
凋落物与根系混合分解及非加和效应研究进展
2022-04-27 12:56:36江鸿泓万晓华余再鹏李帅军许子君
福建农业科技 2022年2期
关键词:根系
江鸿泓 万晓华 余再鹏 李帅军 许子君

摘 要:凋落物分解是森林生态系统养分循环的关键生态学过程,在陆地生态系统中起着重要作用,是全球碳预算的重要组成部分。凋落物分解速率控制着养分循环和能量流动,进而调节大气中的碳排放、土壤有机质组成和养分有效性。凋落物的化学属性、环境气候、根系以及微生物群落组成等因素调控着凋落物分解速率。通过系统总结凋落物分解的基本理论、根与凋落物分解时根系的作用和调控机制以及凋落物混合分解非加和效应的研究进展,探讨了凋落物分解过程中多因素参与的复杂调控机制,并对未来的研究方向进行展望。
关键词:凋落物;混合分解;根系;非加和效应
中图分类号:S 714.3 文献标志码:A 文章编号:0253-2301(2022)02-0076-07
DOI: 10.13651/j.cnki.fjnykj.2022.02.014
Research Progress on the Mixed Decomposition and Non-additive Effect of Litters and Root Systems
JIANG Hong-hong1,2, WAN Xiao-hua1,2, YU Zai-peng1,2, LI Shuai-jun1,2, XU Zi-jun1,2
(1. State Key Laboratory Breeding Base of Humid Subtropical Mountain Ecology, Fujian Normal University, Fuzhou,
Fujian 350007, China; 2. School of Geographical Science, Fujian Normal University, Fuzhou, Fujian 350007, China)
Abstract: Litter decomposition was a key ecological process of nutrient cycling in the forest ecosystem, which played an important role in the terrestrial ecosystem and was an important part of the global carbon budget. Litter decomposition rate controlled the nutrient cycling and energy flow, and then regulated the carbon emission in the atmosphere, the composition of soil organic matters and the nutrient availability. The chemical properties of litters, environmental climate, root system and microbial community composition regulated the decomposition rate of litter. By systematically summarizing the basic theory of litter decomposition, the role and regulation mechanism of root system during the root and litter decomposition, and