
CDDP结合SRAP标记分析杧果品种的遗传多样性
2022-04-15 07:35:16黄国弟莫永龙罗世杏唐玉娟陆祖双
中国南方果树 2022年2期
张 宇,黄国弟,莫永龙,罗世杏,赵 英,唐玉娟,陆祖双,单 彬,荣 涛
(广西农业科学院/广西亚热带作物研究所,南宁,530001)
杧果MangiferaindicaL.多分布于干热河谷地区以及南北回归线之间,是药食两用型植物[1-2]。杧果产业发展依赖于杧果新品种的推陈出新,杧果新品种的产生依靠于多样性的杧果种质资源[3-7]。利用CDDP[8]、ISSR[9-10]、SRAP[11-12]、SCoT[13]、SSR[14]、AFLP[15]等分子标记技术对杧果品种进行遗传多样性和亲缘关系分析,可为杧果产业发展提供参考[16]。但每一种分子标记技术都存在局限性,所得到的结果往往带有片面性。利用DNA保守序列多态性(CDDP)结合相关序列扩增多态性(SRAP)对杧果品种进行遗传多样性分析,目前鲜见相关报道;前者是目的性标记,针对植物DNA的保守序列用单引物扩增目标分子的标记技术,后者是随机性标记,采用双引物对开放阅读框(ORFs)进行扩增,检测其多态性[17-20]。为避免单一分子标记以及同类型分子标记对试验结果造成偏差,笔者采用CDDP结合SRAP分子标记的方法对全球主要栽培杧果品种进行遗传多样性分析,为进一步拓宽杧果分子辅助育种以及种质鉴定提供数据信息支持。
1 材料与方法
1.1 材料与处理
供试材料来自广西亚热带作物研究所杧果种质资源圃(见表1)。供试材料处理、模板获取参考张宇等[21]的方法。引物由华大基因生物工程(深圳)股份有限公司合成(见表2)。

表1 35个供试杧果品种
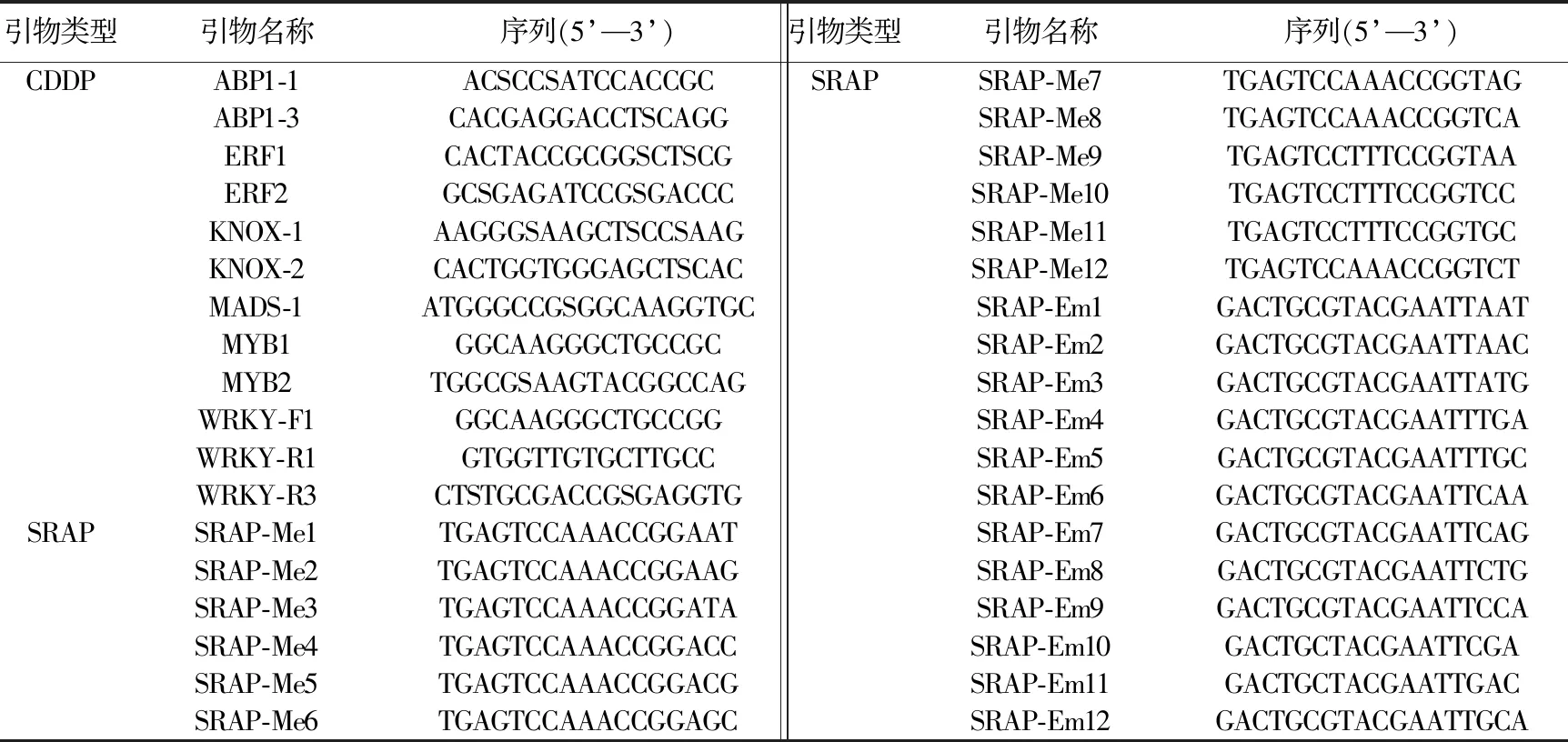
表2 CDDP和SRAP引物信息
1.2 CDDP-PCR与SRAP-PCR扩增及其产物检测
CDDP-PCR、SAP-PCR扩增反应程序和体系各组分用量参考张宇等[8,11]的方法。
1.3 数据信息分析
计算扩增条带数(TNB)、多态性条带数(NPB),多态性条带比率(polymorphism rate,PPB)、有效等位基因(effective allele number,Ne)数量、Nei’s基因多样度(Nei’s gene diversity,H)、Shannon信息指数(Shannon information index,I)和多态性信息含量(Polymorphism information content,PIC)等遗传多样性指标;聚类分析和主成分分析分别参考王丽媛等[22]和高源等[23]的方法。
