
龙竹丛状采伐恢复后林分小气候特征分析*
2022-04-20 04:51陶航涂丹丹刘蔚漪辉朝茂王友
西部林业科学 2022年2期
陶航,涂丹丹,刘蔚漪,辉朝茂,王友
(1.西南林业大学,竹藤科学研究院,丛生竹工程技术研究中心,云南 昆明 650224;2.云南经济管理学院,云南 安宁 650106)
竹资源是森林资源的重要组成部分,我国拥有大量的竹资源。由于地形地貌限制,劳动力成本逐渐上升,机械自动化程度低,竹资源利用程度低、种类少等因素造成了大量竹资源未得到有效采伐利用[1]。竹林是异龄林,传统采伐方式以择伐为主,采伐原则为采伐老龄竹、衰老竹,留存幼龄竹、壮龄竹,在林地里始终保持留存有各个龄级的林木[2],传统择伐方式目前已经无法满足竹产业的发展。针对毛竹(Phyllostachysedulis)一类的散生竹提出了一种全新采伐方式——带状采伐,即按照一定宽度对毛竹林实施条带式皆伐[3];针对小型丛生硬头黄竹(Bambusarigida)也有带状皆伐对其生长及其产量的影响的研究[4]。但对于大型材用丛生竹传统择伐作业困难、采伐成本高等问题,需探索出适用于大型丛生竹培育、能改善传统采伐方式带来的弊端并适用于机械化采伐的龙竹(Dendrocalamusgiganteus)丛状采伐技术模式。由于丛状采伐的采伐强度和采伐面积较大,可能会对龙竹林内小气候造成一些影响,从而影响到新竹的生长,因此有必要对不同采伐方式下的林间小气候因子进行研究。
森林小气候是森林植被影响下形成的特殊小气候,是森林中水、气、热等各种气象要素综合作用的结果[5]。森林小气候对林下植物的生长和发育、有机质分解、土壤温度、土壤酶活性等生态过程都有影响[6]。小气候的改变会引起植物物种组成、盖度、生物量、生理生态因子等发生相应的变化,森林生态系统能发挥其生态系统屏障功能,对植物的生长发育、气候调节和改善环境均有重大意义[7]。现国内外对抚育采伐对森林小气候的影响和改变传统竹林经营模式以达到丰产的研究较多,但这些研究大都是针对传统林分[8-11]以及散生竹竹林开展的[12-14],而关于龙竹这类的大型丛生竹的抚育改造以及小气候的动态变化对新竹生长影响的研究尚处于空白阶段。
龙竹是禾本科(Poaceae)牡竹属(DendrocalamusNees)的一种大型材用丛生竹,在中国云南东南至西南部各地均有分布,台湾也有栽培,国外在亚洲热带和亚热带各国家都有栽培[15],可作为建筑和篾用竹材。本研究通过对不同采伐强度的龙竹林内主要气象因子、新竹生长对不同丛状采伐强度小气候响应的分析和研究,探讨不同强度丛状采伐对龙竹林内小气候的动态变化进而筛选出影响新竹生长的主要因子,以了解龙竹林经营过程中生态环境气候因子与生产之间的关系,为建立适用于机械化作业的丛生竹林的培育技术模式提供理论依据和科学指导。
1 材料与方法
1.1 试验地概况
试验地位于云南省临沧市沧源县勐角乡(99°13′23″~99°15′24″E、23°12′01″~23°10′18″N,海拔1 474~1 850 m),土壤为红壤,土层厚度80 cm以上,肥力中上,适宜龙竹生长。区域内气候温和,雨量充沛,日照充足,冬无严寒,夏无酷暑。年极端最低气温-4.3 ℃,最高气温为33.7 ℃,年平均气温17.4 ℃,年平均日照为1 876.7 h。年均降水量1 763.5 mm,全年无霜期长达317 d。
1.2 试验设置与数据收集
