
中国谷子种业创新现状与未来展望
2022-04-21 08:28贾冠清刁现民
中国农业科学 2022年4期
贾冠清,刁现民
中国谷子种业创新现状与未来展望
贾冠清,刁现民
中国农业科学院作物科学研究所,北京 100081
种业是农业发展的“芯片”,原始创新是农业可持续发展的根本动力。2021年中央一号文件明确提出推进中国农作物种业快速发展的要求,对中国农作物种业原始创新研究提出了明确期望。谷子是中国起源的传统粮饲兼用作物及特色杂粮作物,生产及消费规模均位居世界首位。作为粟类作物,谷子在中国农业生产及农耕文明的起始与发展过程中发挥了重要的推动和支撑作用,已有研究证实谷子在中国拥有悠久的栽培历史,并且形成了分布广泛且多样性丰富的各类种质资源。近年来,谷子在杂交品种选育及杂种优势利用、抗除草剂品种和适于机械化栽培品种推广、基因组学及功能基因研究等领域取得进展,在原始创新的推动下初步形成了以杂交品种和抗除草剂品种为经营主体的谷子种业体系,推动了谷子种业从无到有的突破。中国在谷子优异种质鉴定与创制方法研究、谷子高效育种技术途径研发、谷子关键性状的协调表达与调控规律解析、谷子良种繁育过程基本生物学属性研究以及谷子新品种真实性鉴定方法探索等方面原始创新的进步为谷子种业发展提供了支撑,形成了一套初具规模的种业原始创新技术。目前,谷子种业持续良性发展仍然面临包括优异突破性种质匮乏、育种技术水平相对滞后、品质与产量协调性不够、良种扩繁标准缺乏以及种业市场监管手段有待加强等诸多挑战。为了推进谷子种业持续快速、高效、深入的发展,未来中国谷子种业原始创新研究的主要方向包括:基于表型组学与基因组修饰技术、单倍体育种与全基因组选择技术和关键性状优异单倍型鉴定与整合技术的谷子规模化高效育种技术体系构建;基于种子发育生理调控、分子指纹及杂种优势高效利用技术的谷子种子高效生产储藏与监管技术体系构建;以及基于谷子种业产学研推一体化设计与高效整合的人才培养与创新体系构建等方面。
谷子;种业;原始创新;现状;发展趋势
中国是农作物生产大国和农业用种大国,种业已经发展成为中国基础性、战略性核心产业,是保障国家粮食安全,推动农业稳定可持续发展的根本保证。国务院于2012年发布了《全国现代农作物种业发展规划(2012—2020年)(国办发[2012]59号)》,对中国农作物种业的发展目标进行了规划。2021年中央一号文件再次强调了中国种业发展的紧迫性和必要性,提出了“打赢种业翻身仗”的目标要求,对中国种业实现跨越发展提出了时间表和路线图。谷子又称为粟,是中国传统的粮饲兼用作物,具有抗旱耐瘠薄、营养丰富、保健功能突出等特点,是民众膳食结构改善和种植业结构调整的主体作物[1]。谷子起源于中国黄河流域,在中国的栽培历史可以追溯到距今11 500年前[2],种质资源保有量大且多态性丰富,是中华农耕文明发展的重要载体之一[3]。随着国家“乡村振兴”战略的持续深入实施,谷子作为中小作物的代表,在满足人民膳食健康需求以及推动种业科技进步方面发挥着越来越重要的作用。近年来,中国谷子遗传改良、种子繁育及品种推广的发展逐渐进入了快车道,为深入推动谷子产业和种业高效可持续发展,本文从原始创新角度论述了中国谷子种业发展中面临的问题以及未来的发展方向,为政府决策以及基础科研工作提供参考。
1 中国谷子种业发展历史与现状
1.1 谷子的产业地位
谷子曾经是中国的主要口粮作物,在20世纪50—60年代,谷子种植面积约1 000万hm2,仅次于水稻和小麦,是中国的三大主栽作物之一。随着人民生活水平的提高,以及对肉蛋奶等畜牧产品需求的增加,畜牧业的迅速发展导致玉米等高生物量作物种植面积逐年扩大,同时,由于中国机械化生产条件的不断进步,劳动力成本的逐渐升高以及种植业结构调整政策的实施,谷子的种植面积从20世纪70年代开始表现出减少的趋势[4]。据国家谷子高粱产业技术体系统计,2020年中国谷子种植面积约150万hm2,总产量约1 000万t,是重要的杂粮作物,生产及消费规模均位居世界首位。除了用作口粮外,谷子在酿造业、食品加工业中也发挥了重要作用,并且谷子在中国牧区也被广泛用作牧草,是维持国民经济发展不可或缺的经济作物。在国际贸易方面,谷子原粮主要出口至日本、韩国、德国、荷兰和印度尼西亚等国家,是中国的优势出口作物[5]。
1.2 谷子种业的发展过程
中国谷子品种选育历史悠久,历史上曾经形成了“四大贡米”等优质名牌品种,包括山西沁县的“沁州黄”、河北蔚县的“桃花米”、山东济宁的“金乡小米”以及山东章丘的“龙山小米”,但没有形成规模化的育种体系[6];谷子近代育种工作可以追溯至20世纪30—40年代,新中国成立前,延安时期的光华农场最早开展了新品种的选育研究。金陵大学、燕京作物改良试验场和华北农业科学研究所等单位通过系统育种先后育成并推广了燕京811等谷子品种[7];新中国成立后的50—70年代,谷子生产主要采用常规品种,“粮种不分”,生产依赖于农民自留的农家品种,种子生产资料依靠农业合作社/生产大队自繁、自选、自留、自用,辅之以调剂的方针。在计划经济体制下,谷子没有发展种业,生产用种主要依靠自留及国家调配;改革开放初期的80—90年代,尽管谷子育种有了长足进步,以豫谷1号[8]、昭谷1号[9]和晋谷21[10]为代表的一系列品种培育成功,产量水平得到显著提高,但这些品种均为常规品种,虽有一定的种子经营,但自留种很普遍,谷子种业发展处于萌芽阶段,没有形成规模性的种业。从21世纪初开始,随着抗拿捕净(烯禾啶)除草剂谷子新品种的培育成功,以抗除草剂谷子品种为核心的谷子种业开始形成[11],同时谷子杂种优势在生产上得以成功应用[12-13],基于杂交种种子生产的种子企业也随之形成并发展壮大。2000年《种子法》颁布实施之后,谷子种业发展开始进入商业化时期,民营企业逐步增多成为市场主体,并逐渐向育、繁、推、销一体化转变。其中,河北巡天种业新技术有限责任公司迅速发展,2018年净利润达到了7 300万元,成为中国谷子种业的龙头企业;河北东昌种业有限公司则在抗除草剂谷子新品种经营上创立了品牌,也发展成为谷子种子经营的龙头企业;内蒙古蒙龙种业科技有限公司、内蒙古禾为贵种业有限公司、内蒙古利禾农业科技发展有限公司等一批以谷子种子生产和经营为主体的企业在市场中确立了地位,谷子种业体系初步形成,为谷子种业的高效商业化发展奠定了基础。