the research progress on the non-additive effects of the decomposition of mixed litters, the complex regulation mechanism involved by multiple factors in the process of litter decomposition was discussed, and the future research direction was prospected.
Key words: Litter; Mixed decomposition; Root system; Non-additive effect
凋落物,也可以稱为枯落物或有机碎屑,由生态系统内的地上植物组分枯死产生,最后归还到地表。凋落物维持着生态系统基本功能,同时也作为生态系统分解者和有机质能量来源[1]。凋落物分解是生物地球化学过程中的重要环节,也是生态系统内部和生态系统之间,以及生物圈和大气之间养分、碳和能量循环的基础[2-3]。1876年,Ebermayer[4]就指出,凋落物能够在森林生态系统养分循环方面起作用,而后许多关于凋落物的研究也证实了这个观点[5-6]。凋落物分解是森林生态系统物质循环重要载体,作为养分循环的一个基本的生态过程还在全球森林生产力维持方面发挥重大的作用[7-8]。对于森林生态系统而言,凋落物分解是不可或缺的、极其重要的一环,它既保证了生态系统养分循环和碳循环可以正常地运转[9],还能将分解后的碳通量归还到大气中,而后参与到全球碳预算中[10-11]。伴随着逐级不断分解的可利用性养分源源不断地归还回土壤本身,凋落物分解这一生态学过程对土壤肥力再形成有着积极贡献,能为植物提供再生长的养分,其影响着森林生态系统的物质循环和养分平衡[10]。
凋落物的化学属性(在生态学上简称为凋落物质量)决定了凋落物的相对可分解性[12]。作为影响凋落物分解快慢最主要的因子,凋落物质量与微生物分解者之间的相互作用是凋落物分解过程的一个及其重要的调控因子,在特定的区域范围的条件下发挥着至关重要的作用。分解过程伊始,分解速率主要受环境气候和自身化学性质的影响,其中温度和湿度起主导作用;此外,凋落物各个初始化学性质(如C/N, N/P, lignin/N)同样和分解速率息息相关[10,13];在分解初期凋落物的初始化学组成及微生物群落组成都对分解速率起调控作用,而在分解后期后者则起主导作用,前者对于分解的影响远不及初期[10,14]。
凋落物分解过程中,干重损失和养分释放两个方面对生态系统过程有着较大意义[15-18]。一般来说在生态系统中,地上植被的种类越丰富,对其自身的分解过程、分解生境以及分解者群体也会产生重要影响。森林凋落物的归还、分解和养分释放对森林自养功能的维护、森林生态系统土壤肥力和生产力的提高以及生态功能的发挥起着不可替代的作用[19]。本研究对凋落物分解的基本理论、根与凋落物分解时根系的作用和调控机制以及凋落物混合分解研究进行系统的总结,为凋落物分解的机制研究提供借鉴。
1 凋落物分解的影响因素
影响凋落物分解的因素有很多,主要包括环境因素、气候因素、凋落物性质以及生物因素等。在全球尺度上,对凋落物的分解起着控制作用的主要是气候因素;从区域小尺度来看,凋落物分解同时受到自身质量如木质素(Lignin)、C、N、P含量、Lignin/N、C/N以及土壤养分状况、土壤动物和微生物的影响。处于不同环境、科属不同本身具有差异的凋落物、分解过程中不同阶段,凋落物分解速率都不相同,对分解速率的影响程度也具有较大差别[20]。在凋落物分解的过程中,作为分解者之一的微生物,它们通过产生酶的方式,破碎化难分解的有机底物而后获得能量,在养分富集的地方构建群落;而胞外酶活性也与凋落物的分解速率和养分礦化速率显著相关[21]。