
水稻营养器官镉积累特性对稻米镉含量的影响
2022-05-05 12:28张昕张长波黄永春王常荣薛卫杰刘仲齐
农业环境科学学报 2022年4期
张昕,张长波,黄永春,王常荣,薛卫杰,刘仲齐
(农业农村部产地环境污染防控重点实验室,农业农村部环境保护科研监测所,天津 300191)
随着我国工业化和城镇化的快速发展,废弃物排放量显著增加,由此导致的土地重金属污染受到人们的广泛关注。镉是重金属污染土壤中最常见的元素之一,具有较高的可移动性,可以通过自由扩散或载体蛋白运输的方式进入植物根系,再通过质外体和共质体途径向木质部转运,最后转运至植物的可食用部分,通过食物链传递,危害人体健康。水稻是我国第一大粮食作物,也是全球食用量最高的谷物,因此探究稻米积累镉的特性、筛选低镉积累水稻基因型对于保证稻米品质安全具有重要意义。
水稻积累镉的能力受环境因素和基因型的共同调控。环境因素中土壤有效态镉含量、生长季节、水分管理措施等影响水稻根系对镉的吸收,在多个地点同时种植早稻和晚稻,发现晚稻各器官中的镉含量通常高于早稻。水稻根系对镉的阻控能力、叶片对镉的耐受程度以及镉在水稻各器官内的积累量和分配比例均存在基因型差异。在镉污染环境中,水稻根系通过根表铁锰膜拦截和细胞壁固定等方式阻控镉进入根系维管束的过程,地上部各器官会对镉进行层层固定和拦截,只有少数镉离子转运到稻米中。水稻开花以后,发育籽粒成为代谢中心,叶片和茎秆中储备的各种营养物质开始向籽粒转运,镉会随着锰、锌等营养元素一起转运至籽粒中。灌浆期间的物质流向及其生理调控机制较为复杂,发育籽粒的选择性吸收、营养器官中的矿质元素含量以及叶片蒸腾速率等因素都会影响镉向籽粒的转运,镉在籽粒中积累量易受水稻发育状态和自然因素的干扰。稻穗离体培养法常用来进行营养生理和抗逆生理的研究,是否可以快速鉴别稻穗各部位镉积累特性及其在水稻基因型间的差异,尚未见报道。以高镉和低镉积累品种为试验材料,将离体稻穗放置在镉胁迫强度相同的环境中进行镉转运特性的比较,可以最大限度地排除环境因素的干扰和下部器官镉含量的差异对上部器官镉吸收转运特性的影响,可用于探究穗下节和穗颈等器官对籽粒镉积累量的贡献率,比较籽粒基部维管束镉拦截特性在基因型间的差异。
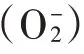
1 材料与方法
1.1 试验材料与地点
以“朝阳 1 号 B”(61)、“湘早籼 24”(X24)和“T 优705”(R705)3 种水稻基因型为试验材料,其中X24 和R705 为湖南省大面积推广的籼稻,X24 为低镉积累水稻,R705 为高镉积累水稻。61 是来自国家水稻核心资源库的地方品种。
离体稻穗和离体叶片试验在农业农村部环境保护科研监测所(简称环保所)人工气候室内进行,盆栽试验在环保所玻璃温室中进行。
1.2 培养与处理方法