目前,谷子种业发展的核心依然为新品种的选育。截至2021年8月底,中国完成登记谷子新品种497个(图1):其中,内蒙古自治区163个,河北省159个,山西省64个,吉林省31个,辽宁省28个,河南省16个,山东省14个,甘肃省7个,北京市5个,黑龙江省4个,陕西省4个,贵州省1个,新疆维吾尔自治区1个。主产省区中只有宁夏尚未有品种登记。从登记单位看,企业登记238个,占47.9%,科研单位登记259个,占52.1%;从品种类型看,497个登记品种中:常规品种448个,占90.1%,杂交品种49个,占9.9%,饲草专用品种1个,粮饲兼用品种1个;只适合春播的336个,占67.6%,春夏播皆宜的139个,占27.9%,仅适合夏播的22个,占4.5%;抗除草剂品种179个,占37.4%,其中常规品种130个,占常规品种登记总数的29.1%,49个杂交种均为抗除草剂品种。谷子种业发展迎来了蓬勃发展期[14]。
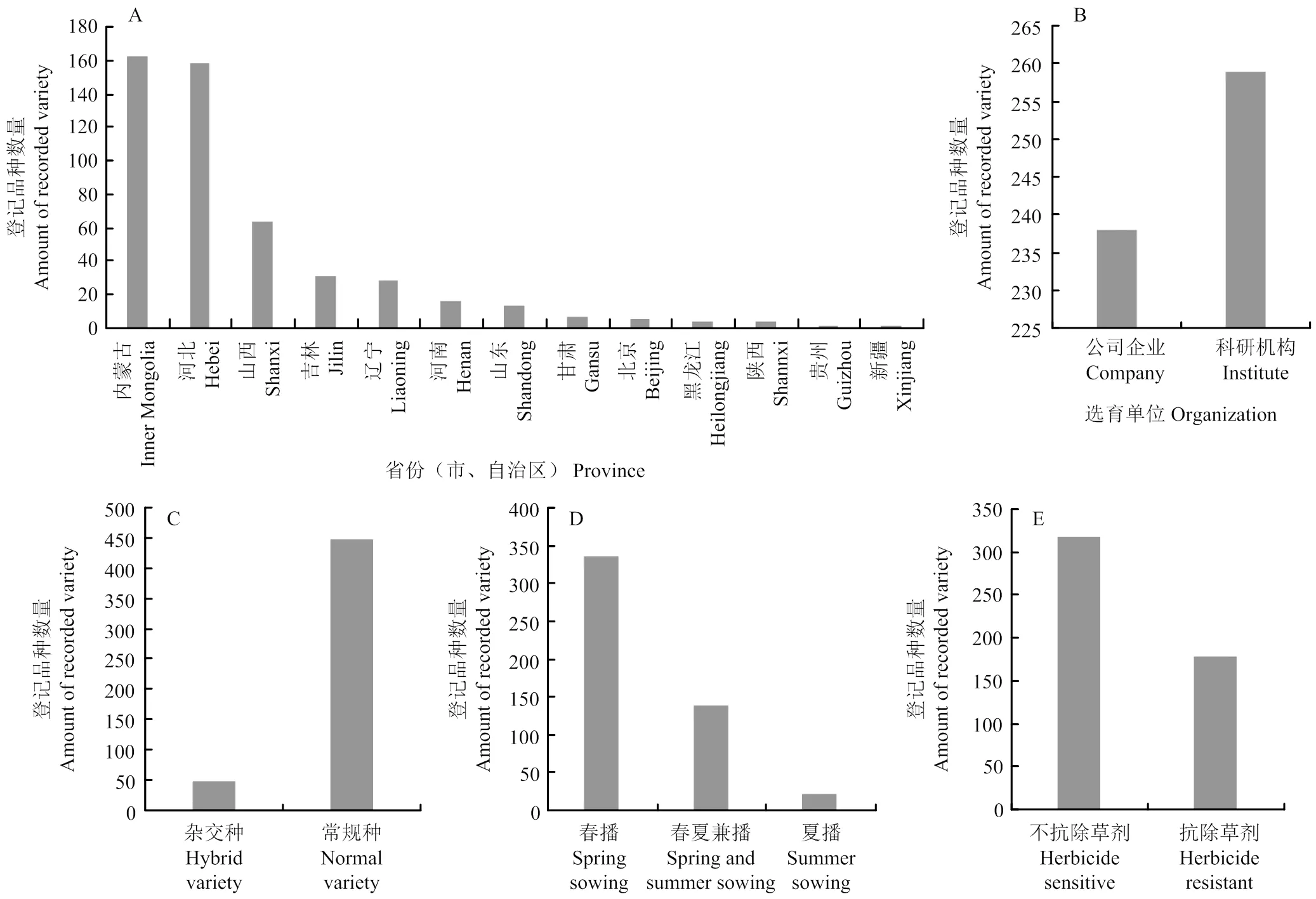
A:各省登记品种分布情况;B:登记品种主体分布;C:登记品种类型组成;D:登记品种适种区域分布;E:登记品种抗除草剂特性
1.3 谷子种业原始创新主要历史突破
商业化品种和种业技术的原始创新是种业发展的基础,随着中国谷子种业持续深入发展,种业对原始创新的形式及内容提出了更高的要求。截至目前,中国保存有各类谷子资源接近3万份,保存量位居世界第一,主要为中国境内收集的地方品种。高度雄性不育系“蒜系28”的育成,正式开启了谷子杂种优势的育种利用[15],通过远缘杂交技术创制的抗除草剂“拿捕净”、“氟乐灵”、“阿特拉津”和“咪唑乙烟酸”等新种质彻底改变了谷子产业化生产模式,并推进了杂交种商业化应用[11]。国外引进新种质“六十日”在夏谷品种选育中作为骨干亲本,在抗倒伏方面发挥了重要作用[16];矮秆骨干亲本“矮88”的创制和育种利用,在全国多个生态区实现株高的显著降低,衍生品种已达100多个,基本解决了谷子倒伏和适应机械化作业的问题;优质品种晋谷21的培育和应用,为小米产业化发展奠定了基础,累计推广种植面积已经超过70万hm2。在不同历史时期,中国还先后选育出了昭谷1号、豫谷1号、冀谷19[17]、张杂谷5号[18]、豫谷18[19]、中谷2等代表性品种,据国家谷子高粱产业技术体系统计,谷子育种进步使夏谷区平均产量由原来的4 500 kg·hm-2提升到7 500 kg·hm-2,为谷子种业持续发展奠定了基础。
1.4 近年来谷子种业原始创新取得的成绩
谷子种业原始创新主要集中于新品种选育环节。2008年国家谷子现代农业产业技术体系启动,通过持续开展原始创新攻关,近十年来,谷子育成品种和原始创新取得了显著进步:在品种株高改良方面,平均株高降低了31 cm,使得机械化管理和收割成为可能;通过远缘杂交将野生种青狗尾草的天然抗除草剂基因转育到谷子中,使谷子由不抗除草剂的作物转变为抗除草剂的作物,通过实现轻简化栽培解决了数千年来谷子栽培人工间苗、人工除草等效率低下问题,生产方式发生革命性改变,轻简栽培技术使得单户综合生产能力提高20倍以上。在解决这些核心卡脖子技术的基础上,育成了覆盖华北、西北、东北三大谷子主产区抗除草剂、矮秆、宜机收的冀谷39[20]、冀谷42、豫谷35[21]、张杂谷13[22]、中杂谷5等系列轻简栽培新品种;在品质改良方面,近十年来,育成了品质与传统主栽优质品种黄金苗相当,并且抗除草剂、产量和抗性优于黄金苗的金苗K1[23]、冀谷45、中谷9等品种,基本打破了农家品种在优质米开发中占主导的局面,为优质小米的产业化开发提供了品种保证;育成适合食品加工新品种冀谷42,其籽粒亚油酸与油酸比值达到3.7,比普通品种降低了32.7%,延长了货架期。多元化谷子新品种选育有效支撑了谷子种业的深入发展。
在种业基础研究方面,率先在国际上完成了谷子基因组的测序[24],构建了国际上第一个谷子基因组单倍型图谱和关联分析群体,发掘出一批谷子重要性状相关的基因位点[25],率先构建了高效的遗传转化体系[26],转化效率是国外同类研究机构的3倍以上,构建了核心种质和突变体库,形成了较为完善的谷子功能基因发掘和基因编辑的育种平台,发起并组织了首届国际谷子遗传学大会,提出并推动谷子成为功能基因研究的模式作物[27],这些研究工作使谷子这一传统作物焕发出新的青春,使中国的谷子遗传育种研究保持国际领先地位。
2 谷子种业原始创新技术研发进展及需求
2.1 优异种质鉴定与创制
据国家谷子高粱产业技术体系统计,中国目前保存谷子种质资源28 915份,占世界谷子资源总量的70%。“十三五”以来入库谷子资源1 733份,引进国外谷子资源46份,狗尾草资源77份,搜集整理国内谷子资源244份,提供育种单位利用4 106份。近五年鉴定出谷子抗除草剂、抗病、抗旱、抗倒、矮秆材料2 236份,创制出谷子抗烟嘧磺隆材料4份,抗咪唑乙烟酸材料2份,创制出5份小米商品性显著提高的夏谷育种材料,2份蒸煮时间显著缩短(减少30%左右)且米色鲜黄的春谷育种材料,有效支撑了国内谷子新品种的选育工作。然而,目前鉴定和创制优异种质材料的技术仍然局限于常规的田间种植和远缘杂交与回交转育,尚未规模开展理化诱变及优异种质定向鉴选研究。远缘杂交创新种质规模较小,目前仅局限于除草剂抗性材料的创制[5],尚未成功获得抗病虫及抗旱新种质。此外,商品性及蒸煮品质优异材料鉴定规模较小,无法满足种业发展对优质品种的需求。在技术层面,张婷等[28]发现小米商品性和蒸饭品质极显著正相关,但是和煮饭品质不相关,并且认为通过L*值、a*值、b*值对米色进行选择是一种直观、便捷地判定小米商品性的方法。目前,谷子优异种质鉴定与创制主要受到表型组技术发展的制约,在规模和效率上均需要提高,此外由于已知功能基因与调控元件解析的数量过少,基因编辑技术平台不够完善,目前尚没有实现规模化应用,导致优异种质创制的效率偏低。
2.2 谷子新品种高效选育
