
微生物诱导白云石沉淀研究进展及面临的挑战
2022-04-02 08:08赵东方谭秀成罗冰王小芳乔占峰罗思聪
沉积学报 2022年2期
赵东方,谭秀成,罗冰,王小芳,乔占峰,罗思聪
1.西南石油大学天然气地质四川省重点实验室,成都 610500 2.中国石油集团碳酸盐岩储层重点实验室西南石油大学研究分室,成都 610500 3.中国石油西南油气田公司勘探开发研究院,成都 610041 4.中国石油杭州地质研究院,杭州 310023
0 引言
白云岩自首次被描述以来[1],其成因机理和油气矿产意义[2-4],一直是地质学家们关注和争论的焦点。多年以来,虽然“白云岩(石)问题”的内涵不断地被学者们补充、完善和总结[5-7]。但其核心内容主要包括以下两个方面:1)白云石在地质历史时期中的分布是不均衡的[8],其广泛分布于古老地层中,而鲜见于全新世沉积物;2)低温条件下(大致25 ℃~60 ℃[9]),如果不存在微生物参与,实验室内难以沉淀“有序的”白云石[10]。针对上述问题,前人提出了多种白云石化机理模式,如渗透回流模式[11]、萨布哈模式[12]、混合水模式[13]、埋藏白云石化[14]、拟晶白云石化[15-16]、构造—热液模式[17]等等。实际上,Neheret al.[18]很早就认识到微生物活动对沉淀白云石的潜在贡献,但在当时并未引起学界的重视。Vasconceloset al.[19]通过实验模拟潟湖环境,在硫酸盐还原细菌的参与下,成功沉淀出与自然界潟湖特征类似的白云石,进而提出微生物诱导沉淀白云石模式。此后的二十余年内,该模式快速地扩展与延伸[20-26],成为一种低温条件下沉淀白云石的重要路径。本文在回顾和梳理微生物诱导白云石机理和特征的基础上,重点总结其局限性和面临的挑战,以期更好地审视微生物诱导模式和为“白云岩(石)问题”提供新的思考和理论依据。
1 微生物诱导白云石沉淀的机理
虽然现今很多水文环境,如海水、部分湖水和地下水等,通常对于白云石而言都是过饱和的[27],但白云石在现代环境中的分布依然十分有限[8]。大量的研究实例都表明,动力学壁障是制约白云石难以在地表低温条件下沉淀的主要原因[10]。主要表现在以
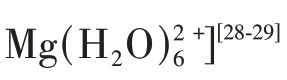


图1 特定官能团破坏Mg2+水合作用的能量壁垒[45]Fig.1 Specific functional groups can break the energy barrier of Mg2+hydration[45]
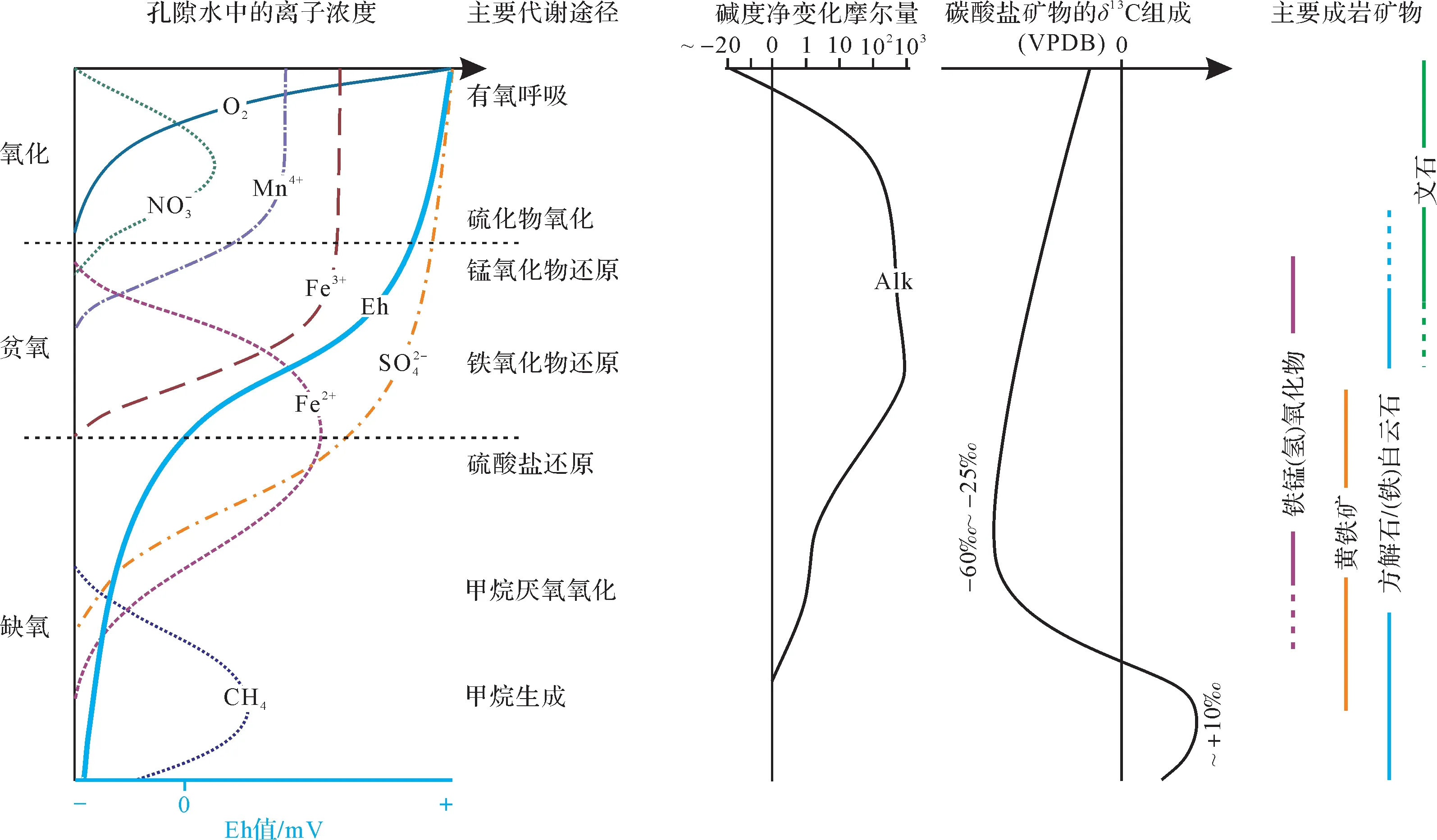
