
GDF9、BMP15 基因在单、多羔黔北麻羊不同组织中的表达研究
2022-05-16 09:26:30韦仕南周志楠陈浩翰
中国畜牧杂志 2022年5期
韦仕南,周志楠,陈浩翰,陈 祥*
(1.贵州大学高原山地动物遗传育种与繁殖教育部重点实验室,贵州省动物遗传育种与繁殖重点实验室,贵州贵阳 550025;2.贵州大学动物科学学院,贵州贵阳 550025)
从动物遗传育种的角度探究如产奶量、产仔数、产肉率等经济性状基因的遗传学作用,可使人们在将来培育出更有利于人类生产的畜禽品种,从而造福人们。因此,与产羔数相关的主要调控基因的筛选一直是研究热点。近年来,已发现许多影响羊繁殖力的主效基因和候选基因,其中就包括生长分化因子9()、骨形态发生蛋白15()。基因是由卵母细胞分泌的一种生长因子,是家畜高繁殖力的候选基因,它调控着与卵丘扩张繁殖相关的几种关键的颗粒细胞酶,是卵巢中启动卵泡发育以及调节排卵率的必需生长因子之一。同时还参与调节动物的生殖性状。作为一个主要的影响产羔数性状的基因,在早期卵泡的生长和分化的调控作用方面人们已经投入了大量的研究。有研究发现在绵羊卵巢中高水平表达,蛋白质编码区全长约2.5kb,由2 个外显子和1 个内含子组成,可编码453 个氨基酸,能够参与影响早期卵泡的发育和分化。Dong 等研究发现,通过对小鼠和绵羊的基因功能失活或表达沉默,造成卵泡发育停滞,卵巢不能正常排卵,从而导致小鼠和绵羊繁殖机能障碍。有研究结果表明,基因可以促进原始卵泡向初级卵泡发育,缺失该基因则导致初级卵泡无法形成,导致雌性不孕。
与基因结构同源且功能类似,故又被称为。BMP15 蛋白是骨形态发生蛋白中的一员,BMP15 蛋白作为一种卵母细胞分泌因子(Oocyte-Secreted Factor,OSF),主要在雌性动物卵母细胞中表达并被分泌到胞外,通过结合到卵母细胞周围颗粒细胞/鞘细胞膜上的特异性受体行使生物学功能。绵羊的基因位于Χ 染色体上,CDS 区长度为1 182 bp,其中包括2 个外显子和1 个内含子。程俐芬等通过比较已知的基因产物的肽链,发现其在哺乳动物中高度保守,具有高度同源性。研究表明,能够以同源二聚体或与形成异源二聚体的方式参与调节卵泡发育和固醇类激素生成等生理过程。Otsuka 等人发现能通过抑制卵泡雌激素受体在颗粒细胞中的表达来抑制颗粒细胞分化。