谷子的遗传改良技术经历了由农家种选择单株(代表性品种:晋谷1号、花脸1号等)到杂交后代系谱选择(代表性品种:新农冬2号、豫谷1号等),由利用自然变异到人工创造变异(组织培养:代表品种冀张谷6号(又称矮88);诱变育种:代表品种张农10号),由常规自交品种选育到杂交种组合选育(代表性品种:张杂谷5号、龙杂谷1号等),由一年一季选择到一年两季南北穿梭育种等过程,虽然育种速度有所提升,但是新品种的原始创新性提升缓慢,主要表现在:1)真正在产量、品质、抗逆、株型等方面有突破性的品种较少,登记品种间趋同性明显,多数新品种实际为优良骨干品种的派生品种;2)快速繁殖与加代技术缺乏,获得稳定的品系需要3—5年的时间,常规育种效率提升缓慢;3)单倍体育种技术缺乏,无法快速获得双单倍体育种亲本,制约了杂交种组配与鉴定选择的效率;4)基于分子标记的辅助选择技术体系缺乏,无法在早期开展定向遗传背景/遗传前景选择,连锁累赘剔除效率低且育种成功率不高;5)基因组选择及预测技术缺乏,无法进行有效的表型预测,品种选育过程依赖经验和运气,效率提升缓慢;6)基于转基因的育种技术缺乏,无法快速开展抗病虫、抗除草剂等育种工作,导致种业市场化提升效率缓慢。
常规杂交育种技术仍然是谷子主流遗传改良技术,随着种业的持续深入发展,实现品质与产量的协同表达对于促进谷子种业的可持续发展至关重要。关于品质性状与产量性状间的协调关系,李荫梅等[29]研究认为,谷子蛋白质含量与籽粒产量呈负相关。刘三才等[30]研究发现小米硒含量与蛋白质含量正相关,并且红粒谷子硒含量明显高于黄、青、褐等粒色的品种。邵丽华等[31]研究发现小米颜色差异显著影响叶酸含量,从高到低依次为:褐色、绿色、黄色、鲜黄、浅黄和白色米粒品种。刘思辰等[32]发现谷子蛋白质含量与茎长、码数存在显著正相关,与穗粗、码粒数、单穗重、穗粒重和千粒重存在负相关;同时发现淀粉含量与千粒重正相关,与码数和蛋白质含量存在负相关;此外还发现淀粉直/支比与穗粗、千粒重正相关,与码数负相关。然而,由于谷子品质调控关键遗传因子尚不清楚,产量相关性状的遗传解析刚刚起步(表1),目前还无法形成对关键农艺性状协调表达规律的整体认识,现有结果还无法直接应用于新品种的选育过程中,分子育种知识原始创新的不足限制了谷子新品种选育技术的进步以及育种效率的提高。
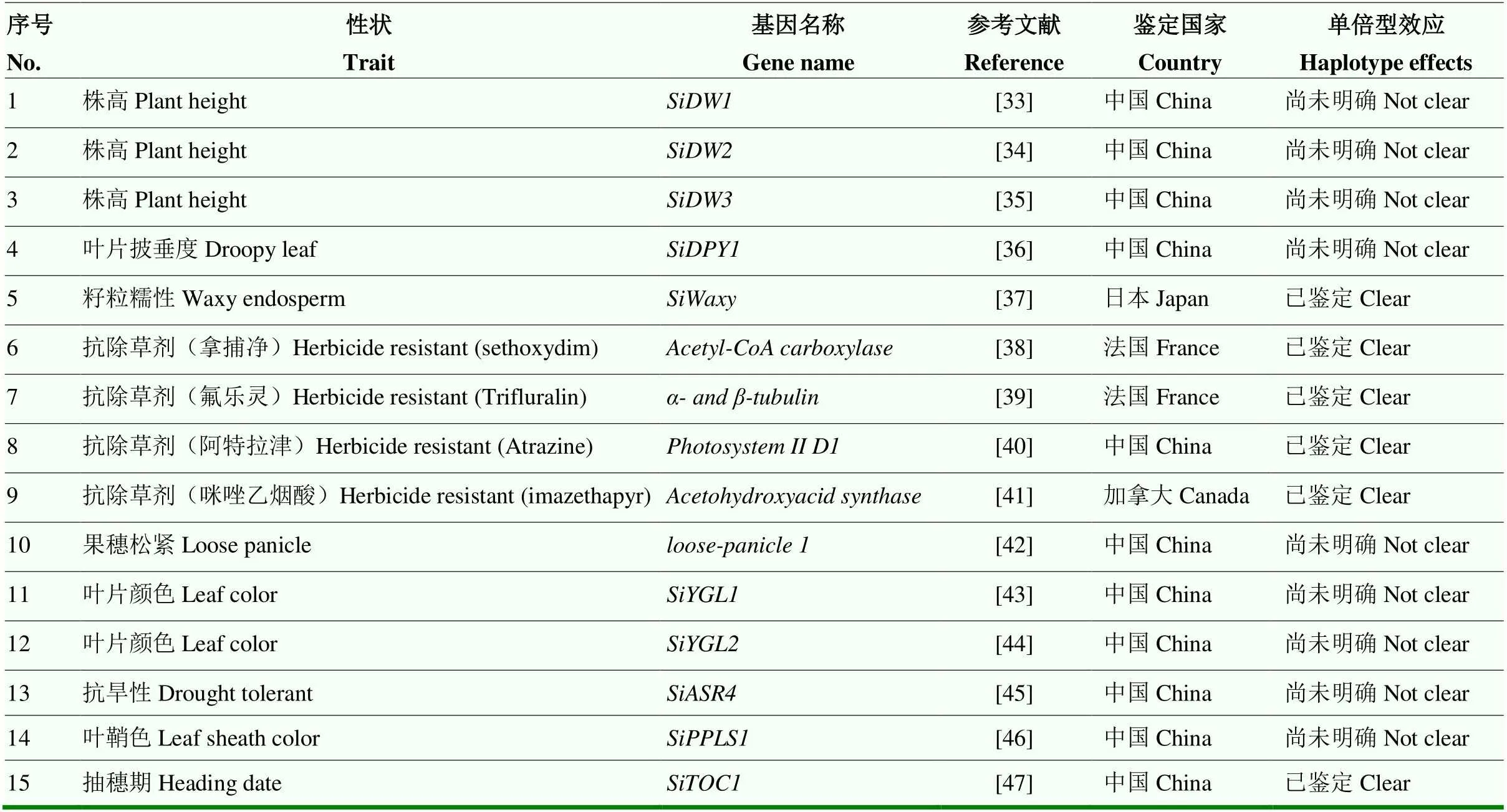
表1 已知谷子关键农艺性状调控基因
2.3 良种繁育及质量控制
谷子良种繁育包括了常规自交品种以及杂交种,涉及种子高效生产、种子快速安全脱水、种子加工和质量控制、种子贮藏和运输分发等过程。智慧等[48]建立了通过加速老化鉴定谷子种子活力的方法,研究发现采用41℃处理48 h,可以有效检测种子活力的差异,并且发现加速老化后的种子发芽率与田间出苗率正相关,可以用来预测田间出苗率。近年来,谷子种子生产和质量控制相关研究几乎处于停滞状态,随着谷子精量播种技术的普及,对谷子种子质量提出了更高的要求。提高良种繁育技术水平,从原始创新的角度,目前亟需开展谷子天然异交率形成与控制的生物学研究,以及相应的物理化学调控技术研发,此外种子成熟过程中活性、水分的变化规律及其遗传、物理调控机制都需要进行解析,在此基础上,形成谷子良种繁育系列技术标准和方法,提高良种繁育的效率和市场竞争力,推动谷子种业的深入发展。
2.4 良种推广与市场监测
品种真实性检测是保证种业市场规范稳定、加快良种推广的主要技术手段。传统的DUS(Distinctness特异性,Uniformity一致性,Stability稳定性)测试鉴定周期长、受环境影响大,从原始创新角度,目前亟需建立基于分子标记手段的快速谷子品种真实性鉴定技术作为补充。Jia等[4]采用SSR分子标记构建了新中国建国以来培育的348个主要谷子品种的分子指纹数据库,研究发现春谷和夏谷类型的品种在中国种业市场中的比例经历了由春谷为主到目前以夏谷为主的转变,并且针对这一变化,整理出了可区分不同谷子品种的SSR分子标记,为建立谷子品种真实性分子指纹技术奠定了基础。然而,随着中国种业市场的规范化深入发展,不仅需要建立谷子登记品种的分子指纹数据库,同时还需要建立针对新选育品种创新性的指纹鉴定标准,在育种过程中避免品种趋同,并在种子营销过程中避免套牌现象的发生,真正做到维护种业市场健康有序发展。
3 谷子种业原始创新问题分析
3.1 优异突破性种质较匮乏,骨干亲本储备不够
从高效、集约、宜机收和商业化的种业发展趋势来看,谷子新品种在现有基础上仍然需要着力解决突破性种质的创制与鉴定问题:
(1)耐密植种质缺乏:通过密植实现增产增效是未来种业的发展方向,也是禾谷类作物近代育种的成功经验。耐密植种质需要同时兼顾株型合理及抗逆性突出的特点,实现谷子直立紧凑株型与抗病虫、耐庇荫、水肥高效的结合。直立紧凑株型通过改良叶片形态可以改善田间密植条件下的通风透光性,提高光合效能及减少病虫害,抗逆性的提高可实现密植条件下单株产量水平的稳定,从而达到密植增产的效果[49]。目前,谷子种质资源中虽然存在一定数量的株型紧凑、叶片直立的材料,但往往穗部较小且结实性差,对产量影响很大。由于缺乏相应的突破性种质,谷子新品种尚未实现直立紧凑株型的改良,可用于密植生产的新品种极为缺乏。
(2)矮秆抗早衰种质单一,现有矮源遗传基础不明确:抗倒伏是谷子田间生产需要着力解决的问题,矮化育种是解决抗倒性的有效手段。目前谷子矮秆育种主要依靠矮88、吨谷等材料。利用矮88及其衍生系为亲本,已经培育出100余个新品种,但多数为中秆品种。已有的矮88降秆基因的遗传解析表明,其株高受第1和第9染色体上的3个主效降秆基因和多个微效降秆基因所控制[50],并且不同于水稻绿色革命的和小麦绿色革命的,具体机制尚不明确,同时,尽管发掘出了骨干亲本矮88的众多穗部性状QTL信息[51],深入的基因解析尚没有开展。其他在育种中利用的矮源包括吨谷、金谷2、矮宁黄等矮秆材料遗传基础均缺乏研究,更多的谷子矮秆种质普遍存在不同程度的早衰现象,很难在育种中利用。