图2 微生物席内的主要功能反应、水化学性质及主要成岩矿物的纵向演化[46]Fig.2 Vertical evolution of functional reactions, hydrochemical properties, and major diagenetic minerals in microbial mats[46]
随着现代观测和相关模拟实验的开展,多种微生物都被证实对白云石的沉淀有积极的促进作用。按照微生物新陈代谢方式的不同(表1),主要包含四类[46]:好氧异养型(aerobic heterotrophy)、化能硫化物氧化型(chemotrophic sulfide oxidation)、异化硫酸盐还原型(dissimilatory sulfate reduction)、甲烷生成型(与甲烷的厌氧氧化相联合,Methanogenesis coupled to anaerobic oxidation of methane)。总体而言,随着研究的深入,诱导白云石模式中涉及微生物的运动方式(包括底栖和浮游微生物)和新陈代谢类型(包括好氧型和厌氧型)都愈发多样,其相关的水文化学参数(淡水、海水和咸水)也愈发丰富。
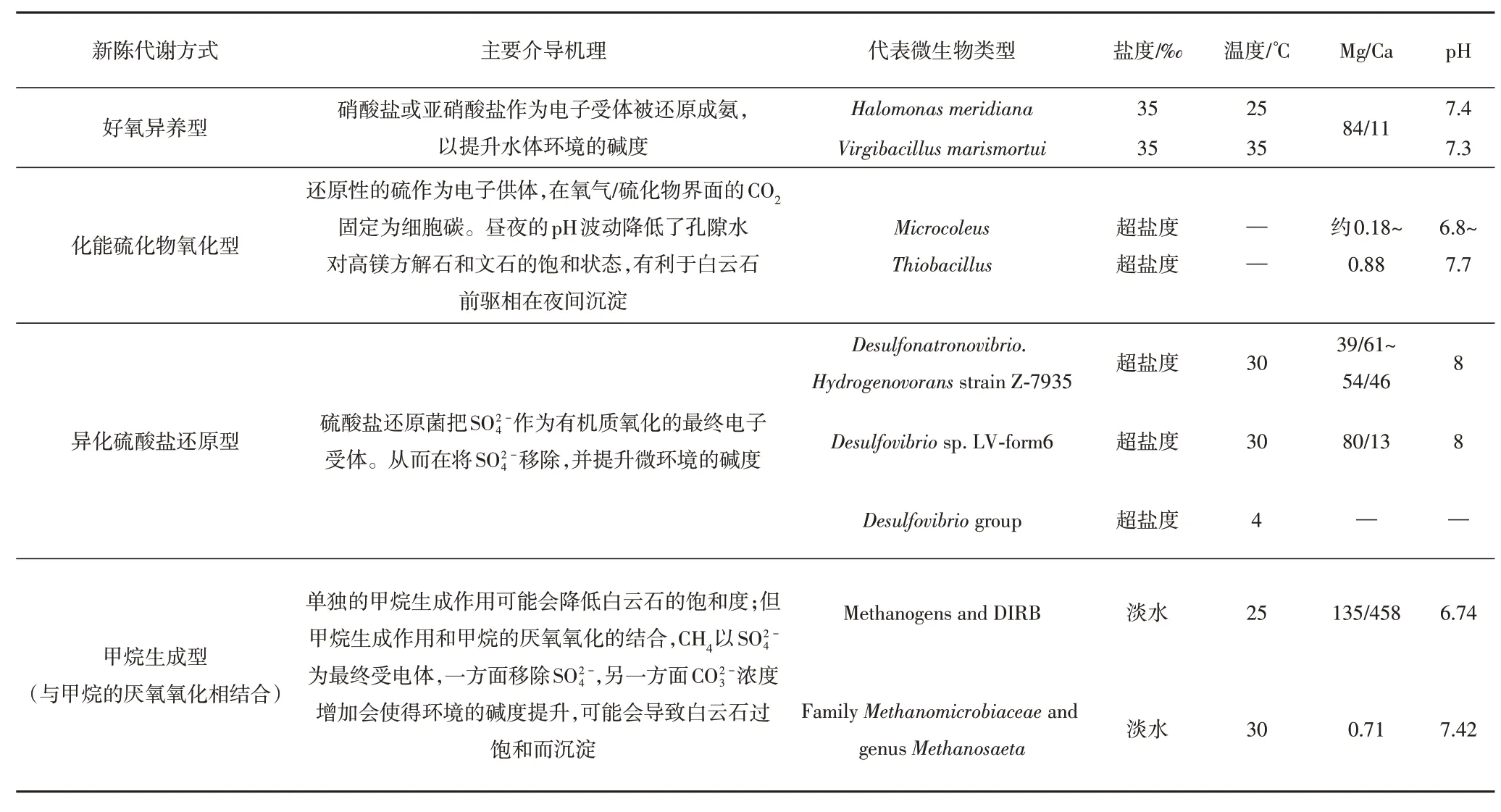
表1 诱导白云石沉淀的微生物新陈代谢类型及相关条件参数(据由雪莲等[22]; Petrashet al.[46])Table 1 Metabolic types of microbes and environmental parameters of microbial dolomite(after Youet al.[22]and Petrashet al.[46])
2 微生物诱导沉淀白云石的潜在识别标志
针对微生物白云石,前人总结了很多潜在的识别标志。但需要注意的是,大部分识别标志都是基于现代观察和室内培养实验总结得出的。对于古老样品而言,微生物成因白云石的识别依然具有挑战性。主要表现在:1)球粒、哑铃状等特殊的微观形貌并不是判识微生物成因的可靠标志;2)缺乏强力的地球化学特征约束;3)古老白云岩经历了复杂的成岩作用,多期次的溶解—重结晶作用,使得其原始形貌和地化信号大多被破坏和覆盖,晶形趋于呈现菱面体。
2.1 微观形貌特征