大量研究表明,和基因在动物卵泡发育中具有协同作用,其共同作用于卵泡的早期发育。卵母细胞分泌因子和通过旁分泌/自分泌信号通路在原始卵泡发育、排卵过程、黄体形成、颗粒细胞增殖和卵母细胞成熟等过程中发挥重要作用。和之间因缺乏半胱氨酸残基而不能形成分子间二硫键,但是它们能够组成非共价键二聚体,并且共同表达时还能够形成异源二聚体。和能够帮助卵丘细胞合成胞内胆固醇,还可以协同促进卵丘细胞增殖和卵母细胞成熟。因此,研究和基因在黔北麻羊单/ 多羔组各组织中的表达规律,对黔北麻羊育种及提高经济性状具有重要的意义。
黔北麻羊又名麻羊,是贵州省地方优良山羊品种,主要产自贵州省北部,属皮肉兼用型品种,它对环境具有较强的适应性,有着耐粗饲、抗病力强、繁殖性能好、繁殖周期长、一年四季皆可发情等优点。近几年,与基因在绵羊、鸡、猪上已有大量研究,但2 种基因mRNA 表达谱在黔北麻羊上的研究却很少。本实验通过实时荧光定量PCR 分析初步探究基因和基因在单、多羔黔北麻羊子宫、垂体、输卵管、下丘脑和卵巢5 个组织样中的表达水平,为今后深入研究影响黔北麻羊繁殖力因素提供理论基础。
1 材料与方法
1.1 实验对象 随机收集相同饲养条件下单羔和多羔经产母羊(各3 只)的组织样本,参照贵州省地方《羊屠宰操作规程》(DB22/T 2740-2017)标准进行屠宰,用经高温灭菌消毒后的剪刀、镊子等器材采集适量且新鲜的子宫、输卵管、垂体、下丘脑和卵巢组织,用马克笔做好标记后放入低温液氮中保存以防样品腐坏,转存于-80℃冷冻室内,便于下一步实验的顺利进行。
1.2 主要仪器与试剂 实验主要仪器有实时荧光定量PCR仪(美国Thermo Fisher 公司CFΧ96 Real-Time System),旋涡混匀器(上海汉诺仪器有限公司ΧH-D),微型掌上离心机(北京东林昌盛生物科技有限公司DL200),制冰机(日本三洋电器有限公司SIM-F140),高速冷冻离心机(安玛西亚中国有限公司NANODROP 2000),以及购自美国BIO-RAD 公司的凝胶成像系统(C1000 Touch)、电泳仪(Universal Hood II)、超微量紫外分光光度计(PowerPacTM HV Power Supply)等。
TRIzol 试剂盒购自贵州绿盟英创生物科技有限公司,cDNA 第一链合成试剂盒购自北京康为世纪生物科技有限公司,0.5%TAE 缓冲液、液氮、实时荧光定量mix、氯仿、75%乙醇、琼脂糖、异丙醇均于贵州鼎国生物有限公司采购。