(3)籽粒快速脱水种质缺乏:轻简栽培在谷子生产中越来越重要,田间收获后的籽粒晾晒问题制约了谷子机械化收获及仓储的效率。谷子收获后传统晾晒过程耗时、费力,无法实现收储连续作业,并且谷子不及时脱水对小米品质影响很大,已经不再适合产业化组织条件下作物机械化生产的需要[52]。需要培育成熟后期快速脱水的谷子新品种,可以在田间达到收获和储运的籽粒安全含水量,并且对小米品质影响较小。目前,针对适合收储连续作业的快脱水谷子新种质创新处于起步阶段,由于谷子育种长期以绿秆成熟为目标,对快速脱水的育种材料缺乏认识,需要开展从头创制,解决谷子“直收粒”面临的配套品种匮乏的问题。
(4)广适性种质依然缺乏:谷子是典型的光温敏感作物,区域适应性窄[53],限制了种业的发展规模。提高谷子品种的广适应性,扩大优良品种的适种区域,是做大做强谷子种业的必然要求。目前谷子生产及育种中广适性品种匮乏,无法支撑种业的深入发展和竞争力的提升。创制广适性新种质是解决种业发展规模问题的有效途径。选育的豫谷18、中谷2、冀谷168等品种表现出了广适性特征,可以实现跨生态区种植,但是均为夏谷类型,遗传背景较为单一,需要继续拓宽广适性谷子品种类型,推进谷子种业快速高效发展。
3.2 育种技术水平相对滞后,无法支撑种业深入发展
自20世纪80年代以来,中国谷子的诱变育种、杂种优势利用先后取得了突破性进展,选育出了张农10号、晋谷21等代表性品种,之后组织培养、远缘杂交、分子标记辅助育种等也取得了较好进展,使中国谷子育种形成了杂交系选育种为主,其他育种手段为辅的多元技术育种格局。目前,谷子新品种选育仍然以杂交后代系选为主,产业化生产中的新品种绝大部分为自交品种,杂交种品种所占比例较低(图1)。从育种技术的整体应用角度而言,谷子分子育种技术手段尚未得以应用,分子标记选择及聚合育种的技术体系刚刚起步,转基因及基因编辑技术尚在建立完善阶段,单倍体育种技术虽然初步构建了诱导系,但距离实际应用还有距离[54],导致中国谷子育种技术发展相对滞后,定向遗传改良技术储备及应用严重不足,制约了谷子种业发展效率和水平的进一步提高。
3.3 品质与产量协调性不够,品种市场竞争力有待提高
品质和产量的协调提升问题仍然制约了谷子种业的深入发展,感官加工品质和蒸煮食味品质的定向改良尚未实质性开展,导致大部分谷子新品种的市场竞争力偏弱,米质的提高已经成为了种业规模化发展的首要限制因素。由于目前开展米质定向改良的技术手段尚不完备,导致相关新品种选育进展缓慢,无法满足种业快速发展的需求。
3.4 谷子杂种优势利用技术单一,杂交种纯度和制种效率亟待改进
谷子杂交种虽然已经实现了产业化应用,但目前杂交种制种利用的是高度雄性不育两系法[11],质核互作三系法和光温敏感两系法在谷子中尚未成功应用。高不育两系法最大的问题是杂交种纯度低,必须依靠恢复系的抗除草剂基因进行种子处理或苗期处理剔除假杂种;同时杂交种制种效率低,影响了种业发展。此外,由于谷子杂交制种依赖高度雄性不育基因,不育系本身通过自交进行保持,通过恢复系与不育系杂交进行制种。为了平衡不育系繁种效率及杂交种的制种需求,商业化的杂交种真杂种率普遍较低,商业化种子的价值受到一定程度影响。同时由于不育系遗传背景有限,且缺少配合力等遗传分析,尚未建立杂种优势群,依据经验的优势杂交组合组配效率较低,制约了杂交种选育的效率和水平。
3.5 谷子良种扩繁标准缺乏,种子质量缺乏保证
目前,谷子种子市场的国家标准主要参照2008年修订的农作物种子质量标准(标准号:GB4404.1- 2008),该标准为多年前制定,已经不适合目前的谷子生产。谷子种业目前处于无国家标准、无行业标准的状态运行。部分种子公司通过制定自己的标准来满足市场监管,极易造成谷子种子市场的混乱。由于谷子杂交种为高不育两系法杂交种,其种子实际为真杂交种种子和不育系自交种子的混合体,制定统一的标准加强监管极为必要。
谷子的高质量种子生产技术以及良种繁育技术体系不够完善,主要体现在高活力种子生产技术、种子快速安全脱水及质量控制技术缺乏,尚未形成常规品种及杂交种制种行业标准与技术规范。由于谷子种子质量尚未形成规范高效的统一标准,在一定程度上制约了谷子种业市场的深入健康发展,也为种子质量的深入监管增加了难度。
3.6 品种趋同及侵权问题较严重,种业市场监管不够
谷子育种材料、优异种质及新品种知识产权的保护力度不够,尚未建立起市场认可的多元化的品种鉴定试验及DUS测试体系,品种登记后的评价机制缺乏,没有形成谷子DNA指纹及分子身份证标准,直接导致种业市场监管难度大,品种趋同现象及套牌侵权问题严重,极大影响了育种者的积极性和种子企业的利益,严重影响了谷子种业的健康发展。
4 谷子种业原始创新的发展方向
中国谷子种业深入发展高度依赖原始创新,新品种选育效率的提升以及制种与鉴定水平的提升可以有效推动谷子种业的快速发展(图2),未来种业原始创新的发展方向主要包括如下几个方面:
4.1 谷子新品种规模化高效选育与评价鉴选技术
作物表型的高通量精准鉴定与基因组的修饰技术代表了未来农作物遗传改良的方向。美国国家科学基金会于2016年将表型组学列为6大科研前沿之一[55]。作物表型一般指能够反映细胞、组织、器官、植株和群体结构及功能特征的物理、生理和生化性状,体现了作物基因图谱的时序三维表达及其地域分布特征和代际演进规律。作物表型组学研究代表了作物科学发展的前沿方向,对开发作物种质资源、培育突破性作物品种保障国家粮食安全具有重要意义[56]。
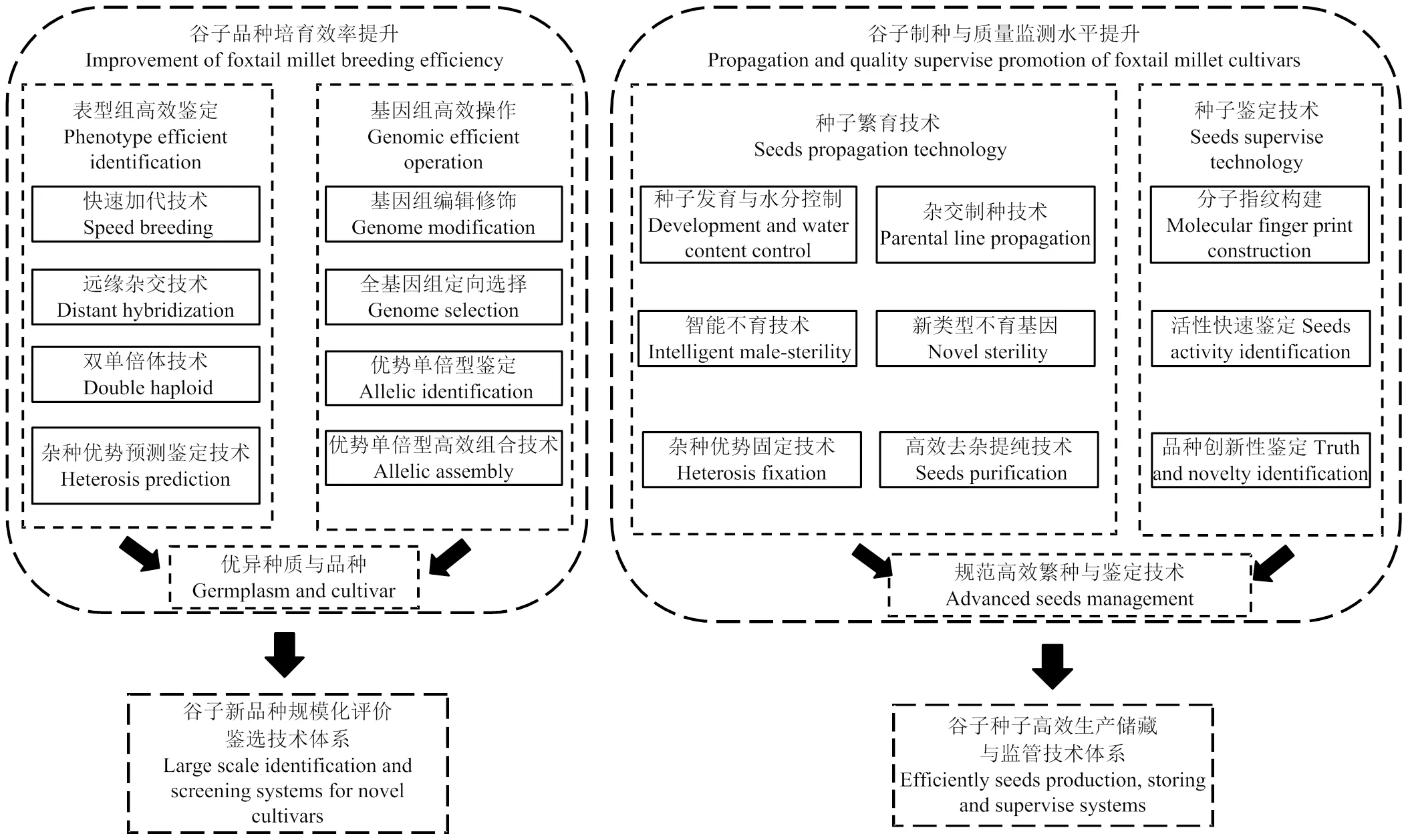
图2 中国谷子种业原始创新发展技术路线
基因组编辑技术近年来发展迅速,Gao[57]认为基因组编辑技术通过实现基因组定点区段变异、基因组定点单碱基替换、基因组定点插入、基因组定点删除以及基因组定点重组,目前已经形成了加速作物遗传改良的8个主要技术途径:1)通过直接定点编辑功能基因(或同源基因)以及引入DNA双链断裂实现功能性改变和定点重组;2)采用多靶点方式实现多倍体同源等位基因编辑或多基因家族编辑;3)编辑数量性状位点获得性状梯度变异新材料;4)实现野生植物的从头驯化,创制新的作物类型;5)通过多基因编辑实现作物杂种优势的固定,改变杂交种的制种方式;6)通过构建先导RNA文库对目标基因进行饱和突变,结合高通量筛选策略实现基因快速进化;7)通过将特定序列元件定点整合至作物基因组中,采用合成生物学策略加速作物驯化;8)通过原位编辑作物微生物菌群实现作物生长力及抗病性改良。谷子的高效遗传转化技术已经突破[26],可以预见,未来基因组编辑技术结合高通量表型组学技术、快速加代育种技术以及全基因组选择技术,一定可以成为加速谷子新品种改良的有效技术手段,推动种业的快速发展。