生物诱导矿化过程中,有机大分子对矿物的形成具有明显的影响[49]。大量的实验观测表明,微生物诱导形成的白云石往往呈特殊的微观形貌,如球粒状、哑铃状、花椰菜状、纳米微球粒集合体等[19,21],并常可见残余的EPS(图3)。这主要是由于在矿物生长的过程中,有机分子附着于特定的晶面上,从而产生“毒害效应”,抑制了相关晶面的生长,进而形成多样的晶体形态[48]。实际上,正是由于这些特殊的矿物形貌与微生物存在相似之处,且常以链状或簇状集合体产出,最初曾一度被认为是“纳米细菌”(nannobacteria)[50]或“纳米化石”(nanofossils)[51]。随着相关研究的深入,目前上述特殊微观形貌特征更趋于被解释成相关细菌经过脱水、分解、成岩压实收缩后的残骸化石或EPS降解矿化后的残余[52]。

图3 扫描电镜下可能的微生物诱导“白云石”典型特殊微观形貌(a)叠层石白云岩中的球粒状形貌,灯四段,磨溪8井,5 110.4 m;(b)凝块石白云岩中的花椰菜状矿物结合体,灯四段,磨溪102井,5 197.53 m;(c)叠层石白云岩中的哑铃状形貌和EPS残片,灯四段,磨溪105井,5 350.57 m;(d)凝块石白云岩中的纳米微球粒集合体,灯二段,康家坪剖面;(e)叠层石白云岩中的纳米微球粒,灯四段,磨溪105井,5 349.37 m;(f)凝块石白云岩中的EPS残片,灯四段,磨溪8井,5 118.23 mFig.3 Typical microscopic morphologies of potential microbial dolomites under scanning electron microscope (SEM)(a)spherulite in stromatolitic dolomite,the 4thmember of the Dengying Formation(Z2dn4),well MX8,depth 5 110.4 m;(b)cauliflower-like mineral aggregates in thrombMoXl i t1i c0 5d,o ldoempitthe ,5Z32d5n04.5,7 w eml l;M(d X)1 na 0n2o,m de e p te trh s5p h1e9r7o.i5d3s m in; t(hcr)odmubmo b li e ti lcl sdaonl o dmf r it aeg,mt h een t2snodfmeexmt ra b ceer lol uf atrh ep oDl yemn g eyt ri incg sFuobrsmt aant i coens ((ZE2PdSn)2)i,nKsat rnogm jia apt oilnigt i cS edcotlioomn;it(ee,)Zn2adnno4m,w et eelrl spheroids in stromatolitic dolomite,Z2dn4,well MX105,depth 5 349.37 m;(f)EPS fragments in thrombolitic dolomite,Z2dn4,well MX8,depth 5 118.23 m