2 结果与分析
2.1 杧果CDDP与SRAP的PCR扩增
查阅相关文献,选用表2中引物与供试材料DNA模板锚定,分别选取12条和12对CDDP和SRAP引物对35个杧果品种进行多态性分析。结果显示,SRAP和CDDP扩增总条带数分别为243条和263条,多态性条带数分别为230条和250条,多态性条带比率分别为94.61%和95.45%,平均多态性信息指数分别为0.59和0.60。说明CDDP标记所产生的总条带数、多态性条带数和多态性条带比率均高于SRAP标记。此外,两种标记的遗传多样性参数也有所差异,除平均有效等位基因数,SRAP标记为1.62略高于CDDP标记的1.59外,Nei’s基因多样性指数、Shannon’s信息指数和多态性信息含量,CDDP标记均高于SRAP标记。推断CDDP标记检测多态性的能力和标记效率高于SRAP标记。
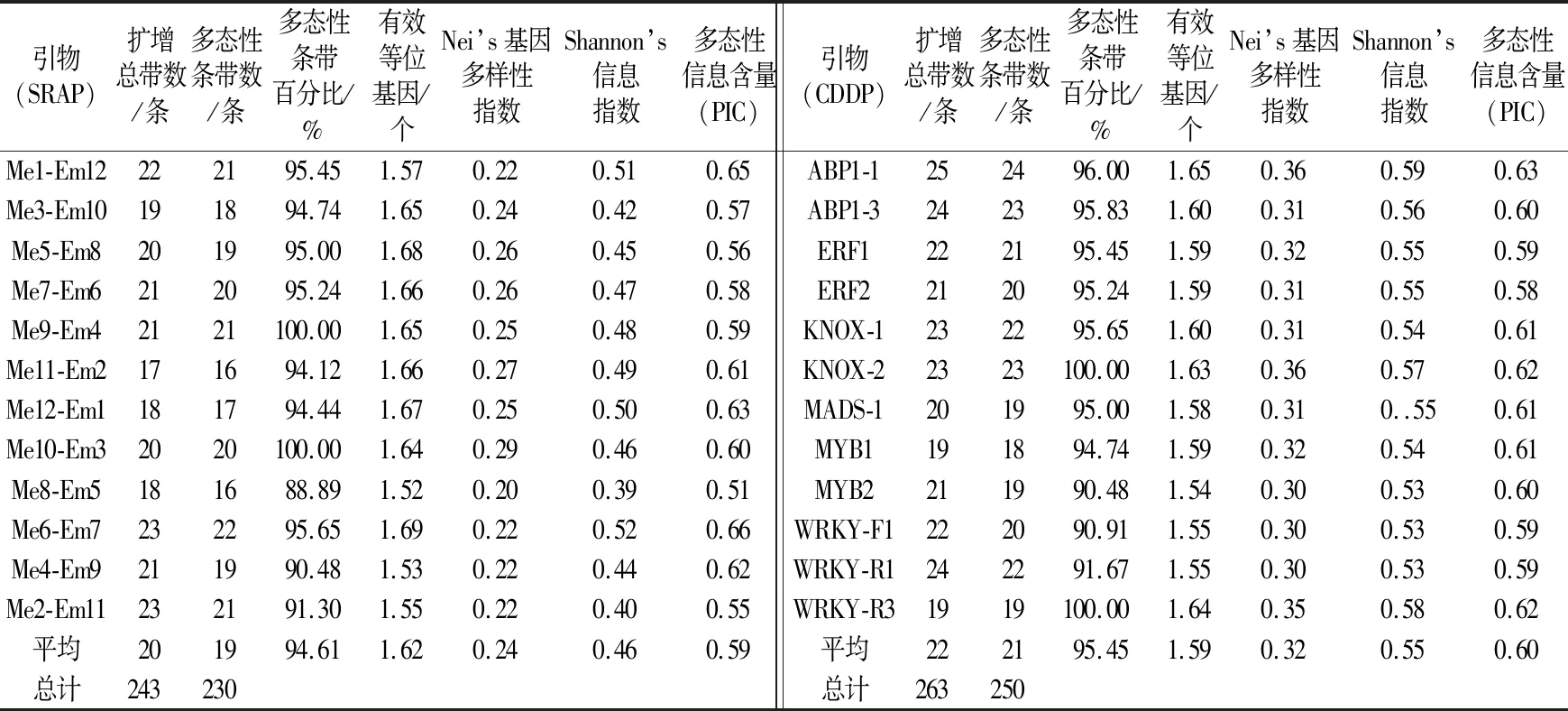
表3 35个杧果品种SRAP和CDDP标记遗传多样性参数
尽管本研究中CDDP标记和SRAP标记分别只选取了12条和12对,引物的数量与已报道的CDDP和SRAP标记引物不算多,且供试材料仅为35个杧果品种;但均能扩增出条带清晰,层次分明,完整性好,数目多的电泳条带,可以较理想地显示出杧果的遗传多样性(见图1),且上述两种标记结合分析杧果种质亲缘关系鲜见报道。
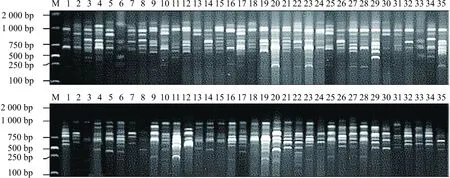
注:上图为ABP1-1引物的PCR扩增,下图为Me1和Em1引物的PCR扩增。M为marker,1~35表示不同杧果品种,详见表1。图1 35个杧果品种CDDP-PCR和SRAP-PCR
2.2 聚类分析
利用UPGMA软件对35个杧果品种进行聚类分析,CDDP标记的遗传相似系数分布在0.64~0.96,平均遗传相似系数为0.80。在遗传相似系数为0.709 5处,35个杧果品种分为A、B、C 3大类,A类为杉林1号、红象牙、农院红杧、黄象牙、吕宋、柳州吕宋、台农1号、凯特、爱文、圣心、R2E2、红凯特、肯特、热农1号、肯辛顿、金煌和台牙,B类为贵妃、玉文、凤凰杧、帕拉英达、金穗、水英达、农院8号、秋杧、紫花和沅江象牙,C类为桂香杧、串杧、桂热杧10号、桂热杧23号、桂热杧82号、桂热杧71号、水仙杧和秋杧。