因此,酶的活性升高后,作为凋落物和土壤有机质分解过程的关键催化剂与调控因子,酶使分解过程变得更加快速,释放养分元素的速率也会提高相应地提高。当处于分解的不同阶段时,因为凋落物自身所处的分解微环境变化大,周围土壤微生物群落组差异性大、变化迅速,使得分解速率的差异化明显。Marschner等[22]研究了小麦残余物的分解,他们认为在分解的头2周,由于微生物更容易竞争到水溶性物质而获得能量,从而导致其群落变化很大;之后,由于水溶性物质尽数被消耗,微生物群落变化相对缓慢许多。Voriskova等[23]研究了温带森林凋落物分解24个月期间真菌群落的动态变化,结果显示大多数丰度较高的真菌群落都只暂时主导着分解底物,分解过程中真菌类群的变化同样也很快,说明分解过程是一个以多种多样的真菌类群为媒介的复杂生态学过程。
2 凋落物与根系的混合分解
植物根系不仅生长在矿质土层里,还会长入处于不同分解阶段的凋落物层中;不难发现,根系长入凋落物层在森林生态系统很普遍[24-25],凋落物分解过程中在自然状态下是有根系长入的,即凋落物分解是与根系的混合分解。根本身就具有向肥性和向水性的根系在土壤中总是趋向着水分、养分富集的斑块生长[26-27];而表层凋落物更容易受温度、水分、光照等环境因素的影响,且分解矿化过程后更多是微生物利用了这部分的养分用以形成群落[28],因此,原本就匮乏的可利用性养分更是少之又少,故不太适宜根系生长[29]。
由轻微破碎的凋落物层和严重破碎的腐殖质层组成的中下层凋落物,因其更强的保水保湿能力,和更高可利用养分浓度的关系,比起位于土壤层和表层凋落物,根系会取向选择生长进这部分凋落物里;故根系在凋落物层中的生长呈现不均匀分布的状态[30]。此外,由于凋落物层根系的不均匀分布,它具有的细根生物量也与凋落物量直接相关。
2.1 根系对分解过程中干重损失和养分释放的不同影响
目前,根系在凋落物层生长对凋落物分解(干重损失)和养分释放的研究结果不一。例如,通过布设在亚马逊流域的野外试验观测到,根系对凋落物的干重损失和N、P等养分元素的释放速率均无显著影响,但进入凋落物层生长的根系仅仅增加Ca、Mg、Mn、Zn等金属元素的释放速率[31]。然而,Wang等[32]在亚热带湿地松和枫香林中的研究发现了截然不同的试验结果,即相对于低质量(高C/N比和C/P比)的凋落物而言,根系对高质量凋落物起了更多作用。细根的存在能够加快高质量凋落物分解且会使凋落物量降低,同时研究发现凋落物的O、C、N、P等养分元素浓度也降低。此外,在凋落物分解晚期,细根在凋落物中的增长更快,发挥的作用也更大[33-34]。总之,根系在凋落物分解的作用是一个动态的过程,而且受多种因素影响。
2.2 根系通过根际激发效应来调控凋落物分解过程
根际激发效应(Rhizosphere priming effect)是指植物根系通过刺激微生物活性来诱导有机质矿化的过程。根系生长通过根际激发效应影响凋落物的分解,主要从两个方面体现:即根系凋落物(死根)和根系分泌物对于分解过程的调控。根系中可利用性碳是微生物分解旧有机质的主要能源[35],因凋落物中有根系的生长,根系存在有激发效应进而调控分解过程。Fontaine等[36]发现,因土壤中缺乏可利用性有机质,土壤微生物的活动受到限制,而土壤有机质的分解随之也被抑制;但含较高能量的糖、淀粉等非结构性C源却能够刺激微生物群落地生长扩张,从而加快了土壤有机质的分解,它们则更多地存在于植物枝叶、根系及凋落物中。根系死亡后作为高能量C源, 刺激旧C源分解,这一点可以通过土壤有机质分解过程中的激发效应得到间接支持。在植物生长过程中,根系不仅从环境中摄取养分和水分,同时根系释放大量的化学物质,加上根组织脱落物一起统称为根系分泌物。