挑选饱满的水稻种子,在体积分数为5%的HO溶液中浸泡30 min进行消毒,随后将种子冲洗干净并在育苗盘上进行萌发。待种子发芽后移入蛭石盘中进行自然生长,期间用四分之一Hoagland 营养液浇灌。水稻生长至20 cm 左右时,将离体稻穗和离体叶片试验所需要的水稻人工插秧至环保所温室内的无污染试验田中,基因型间设置空行;盆栽试验所需水稻插秧至土培盆中,每盆10 株,每个基因型设置4 个重复。盆栽试验的土壤采自于天津市某土壤污染区0~20 cm 的表层土,镉含量为 1.2 mg·kg,pH 为 7.5 左右;盆体积为8 L,每盆装土4.0 kg。生长期间浇灌保持水深2 cm 左右。水稻成熟后将其从土壤中连根拔出,洗净根表残留土壤,烘干至恒质量后分样,测定各器官镉含量。
于水稻开花期对无污染试验田中同日开花的稻穗进行标记,开花后15 d,将稻穗从倒一节下0.5 cm处剪断,去除叶鞘和叶片,进行离体稻穗试验。将稻穗放入含有不同镉浓度的十分之一Hoagland 营养液中培养5 d,处理液中的镉(CdCl·2.5HO)浓度设置为 5 个梯度,分别是 0.9、1.8、2.7、3.6 µmol·L和 4.5µmol·L,处理 2.5 d 时更换一次处理液。样品在75 ℃的恒温烘箱中烘干至恒质量,测定镉含量。

1.3 镉含量的测定
参照本实验室原有的方法,称取穗轴和籽粒干样各0.5 g 于消解管中,加7 mL MOS 级硝酸过夜,次日于ED54 电热消解仪上保持110 ℃进行消解,2.5 h 后冷却消煮管,加入1 mL 过氧化氢溶液,待反应平缓后,继续保持110 ℃消解1.5 h,随后将仪器升温至165 ℃赶酸,用去离子水冲洗消煮管,定容至25 mL后转移至塑料小白瓶保存消解液,用原子吸收仪(AAS ZEEnit 700)测定样品镉含量。
1.4 叶绿素含量的测定
称取离体叶片鲜样0.1 g于95%乙醇中研磨,过滤定容至25 mL,将95%乙醇设置为空白对照,用紫外分光光度计(UV-2010 型)分别在波长665 nm 和649 nm处用测定样品光密度OD值,计算叶绿素含量。
1.5 氧化损伤的测定
将镉溶液中处理过的新鲜叶片浸泡在10µmol·L的HDCFDA 溶液中,避光孵育2 h,染色后用灭菌后的去离子水清洗干净,将样品放置在20%的甘油中保存,防止叶片脱水卷曲。将叶片固定在载玻片上,在三目倒置荧光显微镜(测维光电LWD300-38LT)488 nm 处观察,绿色荧光的分布和强度代表叶肉细胞中HO的位置和数量。
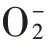
1.6 数据统计及分析
转移系数()=a器官镉积累量/b器官镉积累量
叶绿素 a(鲜质量)(mg·g)=(11.75×A665-2.35×A649)×0.1
叶绿素b(鲜质量)(mg·g)=(18.61×A649-3.96×A665)×0.1
叶绿素总量(鲜质量)(mg·g)=叶绿素 a 含量+叶绿素b含量
采用Excel 对数据进行处理并完成绘图,采用SPSS 22.0 软件进行单因素方差分析(One-way ANO⁃VA)、差异显著性检验(Duncan,<0.05)及相关性分析(Pearson)。
2 结果与分析
2.1 水稻各器官镉含量的基因型差异