1.3 实验方法
1.3.1 RNA 提取 采用Trizol RNA 裂解法提取各组织总RNA,琼脂糖凝胶电泳检测其纯度和完整性,使用超微量分光光度计检测其浓度和纯度(D/D=1.8~2.0),检测结果显示所提取RNA 呈单峰、无杂峰,浓度较高,纯度较好,置于冷冻室保存可用于下一步反应的进行。
1.3.2 cDNA 第一链的合成 依照逆转录试剂盒说明书的步骤对各组织总RNA 逆转录合成cDNA 第一链。合成体系具体如下:dNTP Mix 2.5 mm Each 4 μL,Primer Mix 2 μL,5×RT Buffer 4 μL,RNA 模板1 μL,DTT(0.1 m)2 μL,HiFi Script 1 μL,RNase-Free Water 6 μL。用普通PCR 仪42℃孵育50 min,85℃下孵育5 min,最后用超微量紫外分光光度计进行PCR 产物的浓度和纯度检测,放入-20℃冰箱保存。cDNA 提取全程均在冰上进行。
1.3.3 引物的设计与合成 根据GenBank 上已经发布的山羊基因序列(登录号:EU888137.1)、基因序列(登录号:NM_001285708.1)利用Primer Premier 5.0 软件分别设计2 对引物,引物信息见表1。选取-为内参基因,引物交由重庆擎科生物科技有限公司合成。
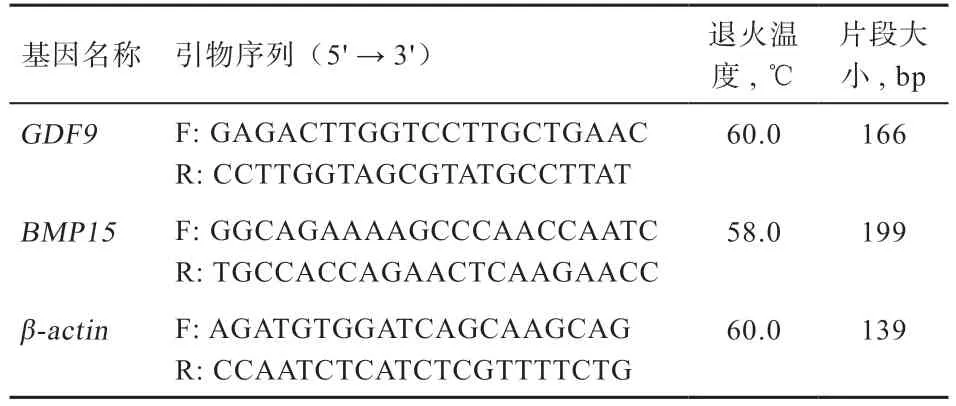
表1 引物序列信息
1.3.4 实时荧光定量PCR 条件优化及反应 以-基因为内参基因,对采集的单、多羔组黔北麻羊的5 个性腺组织、基因mRNA 进行实时荧光定量PCR 检测,溶解曲线由机器自动设置,每个样品各设3 个重复。荧光定量反应体系如下:(2×)SYBR Premix Ex Taq酶5 μL,上下游引物各0.5 μL,cDNA模板1 μL,dd HO 3 μL。荧光定量反应程序如下:95 ℃预变性1 min,95 ℃变性45 s,65 ℃退火15 s,72℃延伸30 s,39 个循环。在此过程中,对退火温度和引物浓度等客观可调节因素进行了探索,以寻找并选用最佳反应条件。
1.3.5 数据分析 根据获得的CT 值,通过2方法计算出的目标基因的相对表达量。数据用Excel 进行处理,并绘制相应的柱形图。采用SPSS 17.0 软件进行单因素方差分析,采用LSD 法进行差异显著性检验。结果以“平均数±标准差”表示,在0.01 和0.05 水平进行差异显著性分析。