对于根系分泌物而言,它们主要是分子类化合物。作为根际激发效应的主要贡献者,根系分泌物在土壤生态和植物养分循环方面有着无与伦比的意义,它不仅作为能量底物(即作为C源)给聚集在根周围的微生物利用,同时还刺激了微生物活性使其自身分泌胞外酶用来使有机质分解加快[37-38]。根际激发效应取决于底物的可利用性养分的浓度是根际激发效应的关键因素,因为对于根系分泌物还是根本身,初始的分解速率很大程度能够说明分解进程。有关土壤呼吸的研究也表明,土壤中C的呼吸显著增强植物根系的根际效应[39]。众多研究能够得出结论,即激发效应的发生主要取决于土壤养分可利用性,进一步说,土壤可利用性养分的有效性与根际激发效应有着千丝万缕的联系。
3 多物种凋落物的混合分解的影响因素
在能够产生凋落物的大多数生态系统中,不论是何种恢复模式,凋落物都以混合物的形式分解。通过多个物种凋落物混合分解与单一物种分解进行对比试验研究发现,多个物种的凋落物混合分解速率往往与混合物组分的预估单一平均分解速率有着较大的差异,凋落物在分解过程中产生了非加和效应(Non-additive effect),即多个物种凋落物混合有可能会促进分解,也有可能会延缓分解[40-42]。如果混合凋落物的实际分解速率等同于它各组分期望分解速率值则称之为加和效应;当估计值与实际值有差距时,则称之为非加和效应,即混合效应。混合凋落物的期望分解速率是根据单种组分凋落物分解速率与质量比例的加权平均数来计算
[43],广泛存在于凋落物分解过程中的混合效应会给全球分解模型中C通量的预测带来误差[44]。非加和效应中又有协同效应和拮抗效应,当混合凋落物的实际分解速率高于期望的分解速率为协同效应,反之为拮抗效应[45]。
3.1 多样性对凋落物混合分解的影响
物种组成多样性比物种丰富度能够更好地解释物种多样性对凋落物分解的影响[46-47]。物种多样性不只是物种数量的简单增加,因为物种数量可能无法捕获功能性生物多样性信息。例如,两个橡树种组合与一个橡树和一个松树种组合相比,树种数量相同,但是后者的系統发育指标被认为是具有更丰富的生态学信息[48]。生态位互补假说(Niche complementarity hypothesis)认为,物种组成的功能多样性越高越有利于提升生态系统功能[49]。与凋落物分解过程相关的植物群落功能特性,通常有叶片物候(常绿与落叶相比)、叶片类型(针叶与阔叶相比)、比叶面积(Specific leaf area, SLA)、叶片干物质含量(Leaf dry matter content, LDMC)、叶片 C/N 比等。例如,与常绿树种相比,落叶树种的凋落叶通常分解较快[50],原因有:第一,落叶树种的叶片具有较高的氮含量、较低的 C/N,而针叶树种叶片通常具有较高的C/N比和木质素含量;第二,落叶树种林分环境下土壤含水量较高,为分解者群落提供了较为适宜的环境条件[51-52]。而与落叶阔叶树种相比,常绿针叶树种一般具有较高的叶面积指数(Leaf area index,单位土地面积上植物叶片总面积占土地面积的倍数)和大量宿存叶片,降雨截留量较高,导致地表比较干燥。比叶面积,是叶片表面积与叶片干重之比,常与叶片养分含量、光合速率呈正相关。比叶面积还被认为是可以指示植物的水分获取和保留能力,与凋落叶持水率呈正相关[53]。此外,叶片干物质含量是叶片干重与饱和鲜重之比,叶片干物质含量高,难分解物质含量多,养分含量低,光合速率低,一般与凋落物分解速率具有负相关关系
[54]。因此,植物群落组成多样性很大程度上会通过这些功能特性指标对分解产生影响(图1)。
3.2 凋落物混合分解的非加和效应理论
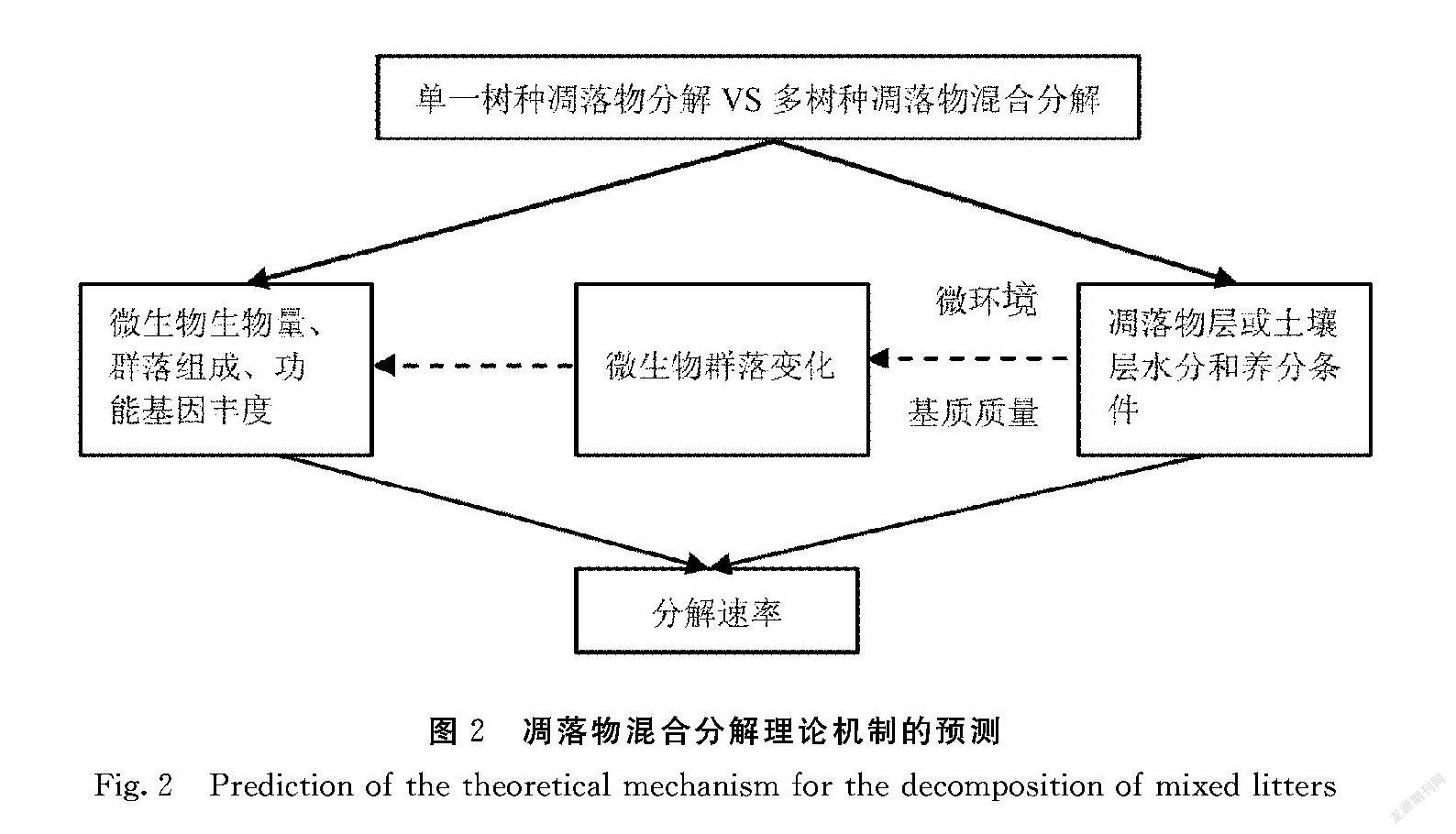