在相似遗传系数为0.763 5处,A类又可分为A1、A2和A3,共计3类,A1类为杉林1号、红象牙、农院红杧、黄象牙、吕宋、柳州吕宋和台农1号,A2类为凯特、爱文、圣心、R2E2、红凯特、肯特和热农1号,A3类为肯辛顿、金煌和台牙。在相似遗传系数为0.776 9处,B类又可分为B1和B2,共计2类,B1类为贵妃、玉文、凤凰杧、帕拉英达、金穗和水英达,B2类为农院8号、秋杧、紫花和沅江象牙。在相似遗传系数为0.818 5处,C类又可分为C1和C2,共计2类,C1类为桂香杧、串杧和桂热杧10号,C2类为桂热杧23号、桂热杧82号、桂热杧71号、水仙杧和秋杧。
聚类分析可知,SRAP标记的遗传相似系数分布在0.64~0.92,平均遗传相似系数为0.78。在遗传相似系数为0.694 7处,35个杧果品种分为A、B、C 3大类,A类为吕宋、黄象牙、爱文、农院红杧、红象牙、黄象牙、台农1号、杉林1号、柳州吕宋、凯特、R2E2、热农1号、肯辛顿、肯特、玉文、台牙和贵妃,B类为农院8号、球杧和凤凰杧,C类为圣心、红凯特、金煌、帕拉英达、桂香杧、紫花、沅江象牙、串杧、水英达、金穗、桂热杧82号、桂热杧71号、桂热杧23号、秋杧、水仙杧。在相似遗传系数为0.761 9处,A类又可分为A1、A2、A3和A4,共计4类,A1类为吕宋、黄象牙、爱文、农院红杧、红象牙、黄象牙、台农1号、杉林1号和柳州吕宋,A2类为凯特、R2E2和热农1号,A3类为肯辛顿、肯特和玉文,A4类为台牙和贵妃。在相似遗传系数为0.806 7处,B类又可分为B1和B2,共计2类,B1类为农院8号,B2类为球杧和凤凰杧。在相似遗传系数为0.746 9处,C类又可分为C1、C2和C3,共计3类,C1类为圣心、红凯特和金煌,C2类为帕拉英达、桂香杧、紫花、沅江象牙、串杧、水英达、金穗和桂热杧82号,C3类为桂热杧71号、桂热杧23号、秋杧和水仙杧。