单倍体育种技术是近年来逐渐发展成熟的快速育种技术,通过采用花粉培养、诱导孤雌生殖及孤雄生殖等方式,经过染色体加倍快速得到纯合稳定的二倍性育种材料,大大缩短育种周期。已有研究表明,单倍体诱导基因在禾谷类作物间具有高度保守性[58],而且在谷子中敲除类似基因同样可以实现单倍体诱导[54],证实了在谷子中开展单倍体育种的可行性,相信随着单倍体诱导效率以及加倍效率的提高,可以大幅度提高谷子育种效率。
全基因组选择技术是后基因组时代迅速发展起来的通过基因型开展表型预测的选择技术[59],通过基因芯片等技术开展待筛选材料的基因型鉴定,可以对目标性状表现进行早期预测并开展材料筛选,通过大幅度减少田间种植鉴定规模,提高育种选择效率。将全基因组选择技术与单倍体育种技术相结合,可以构建出工程化育种体系,在快速高通量获得纯合育种材料的同时,实现早期性状预测,为杂交种选育效率提升提供技术支撑。
优异等位基因的整合与选择技术是育种工作的核心。张林林等[47]在谷子中鉴定到抽穗期基因,发现该基因存在优势单倍型,并且早熟单倍型品种与晚熟单倍型品种在不同纬度条件下平均早抽穗10 d,可以在育种工作中进行选择应用。这一研究结果表明谷子关键农艺性状不仅存在优势单倍型,而且在更多关键功能基因中开展优异单倍型的鉴定研究,可以在育种应用的同时提高品种选育效率。Wei等[60]在水稻中实现了数量性状核酸位点数据库的构建,并系统解析了不同单倍型的表型效应,在分子预测的基础上,实现了水稻目标性状的快速定向改良。这些研究结果表明,深入开展关键功能基因的单倍型效应解析以及整合技术研究,可以在剔除连锁累赘的基础上,有效提升作物品种改良的效率和水平,实现真正意义上的分子选择育种,加快现有育种进程。
规模化多点联合鉴定技术体系对新品种综合鉴评至关重要[61]。截至2016年,中国通过国家和省级审定和鉴定的谷子品种累计达到870多个,为了深入推动谷子新品种选育进程,从2001年起,全国农业技术推广服务中心组织了国家谷子联合区域试验,该试验科学稳定地执行了15年,到2016年,随着新种子法的实施停止执行[62]。从谷子种业发展的长远需求来看,中国仍然需要建立谷子新品种多生态区联合鉴定体系,促进品种适应性评价。通过创新谷子新品种多生态区、多试验点联合鉴定组织模式,实现谷子种业可持续深入发展。
4.2 谷子种子高效繁育与质量监管技术
良种扩繁与机械化收储技术的优化主要依赖于种子发育及其水分生理调控规律研究。Jia等[4]通过分析中国谷子主要育成品种的基因组特征,发现谷子的天然异交率为3%—5%,Shi等[63]研究了谷子花粉飘散规律及混杂风险,发现在20 m距离仍然可以检测到0.002%的混杂。Yano等[64]对小米黄色素积累与收获时期间的关系进行研究,发现谷子抽穗后积温达到850℃时为最佳收获期,此时黄色素含量最高,商品性最好,过度成熟反而会导致小米黄色素含量的下降。截至目前,关于种子活力与收获期的关系,种子水分积累规律与影响因素等都缺少研究,杂交种制种关键环节的基本生物学问题尚未解析清楚,严重制约了良种繁育的效率和种子质量的提升,对种业发展的影响极大。今后围绕良种扩繁和机械化收储需求开展种子发育及其遗传生理调控研究,可以有效解决这一问题。
为了规范种业市场,避免品种雷同和套牌经营,最为有效快捷的途径就是建立品种的分子指纹数据库,并形成相应的国家标准,在品种登记、品种权申请等环节进行监测,对市场流通销售的种子进行检查,杜绝侵权套牌现象的发生。目前,中国农业科学院作物科学研究所于2016年获得发明专利“一组区分谷子品种的SSR标记及其应用”(专利号:ZL 2015 1 0028660.5),已经具备了构建谷子分子指纹数据库的基础,相信不久的将来可以形成一套完整规范的谷子品种分子指纹鉴定技术,服务于谷子种业发展。
分子指纹除了可以辅助判定品种雷同性,还可以用于开展杂种优势群的划分以及杂交组合优势的判定,提高谷子杂交种选育及优势组合选配的效率和水平。目前,基于分子指纹技术划分谷子杂种优势群的研究相对较少,还无法指导杂交种的快速选育,今后应加强相关研究,为提高谷子杂种优势利用水平和种业发展质量提供数据基础。
4.3 谷子种业产学研推体系一体化设计与高效整合
谷子是中国传统特色杂粮作物,在满足民众由“吃得饱”逐渐向“吃得好”膳食需求转变的过程中发挥着不可替代的重要作用。商品化谷子生产对新品种的特征特性提出了新的要求,商品感官与食味营养品质是决定未来谷子种业市场发展的关键,抗除草剂品种及杂交种的推广应用是未来谷子种业发展的主要方向。未来应当优化顶层设计,依托涉农高校和科研院所,构建覆盖遗传育种、种子科学、分子生物学、大数据与自动化技术及市场营销管理等交叉学科的种业科学专业,积极开展专业人才培养及核心技术研发,同时依托种业企业科技力量,建设公益性种业科技与企业种业科技优势互补、相互配合的种业科技研发力量,在育繁推销一体化技术链条各个环节培养适应新时代要求的种业科技人才,积极开展谷子种业核心原创技术的研发,形成具有自主知识产权的谷子新品种规模化评价鉴选技术体系和谷子种子高效生产储藏与监管技术体系,突破卡脖子技术瓶颈,实现谷子种业健康高效可持续发展。
已有研究表明,谷子种业依托原始创新实现了种业发展“0到1”的突破,可以预见,不久的将来,通过继续深入开展原始创新研究,在关键技术环节实现更多的技术突破,必将能够推动谷子种业继续做大做强,实现产业水平和能力的快速提升,为保证中国人民的膳食健康做出积极的贡献。
[1] 刁现民. 禾谷类杂粮作物耐逆和栽培技术研究新进展. 中国农业科学, 2019, 52(22): 3943-3949.