将不同物种的凋落物组合在一起会产生物理和化学功能特征更为多样的凋落物,导致其由于营养物转移而更快地分解,即协同效应[55],或为土壤碎屑和微生物提供补充养分[56]。实际上,在一些情况下,凋落物混合的非加性效应在功能多样性增加的情况下导致更快的分解(图2)。
目前,有4种理论可以解释在各个试验中观察到的凋落物混合物的非加性[57]。这些理论源于分解者受到食物质量和微生境结构差异的影响。养分转移(NT)理论表明,通过主动微生物转移或被动淋滤,优质凋落物种的养分转移到劣质凋落物种。特定化合物效应(ESC)理论不同于第1种理论,它侧重于突出刺激或抑制分解者群落的凋落物种特异性化合物对于分解过程的影响。第3种,不同营养级的相互作用理论(IATL),是基于观察结果,即当凋落物化学质量从单一栽培到混合凋落物时,土壤动物的取食率和食物选择也会发生变化。最后一个理论——改善微环境条件(IMC)理论与IATL理论相联系,该理论指出,更多样化的凋落物混合物改善了微气候条件,从而改善了分解者的栖息地和资源可用性。IMC理论起源于Wardle等[2,58]研究表明,羽毛苔藓能够促进凋落物中共存(即相邻)物种的分解,而水分是植物凋落物分解的控制因素之一,羽毛藓的这种效应被认为是由于其增强凋落物层水分保持能力造成的。总的来说,凋落物混合分解的非加和效应很难用现有的单一理论来分析和解释。
4 展望
森林凋落物的归还、分解和养分释放参与了森林生态系统养分循环和能量流动,在维护森林生态系统功能、提高土壤肥力和生产力以及生态系统功能的正常发挥等几个生态过程中同样无法替代。
根系与凋落物的混合分解中,根系通常会选择进入中下层凋落物生长。凋落物层中根系一方面使凋落物养分释放速率提高,另一方面因为存在使得凋落物层所处的分解环境都在发生变化,引起微生物类群不同程度的响应,进而又影响凋落物分解。当不同凋落物混合在一起时,凋落物的混合分解具有混合分解效应,在大多数情况下,凋落物混合物的分解速率是无法与加性预测相匹配的,换言之,混合分解效应即为非加和效应。但深究其机制,凋落物的质量损失、养分固定和转移以及分解者所处微环境的变化机制十分复杂。在这种加性分解的情况下,功能性状值而不是这些性状中物种间的差异性,似乎可以预测混合物中的分解模式[59]。现有证据表明,整个群落级凋落物的分解对多样性(特别是物种丰富度)的响应在不同的研究系统中不一致。
最近的研究表明,树木混交对土壤微生物并没有直接影响,但对地上凋落物和吸收根的性状以及土壤参数有间接影响[60];叶凋落物物种组成本身对分解速率具有显著影响,而不是凋落物物种丰富度,土壤性质对凋落物质量损失的影响在前期较强,而凋落物物种组成在后期更为重要[61]。尽管大量的植物凋落物研究表明了混合效应、对多样性如何影响凋落物分解的研究进行了全面的综合研究[62],但关键问题仍然存在,包括多样性在何时、何地和以怎么样的方式发挥其作用[63]。在凋落物组成与植物群落组成相匹配的情况下,植物群落多样性是否促进分解,还存在很大的不确定性[64]。植物群落多样性如何通过凋落物多样性和分解微环境影响分解过程,且在其影响过程中的关键机制是什么?还有待于进一步分析和验证。
参考文献:
[1]李宜浓, 周晓梅, 张乃莉, 等.陆地生态系统混合凋落物分解研究进展[J].生态学报, 2016, 36(16):4977-4987.
[2]WARDLE D A, NILSSON M C, ZACKRISSON O, et al.Determinants of litter mixing effects in a Swedish boreal forest[J].Soil Biology & Biochemistry, 2003, 35(6):827-835.