聚类分析可知,CDDP+SRAP标记的遗传相似系数分布在0.62~0.94,平均遗传相似系数为0.78。在遗传相似系数为0.685 0处,35个杧果品种分为A、B、C、D、E 5大类,A类为红象牙、杉林1号、农院红杧、黄象牙、柳州吕宋、吕宋和台农1号,B类为爱文、凯特、圣心、红凯特、肯特、R2E2和肯辛顿,C类为热农1号、金煌和玉文,D类为台牙、贵妃、凤凰杧、农院8号、球杧、帕拉英达、水英达、金穗、紫花、串杧、沅江象牙、桂香杧、桂热杧10号和桂热杧82号,E类为桂热杧23号、桂热杧71号、秋杧和水仙杧。在相似遗传系数为0.734 8处,A类又可分为A1和A2,共计2类,A1类为红象牙、杉林1号和农院红杧,A2类为黄象牙、柳州吕宋、吕宋和台农1号。在相似遗传系数为0.730 6处,B类又可分为B1和B2,共计2类,B1类为爱文、凯特、圣心、红凯特和肯特,B2类为R2E2和肯辛顿。在相似遗传系数为0.827 9处,C类又可分为C1和C2,共计2类,C1类为热农1号和金煌,C2类为玉文。在相似遗传系数为0.756 1处,D类又可分为D1、D2和D3,共计3类,D1类为台牙、贵妃、凤凰杧、农院8号和球杧,D2类为帕拉英达、水英达和金穗,D3类为紫花、串杧、沅江象牙、桂香杧、桂热杧10号和桂热杧82号。在相似遗传系数为0.785 9处,E类又可分为E1和E2,共计2类,E1类为桂热杧23号,E2类为桂热杧71号、秋杧和水仙杧。
本研究对CDDP标记、SRAP标记以及CDDP+SRAP标记对杧果品种遗传多样性进行聚类分析,不同标记聚类分析结果不同,CDDP+SRAP标记聚类分析结果较单独标记聚类分析更细化,但基于分子标记技术的聚类分析结果与杧果原产地无相关性。

注:Ⅰ、Ⅱ、Ⅲ分别为CDDP标记、SRAP标记以及CDDP+SRAP标记聚类分析。编号1~35表示不同杧果品种,详见表1。图3同。图2 35个杧果品种CDDP和SRAP聚类
2.3 主成分分析
基于CDDP+SRAP标记对35个杧果品种的遗传多样性分析结果绘制主成分分析平面PCA图(见图3)。图3中编号分布疏密代表品种之间的遗传关系,对比聚类分析结果,其亲缘关系有相似之处,也有很大不同。聚类分析表明红象牙、杉林1号、农院红杧、黄象牙、柳州吕宋、吕宋和台农一号聚为一类,显示出较其他供试品种较近的亲缘关系;但主成分分析表明,红象牙、杉林1号和农院红杧亲缘关系较近,黄象牙、柳州吕宋、吕宋和台农一号亲缘关系较近,而二部分之间的亲缘关系较远。聚类分析结果表明,爱文、凯特、圣心、红凯特、肯特、R2E2和肯辛顿聚类为一类,显示出较其他供试品种较近的亲缘关系;但主成分分析表明,爱文与台牙、贵妃和凤凰杧的亲缘关系紧密,凯特与热农1号之间的亲缘关系紧密,红凯特和肯特与球杧、帕拉英达的亲缘关系紧密。