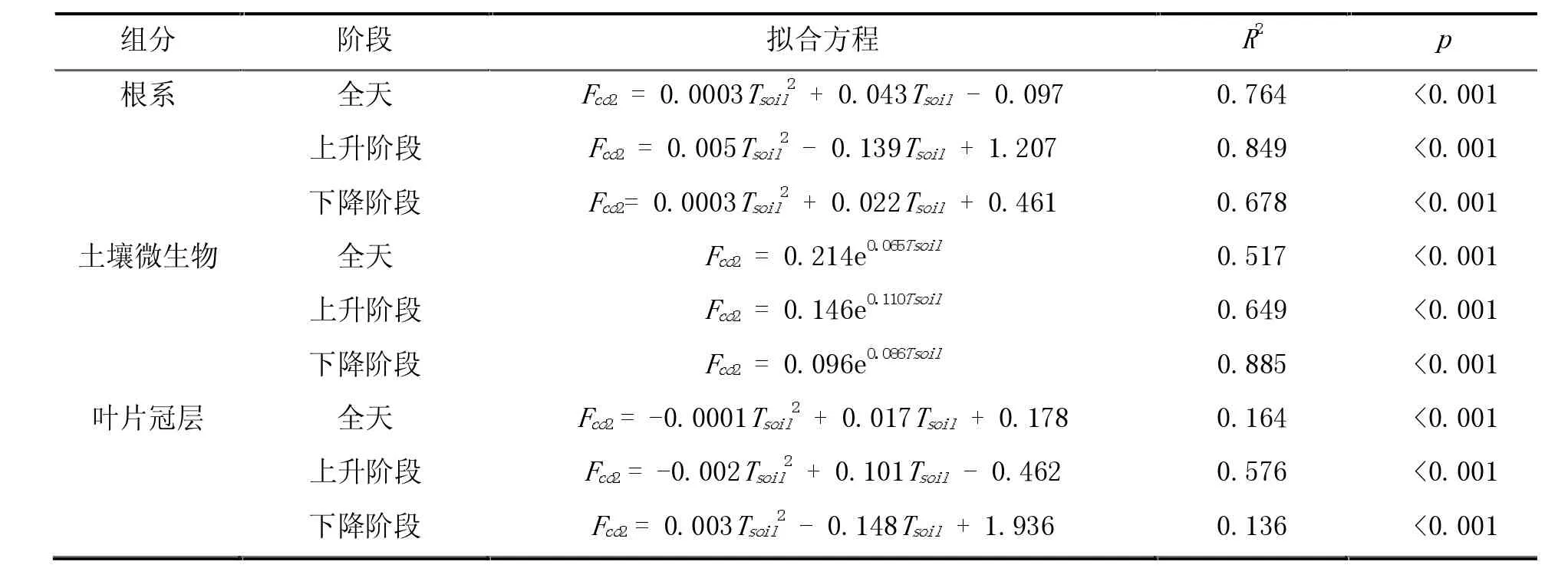
表2 2020年5月28日各组分呼吸速率与5 cm土壤温度的拟合方程
4 结论
(1)不同植被覆盖条件下土壤呼吸速率的时间变化序列一致性较好,都是在日出前或夜间出现极小值(0.23~0.68 μmolm-2s-1),而在 12:00-14:00 出现最大值(3.30~5.0 μmolm-2s-1),具体大小排列为:草地生态系统总呼吸>(土壤微生物呼吸+茂盛根系呼吸)>(土壤微生物呼吸+干根际分解)>土壤微生物呼吸。
(2)土壤微生物的呼吸速率呈明显的“单峰型”日变化特征,在夜间(20:00-06:00)为较小值,12:00 时达极大值。而叶片冠层和茂盛根系呼吸的日变化表现为:“多峰型”波动,前者最低和最高值分别为0.07 μmolm-2s-1和 1.0 μmolm-2s-1,出现在 08:00 时和 12:00 时,后者最值出现时间比土壤微生物滞后2~3 h。降水会有效促进土壤微生物、叶片冠层及根系呼吸的极大值,分别增加了60.9%、26.0%和83.2%。
(3)土壤微生物呼吸的贡献占比在夜间20:00-06:00为较小值,而白天07:00-19:00为较大值,最低和最高值分别为30%、78%,出现在06:00时和10:00时,降水能有效增加该占比的最高值;茂盛根系呼吸的占比在夜间(20:00-08:00)变化范围为 35%~80%,显著高于白天(09:00-19:00)的10%~45%,极小和极大值分别出现在10:00时和23:00时;叶片冠层呼吸的占比表现出较大的波动变化,变化范围在0%~40%,因此构建半干旱区草地生态系统的碳收支模型时,不能忽略叶片冠层的贡献。
(4)当土壤温度在10~40℃时,降水前后茂盛根系和土壤微生物呼吸与5 cm土壤温度呈显著正相关关系,降水会加强根系呼吸的“时滞效应”,而叶片冠层的呼吸速率与土壤温度并没有明显的相关关系。
猜你喜欢
农业工程学报(2022年12期)2022-09-09
上海农业学报(2022年4期)2022-09-06
中国农业气象(2022年8期)2022-08-24
作物杂志(2022年3期)2022-07-06
作物学报(2022年5期)2022-03-16
土壤(2022年1期)2022-03-16
汽车实用技术(2021年14期)2021-08-05
少儿科学周刊·少年版(2019年1期)2019-04-18
江苏农业科学(2017年21期)2017-12-13
中国新技术新产品(2017年23期)2017-11-14