[3]EBERMAYER E.Die GesammteLehre der WaldstreumitRücksicht auf die ChemischeStatik des Waldbaues[M].Berlin:Springer,1876.
[4]严海元, 辜夕容, 申鸿.森林凋落物的微生物分解[J].生态学杂志, 2010, 29(9):1827-1835.
[5]宋影, 严海元, 辜夕容, 等.中亚热带马尾松林凋落物分解过程中的微生物与酶活性动态[J].环境科学, 2014, 35(3):1151-1158.
[6]GE X G, ZENG L X, XIAO W F, et al.Effect of litter substrate quality and soil nutrients on forest litter decomposition: A review[J].ActaEcologicaSinica, 2013, 33(2):102-108.
[7]BRAY J R, GORHAM E.Litter production in forests of the world[J].Advances in Ecological Research, 1964,2:101-107.
[8]VITOUSEK P M.Litterfall, Nutrient Cycling, and Nutrient Limitation in Tropical Forests[J].Ecology, 1984, 65(1):285-298.
[9]WIEDER W R, CLEVELAND C C, TOWNSEND A R.Controls over leaf litter decomposition in wet tropical forests[J].Ecology, 2009, 90(12):3333-3341.
[10]熊勇,许光勤,吴兰.混合凋落物分解非加和性效应研究进展[J].环境科学与技术, 2012, 35(9):56- 60.
[11]HOBBIE S E, EDDY W C, BUYARSKI C R, et al.Response of decomposing litter and its microbial community to multiple forms of nitrogen enrichment[J].Ecological Monographs, 2012, 82(3):389-405.
[12]SWIFT M J, HEAL O W, ANDERSON J M.Decomposition in Terrestrial Ecosystems[M].London: Blackwell Scientific Publications, 1979:372.
[13]MARIE-MADELEINE C, BOTTNER P, BERG B.Litter decomposition, climate and litter quality[J].Tree, 1995, 10:63-66.
[14]BRAY, SARAH R, KITAJIMA, et al.Temporal dynamics of microbial communities on decomposing leaf litter of 10 plant species in relation to decomposition rate[J].Soil Biology and Biochemistry, 2012, 49:30-37.
[15]BERG B, MCCLAUGHERTY C.Plant Litter: Decomposition, Humus Formation and Carbon Sequestration, seconded[M].Berlin, Heidelberg,2008.
[16]FRESCHET, GRGOIRE T, AERTS, et al.Multiple mechanisms for trait effects on litter decomposition: moving beyond home-field advantage with a new hypothesis[J].Journal of Ecology, 2012, 100:619-630.
[17]梁藝凡.亚热带人工林凋落物层细根对凋落叶分解的影响及机制[D].福州:福建师范大学, 2019.
[18]MANZONI S, JACKSON R B, TROFYMOW JA, et al.The global stoichiometry of litter nitrogen mineralization[J].Science, 2008, 321:684-686.
[19]郭剑芬, 杨玉盛, 陈光水, 等.森林凋落物分解研究进展[J].林业科学, 2006, 42(4): 93-100.
[20]SALAMANCA E F, KANEKO N, KATAGIRI S, et al.Nutrient dynamics and lignocellulose degradation in decomposing Quercusserrata leaf litter[J].Ecological Research, 1998, 13(2):199-210.
[21]HERNNDEZ D L, HOBBIES.The effects of substrate composition, quantity, and diversity on microbial activity[J].Plant and Soil, 2010, 335(1-2):397-411.
[22]SCHMIDT M W, TORN M S.Persistence of soil organic matter as an ecosystem property[J].Nature,2011, 478(7367):49-56.
[23]VORISKOVA J, BALDRIAN P.Fungal community on decomposing leaf litter undergoes rapid successional changes[J].Isme Journal, 2013, 7(3):477-486.
[24]ACHAT D, BAKKER M, TRICHET P.Rooting patterns and fine root biomass of Pinuspinaster assessed by trench wall and core methods[J].Journal of Forest Research, 2008, 13:165-175.
[25]HODGE A, HELGASON T, FITTER A H.Nutritional ecology of arbuscularmycorrhizalfungi[J].Fungal Ecology, 2010, 3: 267-273.
[26]CROFT S A, HODGE A, PITCHFORD J W.Optimal root proliferation strategies: the roles of nutrient heterogeneity, competition and mycorrhizalnetworks[J].Plant and Soil, 2009, 351:191-206.