DIAO X M. Progresses in stress tolerance and field cultivation studies of orphan cereals in China. Scientia Agricultura Sinica, 2019, 52(22): 3943-3949. (in Chinese)
[2] YANG X Y, WAN Z W, PERRY L, LU H Y, WANG Q, ZHAO C H, LI J, XIE F, YU J C, CUI T X, WANG T, LI M Q, GE Q S. Early millet use in northern China. Proceedings of the National Academy of Sciences of the United States of America, 2012, 109(10): 3726-3730.
[3] 贾冠清, 刁现民. 谷子((L.) P. Beauv.)作为功能基因组研究模式植物的发展现状及趋势. 生命科学, 2017, 29(3): 292-301.
JIA G Q, DIAO X M. Current status and perspectives of researches on foxtail millet ((L.) P. Beauv.): A potential model of plant functional genomics studies. Chinese Bulletin of Life Sciences, 2017, 29(3): 292-301. (in Chinese)
[4] JIA G Q, LIU X T, SCHNABLE J C, NIU Z G, WANG C F, LI Y H, WANG S J, WANG S Y, LIU J R, GUO E H, ZHI H, DIAO X M. Microsatellite variations of elite Setaria varieties released during last six decades in China. PLoS One, 2015, 10(5): e0125688.
[5] 李顺国, 刘裴, 刘猛, 程汝宏, 夏恩君, 刁现民. 中国谷子产业和种业发展现状与未来展望. 中国农业科学, 2021, 54(3): 459-470.
LI S G, LIU F, LIU M, CHENG R H, XIA E J, DIAO X M. Current status and future prospective of foxtail millet production and seed industry in China. Scientia Agricultura Sinica, 2021, 54(3): 459-470. (in Chinese)
[6] 程汝宏. 产业化生产背景下的谷子育种目标. 河北农业科学, 2010, 14(11): 92-95.
CHENG R H. Breeding objectives of foxtail millet under the background of industrial production. Journal of Hebei Agricultural Sciences, 2010, 14(11): 92-95. (in Chinese)
[7] 国家谷子糜子产业技术体系. 中国现代农业产业可持续发展战略研究-谷子糜子分册. 北京: 中国农业出版社, 2018.
China Agriculture Research System of Foxtail millet and Common millet. Strategic research of sustainable development of Chinese agriculture-Foxtail millet and Common millet. Beijing: China Agricultural Press, 2018. (in Chinese)
[8] 胡咸廷. 豫谷1号. 新农业, 1985(5): 25.
HU X T. Yugu 1. Xin Nongye, 1985(5): 25. (in Chinese)
[9] 蒋士鸿. 昭谷1号. 农业科技通讯, 1981(2): 8.
JIANG S H. Zhaogu 1. Bulletin of Agricultural Science and Technology, 1981(2): 8. (in Chinese)
[10] 史更生, 史关燕, 史宏, 陈瑛. 晋谷21号优良品质的分析与研究. 杂粮作物, 2001, 21(4): 18-19.
SHI G S, SHI G Y, SHI H, CHEN Y. High quality analysis of millet variety ‘Jingu 21’. Rain Fed Crops, 2001, 21(4): 18-19. (in Chinese)
[11] 王天宇, 杜瑞恒, 陈洪斌, 亨利达门西, 阿兰弗郎夷. 应用抗除草剂基因型谷子实行两系法杂种优势利用的新途径. 中国农业科学, 1996, 29(4): 96.
WANG T Y, DU R H, CHEN H B, DARMENCY H, FLEURY A. A new way of using herbicide resistant gene on hybrid utilization in foxtail millet. Scientia Agricultura Sinica, 1996, 29(4): 96. (in Chinese)
[12] 赵治海, 崔文生, 杜贵, 杨少青. 谷子光(温)敏不育系821选育及其不育性与光、温关系的研究. 中国农业科学, 1996, 29(5): 23-31.
ZHAO Z H, CUI W S, DU G, YANG S Q. The selection of millet photo (thermo) sensitive sterile line 821 and a study on the relation of sterility to illumination and temperature. Scientia Agricultura Sinica, 1996, 29(5): 23-31. (in Chinese)
[13] 王玉文, 王随保, 李会霞, 王高鸿, 田岗, 史琴香. 谷子光敏雄性不育系选育及应用研究. 中国农业科学, 2003, 36(6): 714-717.
WANG Y W, WANG S B, LI H X, WANG G H, TIAN G, SHI Q X. Studies on breeding of photoperiod-sensitive male-sterile line in millet and its application. Scientia Agricultura Sinica, 2003, 36(6): 714-717. (in Chinese)
[14] 李荣德, 程汝宏, 陈应志, 孙海艳, 史梦雅. 谷子品种登记实施进展与建议. 种子, 2019, 38(8): 150-153, 158.
LI R D, CHENG R H, CHEN Y Z, SUN H Y, SHI M Y. Progress and suggestions on the implementation of millet variety registration. Seed, 2019, 38(8): 150-153, 158. (in Chinese)
[15] 崔文生, 马洪锡, 张德勇. 谷子雄性不育系“蒜系28”的选育与利用. 中国农业科学, 1979, 12(1): 43-46.
CUI W S, MA H X, ZHANG D Y. The selection and utilization of “suan his 28”-a male sterility strain of millet. Scientia Agricultura Sinica, 1979, 12(1): 43-46. (in Chinese)
[16] 陈家驹, 王雅儒, 王升文, 吴舒致, 王尧琴. 抗倒伏谷子种质“六十日”及其衍生系统的应用. 中国种业, 1984, 3: 30-32+35.
CHEN J J, WANG Y R, WANG S W, WU S Z, WANG Y Q. Lodging resistant foxtail millet germplasm “liu shi ri” and its system application. China Seed Industry, 1984, 3: 30-32+35. (in Chinese)
[17] 程汝宏, 刘正理, 师志刚, 夏学岩, 杨万桥. 水分高效利用型谷子新品种“冀谷19”选育研究. 河北农业科学, 2006, 10(1): 80-81.
CHENG R H, LIU Z L, SHI Z G, XIA X Y, YANG W Q. Breeding of foxtail millet cultivar Jigu19. Journal of Hebei Agricultural Sciences, 2006, 10(1): 80-81. (in Chinese)
[18] 乔春花, 杨立军, 李登来. 优质高产杂交谷子新品种“张杂谷5号”. 现代农村科技, 2009(22): 17.
QIAO C H, YANG L J, LI D L. High quality and grain yield hybrid foxtail millet variety ‘Zhangzagu 5’. Xiandai nongcun keji, 2009(22): 17. (in Chinese)
[19] 闫宏山, 刘金荣, 王素英, 路志国, 刘海平, 蒋自可, 宋中强, 王淑君. 谷子新品种豫谷18的选育. 作物杂志, 2012, 3: 147-148.
YAN H S, LIU J R, WANG S Y, LU Z G, LIU H P, JIANG Z K, SONG Z Q, WANG S J. Breeding of foxtail millet variety Yugu18. Crops, 2012, 3: 147-148. (in Chinese)
[20] 河北省农林科学院谷子研究所. 优质抗除草剂谷子新品种-冀谷39. 现代农村科技, 2018(12).
INSTITUTE OF MILLET CROPS, HEBEI ACADEMY OF AGRICULTURE AND FORESTRY SCIENCES. High quality and herbicide resistant foxtail millet variety Jigu39. Xiandai nongcun keji, 2018(12). (in Chinese)
[21] 闫宏山, 宋慧, 张杨, 邢璐, 解慧芳, 魏萌涵, 付楠, 刘金荣. 抗拿捕净除草剂谷子新品种豫谷35的选育. 中国种业, 2019, 2: 80-81.
YAN H S, SONG H, ZHANG Y, XING L, XIE H F, WEI M H, FU N, LIU J R. Breeding of sethoxydim-resistant foxtail millet variety ‘Yugu35’. China Seed Industry, 2019, 2: 80-81. (in Chinese)
[22] 宋国亮, 赵治海, 王晓明, 王峰, 范光宇, 冯小磊, 赵芳, 史高雷, 张晓磊. 优质杂交谷子新品种张杂谷13号的选育及栽培技术. 现代农村科技, 2019, 5: 18.