水稻茎秆由节间和节构成,节上着生叶和芽。茎基部有7~13个节间不伸长,称为蘖节,是拦截镉的主要部位。在镉含量为 1.2 mg·kg、pH 为 7.5 左右的土壤中进行盆栽试验,发现水稻成熟期61 和X24 的籽粒、穗轴和茎基中的镉含量均显著低于R705,61 和X24 各器官镉含量无显著差异(图1A)。转移系数代表镉在器官间的转运能力,其数值越大表示镉的转运能力越强。镉从穗轴向籽粒的转运效率明显高于从茎基向穗轴的转运(图1B);在3 个基因型之间无显著差异,61 和 X24 的显著低于 R705。说明R705穗轴和籽粒基部拦截镉的能力较61和X24弱,其茎基拦截镉的贡献率更大。

图1 3个基因型水稻籽粒、穗轴、茎基镉含量以及转移系数在基因型间的差异Figure 1 Cadmium content in grains,rachises and basal stems among three genotypes and transfer factor of cadmium between different organs
2.2 离体稻穗镉的吸收动力学特征
水稻离体稻穗在含镉0.9~4.5µmol·L的处理液中培养5 d 后,计算镉的平均流速(µmol·g·h),发现随着处理液中镉浓度的升高,3 个基因型水稻籽粒中镉的吸收动力学特征均符合线性方程=a+b(图2A)。处理液中镉浓度每增加 1 µmol·L,X24 和R705 籽粒中的镉积累速率约增加0.44~0.49 µmol·g·h,而 61 籽粒中的镉积累速率只增加0.14 µmol·g·h。穗轴中的镉积累速率显著大于籽粒,其中X24 和R705 穗轴镉的吸收动力学特征符合线性方程,61 穗轴镉的吸收动力学特征符合指数方程(图2B)。当处理液中的镉浓度低于2.7µmol·L时,穗轴镉积累速率随着处理液中镉浓度提高而增加的幅度较小,积累速率的增长幅度在3 个基因型间差异不显著;当处理液中的镉浓度增加至3.6µmol·L后,61穗轴中的镉积累速率提升加快。处理液中的镉浓度相同时,61 穗轴镉积累速率始终高于X24 和R705。在0.9µmol·L镉胁迫下,3个基因型水稻籽粒吸收镉的最大速率为2.03~4.58 µmol·g·h,穗轴吸收镉的最大速率为11.54~15.62 µmol·g·h,在4.5 µmol·L镉胁迫下,3 个基因型水稻籽粒吸收镉的最大速率为2.53~6.16 µmol·g·h,穗轴吸收镉的最大速率为23.84~53.11 µmol·g·h。
离体稻穗籽粒中的镉含量在基因型间表现出显著差异。当处理中的镉浓度为0.9~4.5 µmol·L时,R705 籽粒中的镉含量高于61 和X24 籽粒中镉含量(图2C)。61籽粒中的镉含量在处理间的差异未达显著水平,X24 和R705 籽粒中的镉含量都随着处理液中镉浓度的增加而显著升高。当处理液的镉浓度为4.5 µmol·L时,籽粒中镉含量在基因型间的差异最为明显,61 和X24 籽粒中的镉含量分别为0.03 mg·kg和 0.06 mg·kg,而 R705 籽粒中的镉含量达到0.08 mg·kg;R705 的籽粒镉含量是 61 籽粒镉含量的2.44倍,是X24籽粒镉含量的1.29倍。