于2018年冬季建立了龙竹机械化采伐培育试验示范林66 666.67 m2,试验设置4种采伐强度处理,分别为:A 常规择伐、B 1/2丛伐、C 1/3丛伐,D全丛采伐(小区之间间隔30 m),按生长健壮的高效培育林分结构算,平均666.67 m2约18丛。为保证试验数据具有统计学意义,选择大约1 000丛龙竹来做试验,设置4个小区,A、B、C每个试验小区的边长大约为180 m×20 m,各有9个标准样地(20 m×20 m),D试验小区的边长大约为360 m×20 m,有18个标准样地(20 m×20 m)。2019年3、6、9、12月中旬,监测样地内光照强度(Lx)、风速(km/h)、空气湿度(%)、空气温度(℃)、50 cm土层土壤温度(℃)等小气候因子的变化,设置每1 h读存数据一次。
竹林调查 在采伐后第一年内(6—10月)对发笋时间、发笋数量(株/丛)等进行跟踪调查,待笋期结束后以每丛为单位调查新竹数量(株/丛)、胸径(cm)等。
新竹生物量测定 将新竹地上部分伐倒,测量并记录新竹胸径(cm)、株高(m)。将竹秆、竹枝、竹叶分离并称量其鲜重。把竹秆进行分段,每段1 m,将每段新竹秆破开于中部取样0.6~0.9 kg。竹枝、竹叶分离前将竹株上、中、下3个部位取样并混合,取样质量分别控制在0.4~0.5 kg、0.3~0.4 kg。将样品带回实验室置于105 ℃的烘箱中烘至恒量,计算得到含水率(%)。再通过将各器官鲜重含水率换算成干重,得到竹秆、竹枝、竹叶生物量(t/hm2),把各器官生物量相加得到单株生物量(t/hm2)。
光照因子测量仪器为TES-1332A光照强度计(TES,Taiwan),土壤温度因子测量仪器为WatchDog纽扣式温度,记录仪风速、空气湿度、空气温度因子测量仪器为Testo-610温湿度仪(Testo AG,Germany)。
1.3 数据处理及分析
将太阳光照强度、空气温度、土壤温度、空气湿度、风速等小气候因子进行步进逐步回归分析,逐步输入自变量,剔除未进入模型的变量,以进行关键生态因子的筛选。
利用方差分析探讨采伐方式对龙竹林内光照强度、土壤温度的影响。
各小气候因子进行相关性分析,衡量各小气候因子与新竹生长之间的关系。
采用主成分分析进行降维,得到综合指标。将不同的样地小气候数据通过主成分分析,构建出主成分函数表达式,计算出各丛状采伐样地综合得分,通过各丛状采伐样地恢复后小气候综合得分,来评价各采伐样地小气候优异程度。
以上分析均在SPSS 25.0中进行。
各系数计算公式如下[16]:
2 结果与分析
2.1 关键生态因子筛选
通过对小气候因子进行步进回归,最后得到进入模型的3个小气候因子为太阳光照强度、土壤温度和空气温度,得到回归模型(P=0.000,R2=0.996):
Y=32.740-0.38X1+0.122X2+0.9X3
式中:Y为新竹生物量,X1表示太阳光照强度,X2表示空气温度,X3表示土壤温度。由回归模型可知,采伐恢复后影响新竹生物量的主要因子是太阳光照强度、土壤温度和空气温度。
2.2 不同采伐方式对龙竹林内光照强度的影响
不同季节龙竹林内光照强度日变化如图1所示。随着太阳高度角的变化和日照时间的长短变化,春季、秋季、冬季的光照强度在12:00达到最大值,夏季的光照强度在13:00达到最大值,光照强度日变化呈低—高—低单峰曲线走势。4个季节内,全丛采伐下的林内光照强度最大(16 460~140 400 Lx)。在春季,1/3丛伐下的林内光照强度变化幅度不大,趋于平稳,常规采伐、1/2丛伐、全丛采伐下的林内光照强度变化幅度较大;在夏季,全丛采伐光照强度皆高于其他林分;在秋季,1/3丛伐在10:00至11:00小幅度的下降,之后开始上升,1/2丛伐12:00至13:00保持不变,之后开始减小,常规择伐和全丛采伐变化趋势基本一致;在冬季,常规择伐、1/2丛伐、1/3丛伐光照强度变化趋势基本一致,全丛采伐在12:00达到峰值后开始下降,在13:00至14:00呈小幅度上升,之后开始下降。