SONG G L, ZHAO Z H, WANG X M, WANG F, FAN G Y, FENG X L, ZHAO F, SHI G L, ZHANG X L. Breeding and cultivating of high quality foxtail millet hybrid variety ‘Zhangzagu13’. Xiandai nongcun keji, 2019, 5: 18. (in Chinese)
[23] 王显瑞, 柴晓娇, 白晓雪, 沈轶男, 付颖, 刘艳春, 刘丹竹, 张姼, 李书田. 优质抗旱抗除草剂谷子新品种“金苗K1”的选育及高产栽培技术. 耕作与栽培, 2021, 41(2): 63-64.
WANG X R, CHAI X J, BAI X X, SHEN Y N, FU Y, LIU Y C, LIU D Z, ZHANG S, LI S T. Breeding of high quality and drought resistance and herbicide-resistant foxtail millet new variety ‘Jinmiao K1’ and key points of high-yield cultivation techniques. Tillage and Cultivation, 2021, 41(2): 63-64. (in Chinese)
[24] ZHANG G, LIU X, QUAN Z, CHENG S, XU X, PAN S, XIE M, ZENG P, YUE Z, WANG W, TAO Y, BIAN C, HAN C, XIA Q, PENG X, CAO R, YANG X, ZHAN D, HU J, ZHANG Y, LI H, LI H, LI N, WANG J, WANG C, WANG R, GUO T, CAI Y, LIU C, XIANG H, SHI Q, HUANG P, CHEN Q, LI Y, WANG J, ZHAO Z, WANG J. Genome sequence of foxtail millet () provides insights into grass evolution and biofuel potential. Nature Biotechnology, 2012, 30: 549-554.
[25] JIA G, HUANG X, ZHI H, ZHAO Y, ZHAO Q, LI W, CHAI Y, YANG L, LIU K, LU H, ZHU C, LU Y, ZHOU C, FAN D, WENG Q, GUO Y, HUANG T, ZHANG L, LU T, FENG Q, HAO F, LIU H, LU P, ZHANG N, LI Y, GUO E, WANG S, WANG S, LIU J, ZHANG W, CHEN G, ZHANG B, LI W, WANG Y, LI H, ZHAO B, LI J, DIAO X, HAN B. A haplotype map of genomic variations and genome-wide association studies of agronomic traits in foxtail millet (). Nature Genetics, 2013, 45(8): 957-961.
[26] YANG Z, ZHANG H, LI X, SHEN H, GAO J, HOU S, ZHANG B, MAYES S, BENNETT M, MA J, WU C, SUI Y, HAN Y, WANG X. A mini foxtail millet with an-like life cycle as a C4 model system. Nature Plants, 2020, 6: 1167-1178.
[27] Diao X, Schnable J, Bennetzen J, LI J. Initiation ofas a model plant. Frontiers of Agricultural Science and Engineering, 2014, 1(1): 16-20.
[28] 张婷, 王根平, 罗焱杰, 李琳, 高翔, 程汝宏, 师志刚, 董立, 张喜瑞, 杨伟红, 许立闪. 色差分析在优质小米选育中的应用. 中国农业科学, 2021, 54(5): 901-908.
ZHANG T, WANG G P, LUO Y J, LI L, GAO X, CHENG R H, SHI Z G, DONG L, ZHANG X R, YANG W H, XU L S. Color difference analysis in the application of high quality foxtail millet breeding. Scientia Agricultura Sinica, 2021, 54(5): 901-908. (in Chinese)
[29] 李荫梅, 凌莉. 夏谷品质与主要农艺性状的相关性分析. 华北农学报, 1989(S1): 44-50.
LI Y M, LING L. Analysis of correlation between quality character and other main agronomic characters of summer-sown foxtail millet. Acta Agriculturae Boreali-Sinica, 1989(S1): 44-50. (in Chinese)
[30] 刘三才, 朱志华, 李为喜, 刘方, 李燕, 黄蓉. 谷子品种资源微量元素硒和蛋白质含量的测定与评价. 中国农业科学, 2009, 42(11): 3812-3818.
LIU S C, ZHU Z H, LI W X, LIU F, LI Y, HUANG R. Evaluation of selenium and protein content of foxtail millet landraces originated from different ecological regions of China. Scientia Agricultura Sinica, 2009, 42(11): 3812-3818. (in Chinese)
[31] 邵丽华, 王莉, 白文文, 刘雅娟. 山西谷子资源叶酸含量分析及评价. 中国农业科学, 2014, 47(7): 1265-1272.
SHAO L H, WANG L, BAI W W, LIU Y J. Evaluation and analysis of folic acid content in millet from different ecological regions in Shanxi province. Scientia Agricultura Sinica, 2014, 47(7): 1265-1272. (in Chinese)
[32] 刘思辰, 曹晓宁, 温琪汾, 王海岗, 田翔, 王君杰, 陈凌, 秦慧彬, 王纶, 乔治军. 山西谷子地方品种农艺性状和品质性状的综合评价. 中国农业科学, 2020, 53(11): 2137-2148.
LIU S C, CAO X N, WEN Q F, WANG H G, TIAN X, WANG J J, CHEN L, QIN H B, WANG L, QIAO Z J. Comprehensive evaluation of agronomic traits and quality traits of foxtail millet landrace in Shanxi. Scientia Agricultura Sinica, 2020, 53(11): 2137-2148. (in Chinese)
[33] ZHAO M C, ZHI H, ZHANG X, JIA G Q, DIAO X M. Retrotransposon-mediated DELLA transcriptional reprograming underlies semi-dominant dwarfism in foxtail millet. The Crop Journal, 2019, 7(4): 458-468.
[34] XUE C X, ZHI H, FANG X J, LIU X T, TANG S, CHAI Y, ZHAO B H, JIA G Q, DIAO X M. Characterization and fine mapping of SiDWARF2 (D2) in foxtail millet. Crop Science, 2016, 56: 95-103.
[35] FAN X K, TANG S, ZHI H, HE M M, MA W S, JIA Y C, ZHAO B H, JIA G Q, DIAO X M. Identification and fine mapping of SiDWARF3 (D3), a pleiotropic locus controlling environment independent dwarfism in foxtail millet. Crop Science, 2017, 57: 2431-2442.
[36] ZHAO M C, TANG S, ZHANG H S, HE M M, LIU J H, ZHI H, SUI Y, LIU X T, JIA G Q, ZHAO Z Y, YAN J J, ZHANG B C, ZHOU Y H, CHU J F, WANG X C, ZHAO B H, TANG W Q, LI J Y, WU C Y, LIU X G, DIAO X M. DROOPY LEAF1 controls leaf architecture by orchestrating early brassinosteroid signaling. Proceedings of the National Academy of Sciences of the United States of America, 2020, 117(35): 21766-21774.
[37] NAKAYAMA H, AFZAL M, OKUNO K. Intraspecific differentiation and geographical distribution of Wx alleles for low amylose content in endosperm of foxtail millet,(L.) P. Beauv. Euphytica, 1998, 102: 289-293.
[38] DéLYE C, WANG T, DARMENCY H. An isoleucine-leucine substitution in chloroplastic acetyl-CoA carboxylase from green foxtail (L. Beauv.) is responsible for resistance to the cyclohexanedione herbicide sethoxydim. Planta, 2002, 214: 421-427.
[39] DéLYE C, MENCHARI Y, MICHEL S, DARMENCY H. Molecular bases for sensitivity to tubulin-binding herbicides in green foxtail. Plant Physiology, 2004, 136: 3920-3932.
[40] JIA X, YUAN J, SHI Y, SONG Y, WANG G, WANG T, LI Y. A Ser-Gly substitution in plastid-encoded photosystem II D1 protein is responsible for atrazine resistance in foxtail millet (). Plant Growth Regulation, 2007, 52: 81-89.
[41] LAPLANTE J, RAJCAN L, TARDIF F J. Multiple allelic forms of acetohydroxyacid synthase are responsible for herbicide resistance in. Theoretical and Applied Genetics, 2009, 119: 577-585.
[42] XIANG J S, TANG S, ZHI H, JIA G Q, WANG H J, DIAO X M. Loose Panicle1 encoding a novel WRKY transcription factor, regulates panicle development, stem elongation, and seed size in foxtail millet [(L.) P. Beauv.]. PLoS One, 12(6): e0178730.
[43] LI W, TANG S, ZHANG S, SHAN J G, TANG C J, CHEN Q N, JIA G Q, HAN Y H, ZHI H, DIAO X M. Gene mapping and functional analysis of the novel leaf color gene SiYGL1 in foxtail millet [(L.) P. Beauv]. Physiologia Plantarum, 2016, 157: 24-37.
[44] ZHANG S, ZHI H, LI W, SHAN J, TANG C, JIA G, TANG S, DIAO X. SiYGL2 is involved in the regulation of leaf senescence and photosystem II efficiency in(L.) P. Beauv. Frontiers in Plant Science, 2018, 9: 1308.
[45] LI J, DONG Y, LI C, PAN Y, YU J. SiASR4, the target gene of SiARDP from, improves abiotic stress adaption in plants. Frontiers in plant science, 2017, 7: 2053.