[27]MOMMER L, VAN RUIJVEN J, JANSEN C, et al.Interactive effects of nutrient heterogeneity and competition: implications for root foraging theory? [J].Functional Ecology, 2012, 26:66-73.
[28]许子君, 万晓华, 梁艺凡,等.根系在凋落物层生长对凋落叶分解及酶活性的影响[J].应用生态学报, 2021, 32(1):31-38.
[29]FUJIMAKI R, MCGONIGLE T P, TAKEDA H.Soil micro-habitat effects on fine roots of ChamaecyparisobtusaEndl: a field experiment using root ingrowth cores[J].Plant and Soil, 2004, 266(1):325-332.
[30]王微, 胡凱, 党成强, 等.凋落物分解与细根生长的相互作用[J].林业科学, 2016, 52(4):100-109.
[31]LUIZO R C, LUIZO F, PROCTOR J.Fine root growth and nutrient release in decomposing leaf litter in three contrasting vegetation types in central Amazonia[J].Plant Ecology, 2007, 192:225-236.
[32]WANG F C, FANG X M, DING Z Q, et al.Effects of understory plant root growth into the litter layer on the leaf litter decomposition of two woody species in a subtropical forest[J].Forest Ecology and Management, 2016, 364:39-45.
[33]COTRUFO M F.Quantity of standing litter: a driving factor of root dynamics[J].Plant and Soil, 2006, 281:1-3.
[34]马承恩, 孔德良, 陈正侠, 等.根系在凋落物层中的生长及其对凋落物分解的影响[J].植物生态学报, 2012, 36:1197-1204.
[35]KUZYAKOV Y.Priming effects: interactions between living and dead organic matter[J].Soil Biology & Biochemistry, 2010, 42:1363-1371.
[36]FONTAINE S, BARDOUX G, ABBADIEL,et al.Carbon input to soil may decrease soil carbon content[J].Ecology Letters, 2004, 7:314-320.
[37]DIJKSTRA F A, WEIXIN C.Interactions between soil and tree roots accelerate long-term soil carbon decomposition[J].Ecology Letters, 2007, 10:1046-1053.
[38]BERTIN C, YANG X H, WESTON L A.The role of root exudates and allelochemicals in the rhizosphere[J].Plant and Soil, 2003, 256(1):67-83.
[39]FU S L, CHENG W X, SUSFALK R.Rhizosphere respiration varies with plant species and phenology: a greenhouse pot experiment[J].Plant and Soil, 2002, 239:133-140.
[40]GARTNER T B,CARDON Z G.Decomposition dynamics in mixed-species leaf litter[J].Oikos, 2004, 104(2):230-246.
[41]MORI A S, CORNELISSEN J H, FUJII S, et al.A meta-analysis on decomposition quantifies afterlife effects of plant diversity as a global change driver[J].Nature Communications, 2020, 11:4547.
[42]PORRE R J, VAN DER WERF W, DE DEYN GB, et al.Is litter decomposition enhanced in species mixtures? A meta-analysis[J].Soil Biology & Biochemistry, 2020, 145:107791.
[43]BADEJO M A, TIAN G, BADEJO M A, et al.Abundance of soil mites under four agroforestry tree species with contrasting litter quality[J].Biology & Fertility of Soils, 1999, 30(1-2):107-112.
[44]MEENTEMEYERV.Macroclimate and Lignin Control of Litter Decomposition Rates[J].Ecology, 1978, 59(3):465-472.
[45]HOORENS B, AERTS R, STROETENGA M.Does initial litter chemistry explain litter mixture effects on decomposition?[J].Oecologia, 2003, 137(4):578-586.
[46]BARANTAL S, SCHIMANN H, FROMIN N, et al.C, N and P fertilization in an Amazonian rainforest supports stoichiometric dissimilarity as a driver of litter diversity effects on decomposition[J].Proceedings of the Royal Society B: Biological Sciences, 2014, 281:20141682.
[47]LECERF A, MARIE G, KOMINOSKI J S, et al.Incubation time, functional litter diversity, and habitat characteristics predict litter-mixing effects on decomposition[J].Ecology, 2011, 92:160-169.
[48]LOSOS J B.Phylogenetic niche conservatism, phylogenetic signal and the relationship between phylogenetic relatedness and ecological similarity among species[J].Ecology Letters, 2008, 11:995-1003.