处理液浓度对离体稻穗穗轴镉含量的影响在基因型间表现出差异。61 穗轴中镉含量在各浓度镉胁迫下均高于X24 和R705 穗轴镉含量(图2D)。随处理液中镉浓度升高,61 和X24 穗轴中镉含量显著增加,而R705 穗轴中镉含量在低镉浓度胁迫下增加不显著,当处理液中镉浓度达到4.5µmol·L时镉含量显著增加。在0.9µmol·L镉胁迫下,穗轴镉含量在基因型间差异不明显,为0.16~0.21 mg·kg。在4.5 µmol·L镉胁迫下,61 穗轴镉含量则明显高于X24 和 R705 穗轴镉含量,达到 0.72 mg·kg,X24 和R705 穗轴镉含量差异不明显,镉含量为0.35 mg·kg左右。

图2 3个基因型离体稻穗籽粒和穗轴中镉的积累速率及镉含量Figure 2 Accumulation rate of cadmium in detached grains and rachises among three genotypes,contents of cadmium in detached grains and rachises among three genotypes
2.3 离体稻穗镉转运特性的基因型差异
穗轴是穗节和穗颈中各种离子向籽粒输送的末端出口。分别对3 个基因型离体稻穗籽粒和穗轴镉含量进行相关性分析,发现都符合线性方程=a+b,但斜率在基因型间有显著差异(图3A)。61的斜率显著小于X24 和R705,穗轴中镉含量每增加1.0 mg,61籽粒中镉含量增加0.01 mg,X24 籽粒中镉含量增加0.11 mg,R705籽粒中镉含量增加0.14 mg。说明61籽粒基部对镉的拦截能力显著大于X24 和R705,R705籽粒基部对镉的拦截能力最弱。
转移系数是判断穗轴向籽粒转运镉能力的重要依据,对5 个浓度镉胁迫下穗轴向籽粒的转移因子进行计算,发现随处理液中镉浓度增加,3 个基因型均逐渐减小,但转运能力在基因型间存在显著差异;在相同浓度镉胁迫下,61 和 X24 的均显著低于 R705,且 61 的显著低于 X24(图3B)。在 0.9 µmol·L镉胁迫下,61 的为0.13,X24 和 R705 的分别为 0.25 和 0.37,是61 的 1.92 倍和2.84 倍;当镉胁迫提高到4.5 µmol·L时,X24 和 R705 的下降到 0.17 和 0.25,但仍大于61在低镉胁迫时的。在4.5 µmol·L镉胁迫下,61的仅有0.05。

图3 籽粒和穗轴镉含量相关性及TF 籽粒/穗轴的基因型差异Figure 3 Relationship between cadmium content of grains and rachises,transfer factors of cadmium from rachises to grains among three genotypes
2.4 不同基因型离体叶片镉积累特性的差异