2.2 同位素特征
2.2.1 碳同位素特征
白云石形成过程中,微生物活动会导致碳同位素发生较为明显的分馏[53]。研究表明,由于生物优先利用12C,使得有机质降解过程中生成HCO-3的δ13CVPDB约为-25‰[54]。正是由于这部分碳源的加入,与微生物有氧呼吸作用和硫酸盐还原作用相关的白云石,其δ13CVPDB会在-25‰~0 范围呈现不同程度的负偏[47,55]。同理,生物成因的甲烷以极负的碳同位素组成为特征(δ13CVPDB约为-60‰)[56],由于甲烷厌氧氧化作用等过程相关的白云石的部分碳源于上述甲烷,导致白云石的δ13CVPDB大多介于-60‰~-25‰[53]。根据平衡原理,生物甲烷生成时,甲烷中13C相对贫乏,相应地,13C 则在HCO-3中富集[56]。这部分富13C 的HCO-3是甲烷生成作用下形成白云石的重要碳源,故这些白云石往往呈现较为显著的碳同位素正偏(δ13CVPDB介于0‰~+26‰,多数集中在10‰左右)[20]。综上,在生物同位素分馏的影响下,随着碳源成因不同和混入比例的差异,微生物诱导形成白云石的δ13CVPDB可以在-60‰~+26‰之间波动(图2)。
需要注意的是,上述碳同位素异常的特征其实更多的表现在有机成因白云石(organogenic dolomite)中。严格意义上而言,有机成因白云石和微生物白云石(microbial dolomite)是有差异的。前者强调白云石的碳源本质上源于有机质,其形成时间宽泛,涵盖(准)同生期至埋藏期,因此有机成因白云石既可以是原生的,也可能是次生的;而微生物白云石更多是指微生物参与或影响下的同生期产物,且其碳同位素也一般不表现出异常特征。随着研究的深入,微生物的生理活性被证实并不是低温条件下催化白云石沉淀的必需因素,即使在微生物死亡或者其新陈代谢不活跃的情况下,部分有机大分子(尤其是富羧基的大分子)依然可以催化沉淀白云石[42,57]。基于此,部分学者不再严格区分有机成因白云石与微生物白云石,趋于将两者归为“微生物催化白云石形成”的范畴[46]。相应地,“微生物白云石模式”也进一步拓展为“微生物(有机)白云石模式”。
2.2.2 非传统同位素特征