聚类分析不能较好地区分出原产地一致的杧果品种,但主成分分析可以将一些来源地不一致的杧果品种区分出来。这很有可能是因为不同的分析方法采用的分析软件不同、考量的区分因素不同;即使存在相同的考量因素,但是侧重点不同,因此导致分析结果不同。对比两种遗传多样性的分析方法可知,主成分分析可以更加直观立体地表明杧果品种之间的遗传疏离、远近关系,即能体现聚为一类品种之间的亲密关系,也可以反应不同类别之间的离散程度。

图3 35个杧果品种的主成分分析
3 结论与讨论
杧果新品种的选育依赖于杧果丰富的遗传多样性[24-26]。杧果高度异花授粉,种子高度杂合,遗传背景复杂,传统的单一分子标记已经不能准确地分析其遗传多样性,采用多种分子标记结合的方式从DNA水平反映遗传变异,对杧果遗传育种研究具有重要意义。本研究采用CDDP结合SRAP标记对35个杧果品种进行了遗传信息数据分析、聚类分析和主成分分析。试验结果表明,除平均有效等位基因数,SRAP标记略高于CDDP标记外,多态性条带比率、Nei’s基因多样性指数、Shannon’s信息指数和多态性信息含量均为CDDP标记高于SRAP标记。由此判断,CDDP标记检测多态性的能力和标记效率高于SRAP标记。尽管本研究中只选取了已报道CDDP和SRAP标记引物的小部分,且供试样本量仅为35个杧果品种,但遗传信息数据表明CDDP标记和SRAP标记均可较理想地显示出杧果的遗传多样性。
对35个杧果品种进行聚类分析和主成分分析反映出的品种间亲缘关系有相似之处,但也有很大不同。采用CDDP标记、SRAP标记以及CDDP结合SRAP标记分别绘制聚类分析图,依次在遗传相似系数0.694 7、0.709 5和0.685 0处,35个杧果品种分别分为A、B、C三大类,A、B、C三大类,A、B、C、D、E五大类,由此可以判定CDDP结合SRAP标记较CDDP和SRAP标记可以将供试杧果品种分类更全面细化。3种标记方法又可以分别分为不同的小类,将亲缘关系更紧密的杧果品种聚集在一起。通过聚类分析可知CDDP、SRAP以及CDDP结合SRAP标记得出聚类分析结果不相同,如CDDP结合SRAP和CDDP标记聚类分析可以较好地体现出吕宋杧和柳州吕宋的密切亲缘关系,而SRAP标记虽然也可以将吕宋杧和柳州吕宋聚集在A1类,但遗传亲缘关系上却显得没有那么密切,可能是由于CDDP和SRAP标记的技术原理不同造成聚类结果不同。