2.3 不同采伐方式对龙竹林林内土壤温度的影响
龙竹林内不同季节土壤温度差异如表2所示。全丛采伐样地土壤温度均高于其他3种采伐样地土壤温度,夏、秋季节常规择伐样地土壤温度最高。同一季节内不同采伐处理下的平均土壤温度差异显著(P<0.05),夏季和秋季常规择伐龙竹林内土壤温度皆比其他林分高,春季和冬季又以全丛采伐土壤温度最高。
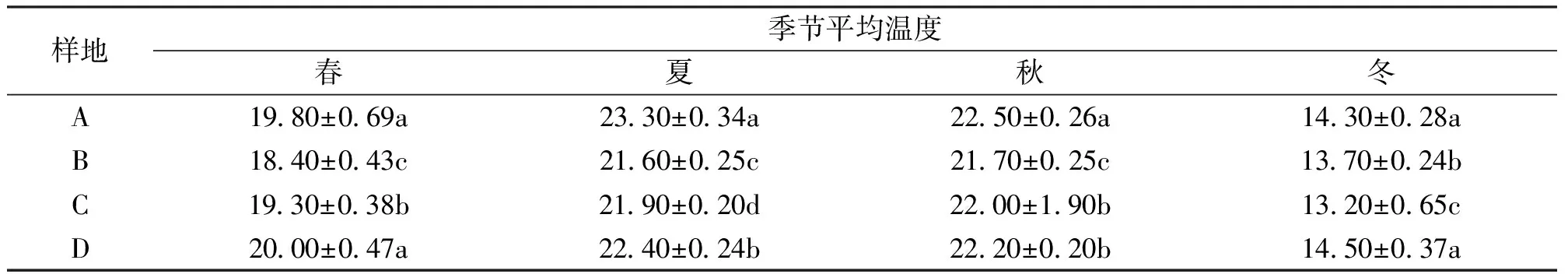
表2 样地土壤温度差异
2.4 相关性分析
各小气候因子相关分析结果如表3所示。光照强度和空气温度呈现显著正相关;空气湿度和光照强度、空气温度呈现负相关。风速和光照强度、空气温度呈现正相关,和空气湿度呈现极显著负相关。土壤温度和光照强度呈现正相关,和空气温度呈现极显著正相关,与空气湿度呈现负相关。生物量和发笋数呈现负相关,发笋数和光照强度呈现极显著正相关。结果表明光照强度越高,空气温度越高,发笋数越高;光照强度和空气温度越高,空气湿度越高;空气温度越高,土壤温度越高。光照和空气温度增高,空气湿度相对降低,但是影响不显著。

表3 各小气候因子相关性分析
2.5 采伐效果评价
将不同丛状采伐样地的各小气候因子和新竹生物量以及单位面积发笋数进行主成分分析,得到不同丛状采伐样地主成分方差贡献率(表4)。得到方差特征值大于1的主成分有3个,累计方差达到81.149%。主成分载荷矩阵见表5,主成分1包含的因素是温度、风速、和光照,主成分2包含的因素是空气湿度,主成分3包含的因素是土壤温度、生物量和发笋数。

表4 不同丛状采伐样地主成分方差贡献率

表5 不同丛状采伐样地主成分载荷矩阵
主成分1、2、3的函数表达式如下。
Y1=0.49X1+0.46X2+0.43X3-0.36X4+0.32X5+0.35X6-0.11X7
Y2=0.24X1-0.47X2+0.34X3+0.61X4+0.03X5+0.36X6-0.32X7
Y3=0.35X1-0.05X2-0.35X3-0.09X4-0.56X5+0.55X6+0.36X7
式中:Y1、Y2、Y3分别代表主成分1、2、3的特征成分权重;X1、X2、X3、X4、X5、X6、X7分别为空气温度、风速、光照强度、空气湿度、发笋个数、土壤温度和生物量标准化数据;Y为样地综合得分,Z1、Z2、Z3为主成分1、2、3中的主成分特征值。
将X1、X2、X3、X4、X5、X6、X7代入主成分1、2、3的函数表达式中得出Y1、Y2、Y3,再根据样地综合得分式计算出各样地综合得分(表6)。

表6 不同丛状采伐样地综合得分
不同丛状采伐样地综合得分如表6所示,1/2丛采伐得分最高,全丛采伐样地次之,其次是1/3丛采伐,最后是常规采伐。
2.6 采伐后恢复特征
不同丛状采伐样地新竹生物量如表7所示。不同丛状采伐处理间新竹生物量、发笋数、胸径、成竹数间皆显著(P<0.05)。1/2丛伐新竹生物量最大,其次是常规择伐,然后是1/3丛采伐仅次之,全丛采伐新竹生物量最小;发笋数全丛采伐皆高于常规采伐、1/2丛伐、1/3丛伐,但只比1/2丛伐高出1.6株/丛;胸径以1/3丛伐最大,全丛采伐的最小。全丛采伐丛间发笋数多,成竹数也多,丛间密度相对较大,相互竞争吸收养分,最终生物量较另外3种丛状采伐后新竹生物量要小。

表7 不同丛状采伐后恢复更新对比
3 讨论与结论
杨建祥等[17]认为在抚育采伐前单位面积林地上植物株数较多,林冠郁闭度大,森林内光照强度较差,经过采伐后,密度减小,林内光照强度增加。林内光照辐射量随着林下全天空系数的增加而增加,全天空系数对林下光照起着至关重要的作用[18]。叶冠开度大,叶面积指数小的林型林下光照强度强,林分郁闭度高,叶片浓密的林型林下光照强度弱[19]。因此,林内光照辐射会因林冠郁闭度、林冠开度、天空系数等因素的差异而发生变化。本研究结果显示不同采伐方式对龙竹林内光照辐射皆有影响,全丛采伐林内光照辐射响应最为明显。主要是因为4个样地对龙竹林不同程度的采伐,改变了龙竹林分密度以及冠层,导致林隙增大,林内光照强度增加,太阳光照强度达到显著性差异[20]。