离体叶片浸泡在0.9~4.5µmol·L的镉溶液中培养3 d后,会随镉胁迫增强和时间推移而逐渐变黄(图4A)。3个基因型水稻在自然生长条件下叶片颜色较为均一,施加不同浓度镉胁迫后,X24 和61 在中等浓度的镉胁迫下就出现失绿现象,且失绿速率快,说明X24 和61 对镉胁迫更加敏感。X24 在镉胁迫达到13.5µmol·L时出现明显的失绿现象;61在镉胁迫达到 18.0 µmol·L时叶片黄化较为严重;R705 叶片在高镉胁迫下仍保持绿色。
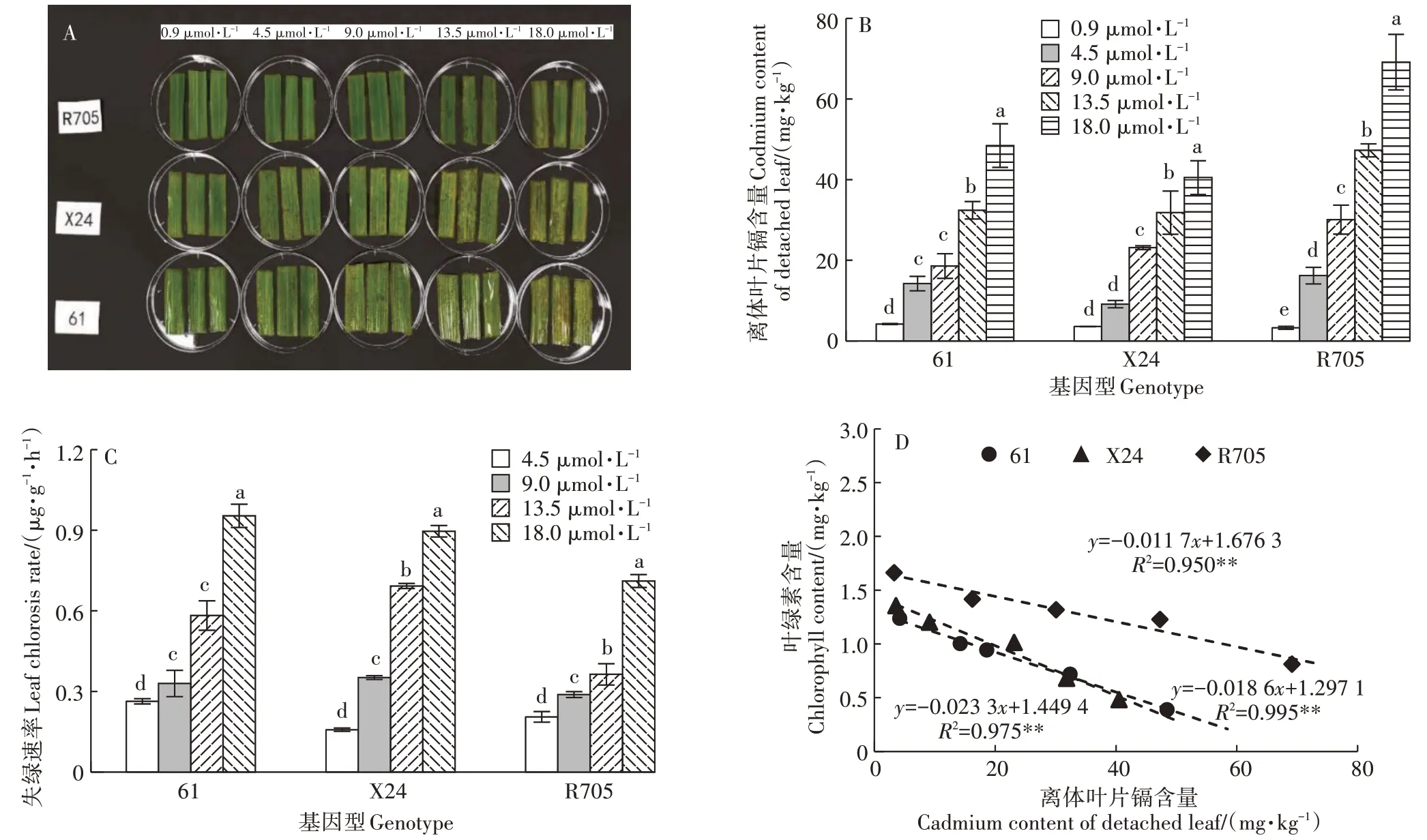
图4 三个基因型离体叶片失绿情况、镉含量、失绿速率及离体叶片镉含量和叶绿素含量相关性Figure 4 Etiolating of detached leaves,cadmium content and leaf chlorosis rate of detached leaves among three genotypes,relationship between cadmium content of detached leaves and chlorophyll content of detached leaves
离体叶片处理3 d 后测定镉积累量,发现离体叶片镉含量随处理液镉浓度升高而增加,并且在基因型间存在差异(图4B)。在0.9 µmol·L镉胁迫下,R705镉含量最低,但3 个基因型间无显著差异;当镉胁迫浓度在4.5~18.0 µmol·L之间时,R705镉含量始终高于61 和X24。基因型间镉含量差异在18.0 µmol·L镉胁迫时达到最大,R705 镉含量分别是61 和X24 的1.48 倍和1.70 倍。R705 离体叶片镉积累的速率比61和X24快,61和X24离体叶片在低镉浓度下镉积累速率缓慢且稳定。
叶片受到镉胁迫后,叶绿体被损坏并分解,叶片失绿状态反映其耐镉特性,计算离体叶片失绿速率,发现随镉胁迫增强,3 个基因型离体叶片失绿速率均显著加快,且随镉胁迫增强失绿速率加快幅度增大(图4C)。在镉胁迫浓度低于 9.0 µmol·L时,3 个基因型离体叶片失绿速率均较慢。当镉胁迫浓度达到13.5 µmol·L时,61 和 X24 失绿速率加快,分别是R705 的 1.60 倍和 1.90 倍。在 18.0 µmol·L镉胁迫时,R705失绿速率也显著加快。
对离体叶片中镉含量和叶绿素含量进行相关性分析,发现二者呈现负相关性,相关性符合线性方程=a+b(图4D)。斜率反映了叶绿素含量降低的幅度,R705 的斜率最小,其叶片每吸收1 mg 的镉,其叶绿素含量降低0.01 mg,61 和X24 的叶片每吸收1 mg的镉,叶绿素含量降低0.02 mg。说明61 和X24 叶片对镉胁迫的敏感性较强,R705 叶片对镉毒害的反应较为迟钝。
2.5 离体叶片耐镉能力的基因型差异


3 讨论