SRAP引物较为随机,没有针对目的基因片段序列进行锚定,随机性较强;而CDDP标记虽然其引物数量不多,但其引物设计是依据植物材料中特有的功能基因以及基因家族,目的性较强;而CDDP结合SRAP标记,极大地吸收了两种标记的优点,将随机性和确定性紧密地结合在一起,可以较大程度弥补单一标记的不足,提高分析结果数据的可靠性。聚类分析结果显示,紫花杧和金穗杧的遗传相似系数偏大,很可能是因为CDDP和SRAP标记引物可以较理想地锚定两者之间的基因差异序列,表现出极为显著的遗传特异性,反应出二者之间较大的遗传距离。主成分分析可知,爱文与台牙、贵妃和凤凰杧的亲缘关系紧密,凯特与热农1号之间的亲缘关系紧密,红凯特和肯特与球杧、帕拉英达的亲缘关系紧密。但聚类分析不能显示出上述品种间的紧密亲缘关系,只能将亲缘关系较为接近的杧果品种聚为一类,且不能显示出聚为一类的杧果品种间的空间遗传距离,以及不同聚类品种间的遗传差异。比较聚类分析图和主成分图可知,主成分图更加立体,直观地反映杧果品种间的遗传疏离、远近、分布情况,而聚类分析图无法达到前者的显示效果。这很有可能是由于不同分析方法采用的分析软件不同、考量的区分因素不同,即使存在相同的考量因素,但是侧重比例不同。总体来说,主成分分析既能体现聚为一类品种之间的亲密关系,也可以反应不同类别之间的离散程度。聚类分析不能区分出原产地一致的杧果品种,但主成分分析可以将一些来源地不一致的杧果品种区分出来。
猜你喜欢
青少年科技博览(中学版)(2023年5期)2023-06-26 09:39:44
海洋通报(2022年6期)2023-01-07 02:48:56
作物学报(2022年2期)2022-11-06 12:11:02
中国现代中药(2021年9期)2021-11-16 07:43:32
湘潮(上半月)(2021年3期)2021-07-20 07:27:50
源流(2020年6期)2020-08-03 01:59:35
作文·小学低年级(2019年2期)2019-09-10 07:22:44
传播力研究(2019年1期)2019-03-20 11:28:14
天然产物研究与开发(2018年10期)2018-11-06 07:43:46
广东农业科学(2017年5期)2017-08-29 10:37:26