2 结果与分析
2.1、基因普通PCR 扩增产物检测 由图1 可知,扩增片段与靶片段相同,长度分别为166 bp 和199 bp,清晰度高,特异性强,无拖带现象,没有发现引物的3'端间相互错配扩增现象,可直接用于下一步实验。
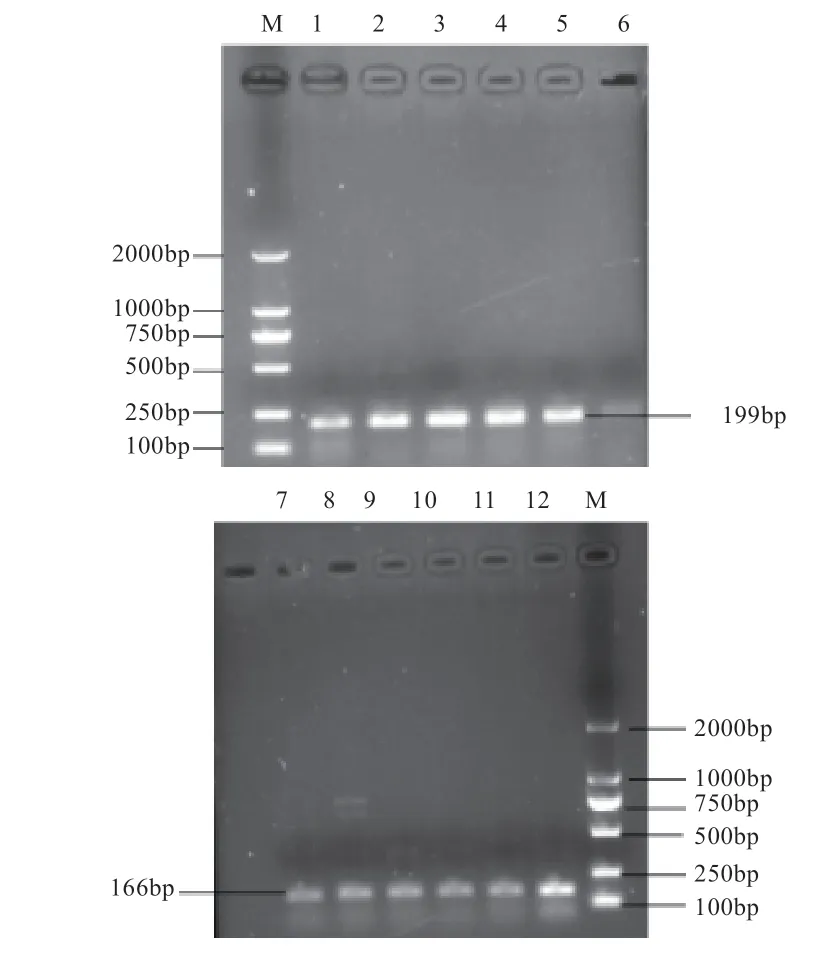
图1 BMP15、GDF9 基因PCR 扩增产物琼脂糖凝胶电泳图像
2.2 qPCR 产物特异性分析 通过对基因和基因进行qPCR 扩增获得的扩增曲线和溶解曲线如图2、图3 所示。可以看出,溶解曲线和扩增曲线都显示出单峰现象,扩增产物具有特异性,荧光反应明显,没有出现非特异性扩增产物和其他引物二聚体异峰。这表明黔北麻羊基因和基因的扩增结果是合理的,具有良好的代表性。说明定量检测结果精确可信,获得的数据可用于分析接下来的测试结果。

图2 BMP15 基因在黔北麻羊单、多羔组不同组织扩增曲线与溶解曲线
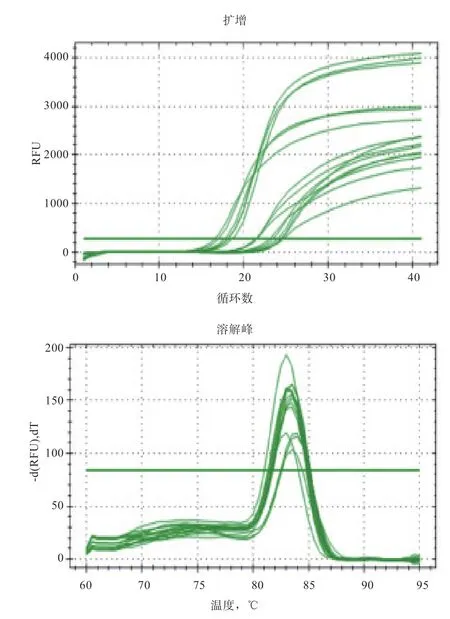
图3 GDF9 基因在黔北麻羊单、多羔组不同组织扩增曲线与溶解曲线
2.3、基因在单羔与多羔黔北麻羊各组织之间的相对表达分析 由图4 可知,基因在单、多羔黔北麻羊的性腺轴5 个组织中表达的高低顺序不相同。单羔组中基因在5 种组织中的表达量由多到少的顺序为:卵巢>垂体>输卵管>下丘脑>子宫,而且基因在卵巢组织中的相对表达极显著高于垂体、输卵管、下丘脑和子宫,其他的组织之间差异不明显;多羔组5 个组织中基因的相对表达顺序依次为:卵巢>垂体>下丘脑>输卵管>子宫。卵巢和垂体之间表达差异为极显著,且垂体极显著高于下丘脑、输卵管和子宫,下丘脑、子宫和输卵管之间的差异不显著。总体上单羔和多羔的表达趋势趋于一致。通过组间比较发现基因在多羔组的垂体相对表达量显著高于单羔组,在输卵管中单羔组的相对表达量显著高于多羔组,其余组织间则差异不显著。
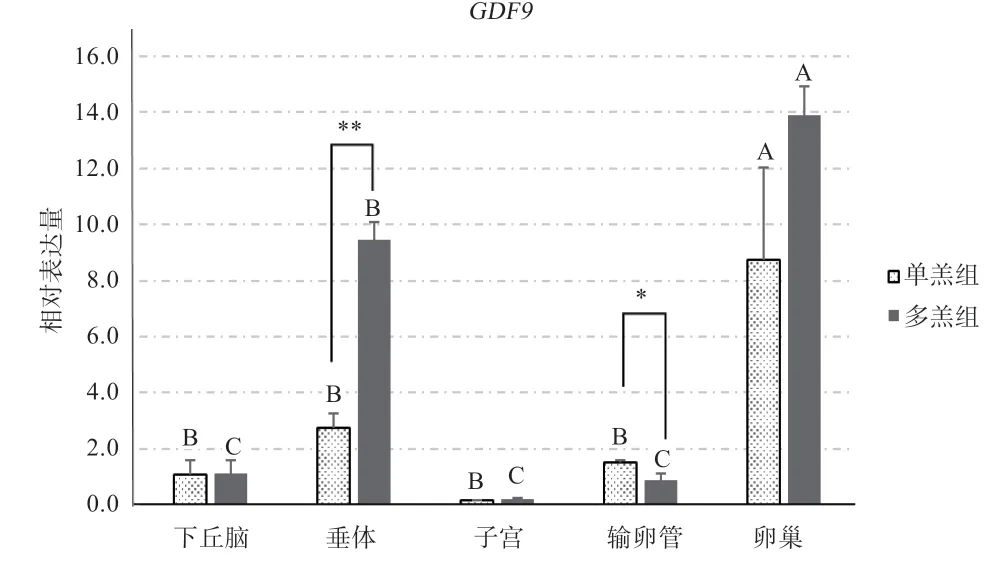
图4 GDF9 基因在黔北麻羊各组织mRNA 的相对表达量