土壤温度除受海拔、经度、纬度等因素影响外还受地表覆盖、土壤质地、土壤湿度等因素的影响[21]。郁闭度、冠长、冠幅等因子对40~60 cm深处的土壤温度影响最为显著,太阳辐射对土壤温度的影响越大,那么郁闭度、冠长冠幅等因子就越小,并且林分胸径和树高生长与土壤温度成负相关[22]。本研究显示,不同采伐强度处理下同一土层(50 cm)土壤温度之间差异显著(P<0.05),证明了采伐对土壤温度是有影响的;林冠冠幅减小,影响龙竹林郁闭度,光照强度增大,太阳光直射地面,太阳辐射对土壤温度的影响增大,这也是全丛伐胸径最小的原因。与温亮[23]对辽东山区人工红松(Pinuskoraiensis)林进行的小气候特征研究得到的2—3月间林内土壤温度开始低于林外,9—10月间林内土壤温度开始高于林外,以及杜颖等[24]对长白山阔叶红松林温度效应的研究得到的3—10月间林内的土壤温度低于林外的结果有所不同。春、冬季节,全丛采伐土壤温度皆高于常规择伐、1/2丛伐和1/3丛伐,夏、秋季节,常规采伐土壤温度皆高于1/2丛伐、1/3丛伐以及全丛采伐。可能是由于地理环境的不同和林分的差异造成的,龙竹地下茎发达,夏、秋季节属于龙竹生长季,春、冬季节龙竹生长速度减慢,地下根系不活跃,再加上枯枝落叶层对土壤热收支起阻隔作用,从而导致对土壤温度季节变化的影响有所差异。
曾宪礼等[3]在安徽省黄山区毛竹林带状采伐后恢复质量特征研究中得到最强采伐带处理下单位面积新竹生物量小于中度采伐处理的结果。在本研究中,龙竹林在全丛采伐这一高强度采伐后表现为新竹生物量最低,1/2丛伐区新竹生物量最大。适度采伐增加林内光照强度、空气温度和土壤温度等小气候因子,对新竹生物量有一定的影响,虽然全丛采伐时各小气候因子都是最大值,但是生物量却是最小的,这也是太阳光照强度与生物量呈负相关的原因;生物量最小,这可能是由于全丛采伐强度过大,前一年中新竹光合作用比较弱,没有留竹等原因所致,这种情况下小气候因子对新竹生物量的影响还是有限的。
综上所述,不同丛状采伐强度小气候的变化,对后期龙竹林的竹笋生长和龙竹林分更新以及新竹生长产生影响。从各样地综合评价得分以及新竹生长对采伐强度的响应及森林资源可持续利用与可持续发展等来看,1/2丛伐是最适宜的丛状采伐方式。
龙竹丛状采伐后恢复是一个长期的过程,本文仅初步研究了龙竹竹丛在不同采伐程度下经营一年林内小气候的变化以及新竹生长情况,采伐抚育间隔时间较短,龙竹林仍处于生长恢复期。所以,想要更科学的掌握不同丛状采伐强度对龙竹林下小气候变化规律以及对新竹生长变化的影响仍需进行长期的观测,收集更多数据进行研究分析,进而对建立高效龙竹林培育模式与可持续经营和科学管理模式提供更有利的参考。
猜你喜欢
农业灾害研究(2022年1期)2022-05-07
阅读(高年级)(2018年6期)2018-05-14
阅读(高年级)(2018年1期)2018-05-14
南方农业·下旬(2016年9期)2017-04-27
江苏农业科学(2017年5期)2017-04-15
科技与创新(2016年18期)2016-11-04
小天使·一年级语数英综合(2015年8期)2015-07-06
湖北农业科学(2014年21期)2014-12-22
小天使·一年级语数英综合(2014年6期)2014-07-22
青年文摘·上半月(1988年2期)1988-11-01