[49]DIAZ S, LAVOREL S, DE BELLO F, et al.Incorporating plant functional diversity effects in ecosystem service assessments[J].Proceedings of the National Academy of Sciences of the United States of America, 2007, 104:20684-20689.
[50]AUGUSTO L, DE SCHRIJVER A, VESTERDAL L, et al.Influences of evergreen gymnosperm and deciduous angiosperm tree species on the functioning of temperate and boreal forests[J].Biological Reviews, 2015, 90:444-466.
[51]JOLY F-X, MILCU A, SCHERER-LORENZEN M, et al.Tree species diversity affects decomposition through modified microenvironmental conditions across European forests[J].New Phytologist, 2017, 214:1281-1293.
[52]MAKKONEN M, BERG M P, VAN LOGTESTIJN R S P, et al.Do physical plant litter traits explain non-additivity in litter mixtures? A test of the improved microenvironmental conditions theory[J].Oikos, 2013, 122:987-997.
[53]SCHERER-LORENZEN M, LUIS BONILLA J, POTVIN C.Tree species richness affects litter production and decomposition rates in a tropical biodiversity experiment[J].Oikos, 2007, 116:2108-2124.
[54]覃宇.马尾松与阔叶树种混合凋落叶中难降解物质的分解特征[D].雅安:四川农业大学, 2019.
[55]H TTENSCHWILER S, TIUNOV A V, SCHEU S.Biodiversity and Litter Decomposition in Terrestrial Ecosystems[J].Annual Review of Ecology Evolution & Systematics, 2005, 36(1):191-218.
[56]CORTEZ J, GARNIER E, PREZ-HARGUINDEGUY N, et al.Plant traits, litter quality and decomposition in a Mediterranean old-field succession[J].Plant & Soil, 2007, 296(1-2):19-34.
[57]SCHIMEL J P, H TTENSCHWILER S.Nitrogen transfer between decomposing leaves of different N status[J].Soil Biology & Biochemistry, 2007, 39(7):1428-1436.
[58]BONANOMI G, INCERTI G, ANTIGNANI V, et al.Decomposition and nutrient dynamics in mixed litter of Mediterranean species[J].Plant & Soil, 2010, 331(s1-2):481-496.
[59]MAKKONEN M, BERG M P, HANDA T I,et al.Highly consistent effects of plant litter identity and functional traits on decomposition across a latitudinal gradient[J].Ecology Letters, 2012, 15(9):1033-1041.
[60]LIN H, ZHANG N L, MA K P.What drives leaf litter decomposition and the decomposer community in subtropical forests-The richness of the above-ground tree community or that of the leaf litter?[J].Soil Biology and Biochemistry, 2021, 160:108314.
[61]GILLESPIE L M, H TTENSCHWILER S, SHIHAN A, et al.Tree species mixing affects soil microbial functioning indirectly via root and litter traits and soil parameters in European forests[J].Functional Ecology, 2021,35:2190-2204.
[62]LECERF A, KOMINOSKIJ.Functional leaf traits and biodiversity effects on litter decomposition in a stream: Comment[J].Ecology, 2010, 91(6):1867-1871.
[63]HANDA I T, AERTS R, BERENDSE F, et al.Consequences of biodiversity loss for litter decomposition across biomes[J].Nature, 2014, 509(7499):218-221.
[64]GROSSMAN J J, JEANNINE CAVENDER-BARES, HOBBIE S E.Functional diversity of leaf litter mixtures slows decomposition of labile but not recalcitrant carbon over two years[J].Ecological Monographs, 2020,90(3):e01407.
(責任编辑:柯文辉)
猜你喜欢
现代园艺(2022年7期)2022-11-19 10:18:42
林业科技(2022年5期)2022-10-08 12:00:16
四川劳动保障(2021年3期)2021-06-09 07:08:56
西藏农业科技(2019年3期)2019-11-04 00:35:08
水土保持通报(2018年6期)2018-02-13 07:44:52
现代园艺(2017年13期)2018-01-19 02:28:05
广东农业科学(2017年5期)2017-08-29 10:37:19
西南农业学报(2016年5期)2016-05-17 05:42:36
中国农业文摘-农业工程(2016年5期)2016-04-12 05:38:09
华南农业大学学报(2015年5期)2015-12-04 03:04:38