由图5 可知,基因在单羔组和多羔组的5个组织中的相对表达趋势相近,均为在卵巢中表达最高。单羔组基因在5 个组织中的相对表达量顺序为:卵巢>输卵管>垂体>下丘脑>子宫,其中卵巢相对其他4 个组织表达差异为极显著,输卵管相对于子宫的表达差异为极显著,垂体相比较于子宫差异也达到了极显著水平,而输卵管相对于下丘脑的表达仅为显著,下丘脑和子宫之间的相对表达为不显著;基因在多羔组5 个组织中的相对表达量由高到低顺序为:卵巢>垂体>下丘脑>输卵管>子宫,基因在卵巢中的相对表达量极显著高于其他4 个组织,基因在垂体中的相对表达量显著高于下丘脑、输卵管和子宫,下丘脑、子宫和输卵管之间差异不显著。通过组间比较发现,基因在单羔组输卵管中的表达极显著高于多羔组,而在单羔组子宫中的相对表达量仅显著高于多羔组,其余组织间则差异不显著。
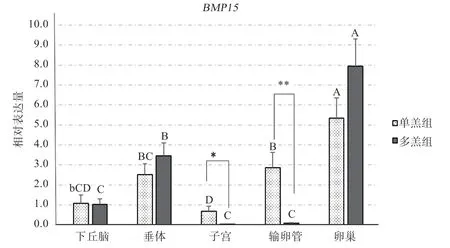
图5 BMP15 基因在黔北麻羊各组织mRNA 的相对表达量
3 讨 论
3.1 黔北麻羊基因在5 个性腺组织中的表达规律TGF-家族和BMP 信号通路是目前有关绵羊多羔性状候选基因的研究热点。作为TGF-家族重要成员之一的GDF9是哺乳动物和卵生动物卵巢内重要的调节因子。近年来,研究证实基因能够促进母羊前腔卵泡的生长成熟,并直接影响母羊的多羔生产性能。习欠云等人发现基因mRNA 在小鼠的性腺轴器官上皆有表达,且发现对的下调可使实验对象的生殖能力及生长发育有所改变,与本实验在黔北麻羊单、多羔组的性腺轴组织中均有表达的结果相同,但基因在多羔组的垂体和卵巢中的表达相比较于单羔组显示为上调,与上述发现有出入,这可能与基因在不同物种的性腺组织中的表达规律不同。贾超在其发现中证明了基因在绵羊3 个成长阶段的卵巢等组织中均有表达,并且不同年龄与组织之间都存在一定差异。其中2 月龄与本实验的结果一致,mRNA 表达量均为卵巢>输卵管>子宫,但与6 月龄和12 月龄的表达规律不符,故推测不同月龄母羊的各性腺组织表达规律不统一。这可能是由于该基因在卵泡早期发育时期较为活跃,随着月龄增长卵泡后期发育已趋于完善,故6 月龄和12 月龄的母羊基因在卵巢表达量降低。田蕾等人分析了各个组织和器官中基因的表达,结果表明在牛的卵巢和子宫中该基因高水平表达,但在输卵管、肾脏等器官表达水平偏低。这些结果均与本次实验结果一致。因此,基因对于卵泡生长发育的影响非常关键,但其对山羊产羔能力的调节仍有待深入探讨。