在相同的自然环境中,低镉积累品种的稻米镉含量显著低于高镉积累品种,但稻米镉含量的遗传稳定性很差,同一品种的稻米镉含量在不同年份和不同地点可以相差2~4倍。在盆栽试验中表现为低镉积累的品种,在田间试验中未必表现出低镉积累特性。然而,无论是田间试验还是盆栽试验,水稻穗轴和籽粒中的镉含量总是高度线性相关,灌浆期高镉积累品种营养器官的镉输出量总是显著高于低镉积累品种。为了排除环境因素的干扰,本研究将不同品种的稻穗及穗下节浸泡在相同浓度镉溶液中,比较稻米镉含量在水稻基因型间的遗传差异,发现离体稻穗籽粒和穗轴的镉含量均随镉浓度的增强而增加,高镉积累水稻T 优705 籽粒镉含量显著高于低镉积累品种湘早籼24和朝阳1号B籽粒镉含量,这说明在下部营养器官镉含量相同的情况下,高镉积累水稻的籽粒和穗轴吸收转运镉的能力更强。根据离体稻穗籽粒镉含量的高低,可以区分高镉和低镉积累品种,为快速筛选种质资源提供新的思路和依据。
水稻根系从环境中吸收镉离子后,会把大部分镉离子转化成难溶态镉固定在根系组织中,只有少数可溶态镉被转运到地上部,茎基部压缩的蘖节和地上伸长的4~5 个节间以及节和叶片会继续对镉进行层层拦截,将其储存在茎基、节和叶片中。水稻开花以后,茎叶中储存的部分镉离子被活化,和锰、锌等营养元素一起向穗轴和稻米中转运。镉和营养物质流经穗轴维管束到达籽粒基部,籽粒基部对转运而来的小分子化合物进行选择性吸收后运输至籽粒内部。本研究发现,镉从3 个基因型水稻的茎基向穗轴转运的比例()均小于,说明水稻茎基对镉的固定能力较强,能有效减少镉向上部器官转运。在离体稻穗试验中,高镉积累水稻T优 705 的显著高于低镉积累品种湘早籼 24和朝阳1 号B,说明T 优705 籽粒基部对镉的拦截作用较弱。由此可见,籽粒基部对镉具有识别和拦截作用,根据镉浓度在籽粒和穗轴间的比例可以快速判断发育籽粒镉积累特性在基因型间的差异,从而筛选籽粒基部对镉的拦截能力较强的低镉积累品种。

4 结论
(1)根据离体稻穗稻米和穗轴中的镉含量,可以快速准确地区分高镉积累和低镉积累基因型。离体稻穗在含镉营养液中灌浆时,稻米和穗轴镉含量显著线性相关,穗轴镉含量每增加1.0 mg,朝阳1 号B、湘早籼24 和T 优705 籽粒中的镉含量分别增加0.01、0.11 mg和0.14 mg。
(2)稻米镉含量由水稻各器官镉含量和镉在器官间的转运能力共同决定。低镉积累型水稻湘早籼24和朝阳1 号B 茎基、穗轴和籽粒中镉含量均显著低于高镉积累型水稻T 优705,且湘早籼24 和朝阳1 号B籽粒基部拦截镉的能力强于T优705。

猜你喜欢
新农业(2022年19期)2022-10-18
作物学报(2022年12期)2022-10-14
安徽农学通报(2022年6期)2022-04-07
作文·小学低年级(2021年10期)2021-01-25
作文评点报·小学三、四年级(2019年3期)2019-03-05
草原(2018年10期)2018-12-21
新课程·下旬(2018年8期)2018-11-10
小溪流(故事作文)(2017年10期)2017-11-09
科学种养(2017年6期)2017-06-13
中学生理科应试(2016年4期)2016-11-19