[46] BAI H, SONG Z, ZHANG Y, LI Z, WANG Y, LIU X, MA J, QUAN J, WU X, LIU M, ZHOU J, DONG Z, LI D. The bHLH transcription factor PPLS1 regulates the color of pulvinus and leaf sheath in foxtail millet (). Theoretical and Applied Genetics, 2020, 133: 1911-1926.
[47] 张林林, 智慧, 汤沙, 张仁梁, 张伟, 贾冠清, 刁现民. 谷子抽穗时间基因的表达与单倍型变异分析. 中国农业科学, 2021, 54(11): 2273-2286.
ZHANG L L, ZHI H, TANG S, ZHANG R L, ZHANG W, JIA G Q, DIAO X M. Characterization of transcriptional and haptotypic variations of
[48] 智慧, 陈洪斌, 凌莉, SUSANA GOGGI, YUH-YUAN SHY. 加速老化法测定谷子种子活力的研究. 中国农业科学, 1999, 32(3): 66-71.
ZHI H, CHEN H B, LING L, GOGGI S, SHY Y. Studies on accelerated aging seed vigor test for foxtail millet. Scientia Agricultura Sinica, 1999, 32(3): 66-71. (in Chinese)
[49] 程式华. 中国超级稻育种技术创新与应用. 中国农业科学, 2016, 49(2): 205-206.
CHENG S H. Breeding technique innovation and application China’s super rice. Scientia Agricultura Sinica, 2016, 49(2): 205-206. (in Chinese)
[50] HE Q, ZHI H, TANG S, XING L, WANG S, WANG H, ZHANG A, LI Y, GAO M, ZHANG H, CHEN G, DAI S, LI J, YANG J, LIU H, ZHANG W, JIA Y, LI S, LIU J, QIAO Z, GUO E, JIA G, LIU J, DIAO X. QTL mapping for foxtail millet plant height in multi-environment using an ultra-high density bin map. Theoretical and Applied Genetics, 2021, 134: 557-572.
[51] ZHI H, HE Q, TANG S, YANG J, ZHANG W, LIU H, JIA Y, JIA G, ZHANG A, LI Y, GUO E, GAO M, LI S, LI J, QIN N, ZHU C, MA C, ZHANG H, CHEN G, ZHANG W, WANG H, QIAO Z, LI S, CHENG R, LU X, WANG S, LIU J, LIU J, DIAO X. Genetic control and phenotypic characterization of panicle architecture and grain yield-related traits in foxtail millet (). Theoretical and Applied Genetics, 2021, 134: 3023-3036.
[52] 柴宗文, 王克如, 郭银巧, 谢瑞芝, 李璐璐, 明博, 侯鹏, 刘朝巍, 初振东, 张万旭, 张国强, 刘广周, 李少昆. 玉米机械粒收质量现状及其与含水率的关系. 中国农业科学, 2017, 50(11): 2036-2043.
CHAI Z W, WANG K R, GUO Y Q, XIE R Z, LI L L, MING B, HOU P, LIU C W, CHU Z D, ZHANG W X, ZHANG G Q, LIU G Z, LI S K. Current status of maize mechanical grain harvesting and its relationship with grain moisture content. Scientia Agricultura Sinica, 2017, 50(11): 2036-2043. (in Chinese)
[53] 王殿瀛, 陈玉香. 谷子品种南北异地种植主要性状变化及其应用. 中国农业科学, 1980, 13(4): 33-38.
WANG D Y, CHEN Y X. The main characters variation of the millet varieties grown both in north and south areas and its use in practice. Scientia Agricultura Sinica, 1980, 13(4): 33-38. (in Chinese)
[54] Cheng Z, Sun Y, Yang S, Zhi H, Yin T, Ma X, Zhang H, Diao X, Guo Y, Li X, Wu C, Sui Y. Establishing in planta haploid inducer line by edited SiMTL in foxtail millet (). Plant Biotechnology Journal, 2021, 19(6): 1089-1091.
[55] Córdova F. NSF director unveils big ideas. Science, 2016, 352(6287): 755-756.
[56] YANG W, FENG H, ZHANG X, ZHANG J, DOONAN J, BATCHELOR W, XIONG L, YAN J. Crop phenomics and high- throughput phenotyping: past decades, current challenges, and future perspectives. Molecular Plant, 2020, 13: 187-214.
[57] GAO C. Genome engineering for crop improvement and future agriculture. Cell, 2021, 184: 1621-1635.
[58] ZHU H, LI C, Gao C. Applications of CRISPR-Cas in agriculture and plant biotechnology. Nature Reviews Molecular Cell Biology, 2020, 21: 661-677.
[59] DESTA Z A, ORTIZ R. Genomic selection: genome-wide prediction in plant improvement. Trends in Plant Science, 2014, 19: 592-601.
[60] WEI X, QIU J, YONG K, FAN J, ZHANG Q, HUA H, LIU J, WANG Q, OLSEN K M, HAN B, HUANG X. A quantitative genomics map of rice provides genetics insights and guides breeding. Nature Genetics, 2021, 53: 243-253.
[61] 盖钧益, 刘康, 赵晋铭. 中国作物种业科学技术发展的评述. 中国农业科学, 2015, 48(17): 3303-3315.
GAI J Y, LIU K, ZHAO J M. A review on advances in science and technology in Chinese seed industry. Scientia Agricultura Sinica, 2015, 48(17): 3303-3315. (in Chinese)
[62] 刁现民, 程汝宏. 十五年区试数据分析展示谷子糜子育种现状. 中国农业科学, 2017, 50(23): 4469-4474.
DIAO X M, CHENG R H. Current breeding situation of foxtail millet and common millet in China as revealed by exploitation of 15 years regional adaptation test data. Scientia Agricultura Sinica, 2017, 50(23): 4469-4474. (in Chinese)
[63] SHI Y, WANG T, LI Y, DARMENCY H. Impact of transgene inheritance on the mitigation of gene flow between crops and their wild relatives: the example of foxtail millet. Genetics, 2008, 180: 969-975.
[64] YANO A, TAKAKUSAGI M, QIKAWA K, NAKAJO S, SUGAWARA T. Xanthophyll levels in foxtail millet grains according to variety and harvesting time. Plant Production Science, 2017, 20(1): 136-143.
Current status and perspectives of innovation studies related to foxtail millet seed industry in China
JIA GuanQing, DIAO XianMin
Institute of Crop Sciences, Chinese Academy of Agricultural Sciences, Beijing 100081
Seed industry was the ‘chip’ of agricultural development, and original innovation have played essential roles in maintaining stable development of modern agriculture. The No. 1 central document of China released in 2021 has put forward requirements of original innovation researches essential for supporting further developments of Chinese crop seed industry. Foxtail millet is a traditional crop species cultivated for both forage and grain food consumption in China, and to date, foxtail millet was still widely planted as minor cereals in China with the largest scale of field production and commercial consumption across the globe. Foxtail millet was originated from China and cultivated for thousands of years to ensure development of Chinese agricultural culture and field crop production. Original innovation in foxtail millet has promoted initial development of foxtail millet seed industry based on operation of herbicide-resistant varieties in recent decades, including breakthroughs in areas of heterosis utilization, herbicide resistant breeding, dwarfing variety creation and genomics study of this important crop species. Achievements of fundamental researches including germplasm characterization, development of breeding tools, coordination and regulation of vital traits, seeds propagation and truth identification of commercial varieties have provided more opportunities for further development of the seed industry of foxtail millet. However, challenges of seed industry development still exist in China, including deficiency of excellent germplasms, backward breeding approaches, inharmonious of yield and quality characters, unclear criterion of seed propagation and market supervision problems. Future direction of original innovation studies related to foxtail millet seed industry were as follows: 1) large scale breeding systems constructed from utilizations of crop phenomics and genomic modification technologies, double haploid breeding and genome selection tools, identification and pyramiding of superior haplotypes; 2) seeds production, store and quality supervision systems constructed from techniques of development and water content of commercial seeds, and establishment of molecular fingerprints and efficient utilization of heterosis in foxtail millet; 3) Construction of innovation system through integrating education, research and promotion sectors of seed industry and stimulate personnel training in China.
foxtail millet; seed industry; original innovation; development status; future perspectives

10.3864/j.issn.0578-1752.2022.04.003
2021-09-06;
2021-11-29
国家自然科学基金(31871630)、财政部和农业农村部:国家现代农业产业技术体系(CARS-06-14.5-A04)、中国农业科学院创新工程
贾冠清,E-mail:jiaguanqing@caas.cn。通信作者刁现民,E-mail:diaoxianmin@caas.cn
(责任编辑 李莉)
猜你喜欢
今日农业(2022年4期)2022-11-16
农业知识(2022年9期)2022-10-13
农业知识(2022年9期)2022-10-13
中国农业科技导报(2022年7期)2022-09-27
今日农业(2022年14期)2022-09-15
上海农业科技(2022年3期)2022-06-11
今日农业(2022年2期)2022-06-01
当代陕西(2021年21期)2022-01-19
延河(2018年6期)2018-06-11
农业工程技术·温室园艺(2009年9期)2009-05-17