3.2 黔北麻羊基因在5 个性腺组织中的表达规律基因从发现和克隆至今已有近20 年的历史,关于其功能的研究已有大量报道。体外细胞培养实验、动物自发突变和基因敲除模型均表明,对哺乳动物的卵泡发育和卵巢正常功能的行使不可或缺。Dube 等人首先使用诺瑟杂交和原位杂交技术在卵巢卵母细胞中检测到了的特异性表达。刘世佳等人对湖羊的研究中发现,基因在雄性湖羊的心脏、消化系统、性腺器官、脂肪等组织中均有表达,以十二指肠表达最多。但是在雌性湖羊卵巢中基因的表达明显高于其他组织。董新龙等人发现在卵巢等性腺器官、内脏器官、肌肉和脂肪组织中都有不同程度的表达,且该基因在卵巢组织中的表达明显高于其他组织。本实验研究结果为基因在黔北麻羊单、多羔组中的卵巢组织中均显著高于其他组织,与上述研究结果一致。
有研究表明,和仅在哺乳动物卵巢中特定表达,但在之后的研究中发现它们在不同物种间表达的位置存在不同,表达量也并不统一。在鲤鱼、牦牛的研究中也发现或基因在卵巢中高表达。Fenwick 等发现通过与相互协同促进小鼠的原始卵泡生长,并拮抗对原始卵泡颗粒细胞的凋亡。汤继顺等研究表明小尾寒羊的2 个基因在卵巢的表达量均较高,并推测在小尾寒羊卵巢中高表达可能与产羔数存在正相关,而较低表达猜测可能与产羔数有一定的负相关,与本实验研究结果一致。这暗示基因可能作用于卵巢或抑制卵巢进一步分泌某种物质从而直接或间接影响动物的繁育与发育。
4 结 论
本研究显示、基因在单、多羔黔北麻羊5 个组织中均有表达,且表达趋势一致,2 个基因在卵巢中的表达水平均为最高,在子宫中的表达水平较低,两组织间均呈差异极显著。表明、基因与黔北麻羊的卵巢发育可能存在联系,卵巢可能是2 个基因的主要靶器官之一。
猜你喜欢
昆明医科大学学报(2020年12期)2021-01-26 00:44:02
中老年健康(2017年9期)2017-12-13 07:16:39
农产品市场周刊(2017年35期)2017-10-25 07:17:24
中成药(2017年6期)2017-06-13 07:30:34
海南医学(2016年8期)2016-06-08 05:43:00
中国民族医药杂志(2016年7期)2016-05-09 07:49:19
磁共振成像(2015年9期)2015-12-26 07:20:34
乡村地理(2015年3期)2015-11-10 06:33:38
中国煤层气(2015年6期)2015-08-22 03:25:14
中学科技(2014年11期)2014-12-25 07:27:11