(1)钙同位素。钙有6 种稳定同位素(40Ca、42Ca、43Ca、44Ca、46Ca 和48Ca),其 中40Ca 和44Ca 丰度 最高[58]。微生物诱导碳酸盐岩矿物沉淀过程中,由于EPS的存在以及微生物活动造成的微环境化学性质的差异,使得钙同位素产生不同程度的分馏[59]。Krauseet al.[41]利用Desulfobulbus mediterraneus的相关培养实验,证实了EPS及其诱导白云石中趋于富集轻钙(40Ca),并提出上述分馏过程大致分为两个阶段,即Ca2+与微生物膜的结合阶段和Ca2+进入白云石晶格阶段。随后,Krauseet al.[60]进一步利用Alcanivorax borkumensis及其诱导矿物,证实微生物活动造成的δ44Ca 分馏大致为(-1.19±0.22)‰,比非生物成因碳酸盐矿物[61-62]分馏更明显。此外,Bradburyet al.[63]的研究表明细菌的生长速率也是影响矿化产物中钙同位素值的重要因素,微生物的生长速率越快,钙同位素分馏越明显,相应的诱导矿物的δ44Ca越偏负。鉴于微生物生长速率受水体盐度、温度、pH和营养元素浓度等多种因素的影响,因此,生化过程中钙同位素的分馏机制依然较为复杂。
(2)铁同位素。铁有54Fe(5.84%)、56Fe(91.76%)、57Fe(2.12%)和58Fe(0.28%)这4 种稳定同位素[64]。研究发现,生物作用普遍选择性吸收和利用轻铁[65],进而对铁同位素分馏有明显影响,并对现代表层海水溶解铁中趋于富集56Fe有重要贡献[66]。但即便如此,部分研究实例表明,生物诱导矿化过程中的铁同位素分馏过程可能是复杂的过程,并不能用“生物优先富集轻铁”来简单概括。如,生物成因的菱铁矿[67]和全新统的微生物碳酸盐岩[68]中的铁同位素几乎没有发生分馏现象。这可能是由于生物矿化过程中铁同位素受到沉积环境的氧化还原条件[69-70]、微生物种类[71]、甚至是诱导矿物的化学组成[72]等多种因素的影响。
(3)镁同位素。镁有24Mg、25Mg 和26Mg 这3 种稳定同位素,其相对丰度分别为78.99%、10% 和11.01%[73]。目前,镁同位素已经成功应用于白云岩成因研究[74-76]。但对于微生物成因白云石,还处于镁同位素特征的简要表征阶段[77-78],尚缺乏系统报道。虽然,Mavromatiset al.[79]的实验结果表明,蓝细菌参与下沉淀的碳酸盐岩矿物与非生物成因的矿物间存在类似的镁同位素分馏现象,即蓝细菌的参与对镁同位素的分馏作用没有影响。但实际上,大量的研究都表明,生物矿化过程中,矿物与流体间的镁同位素分馏机制十分复杂[76],尚未达成共识。截至目前,生物种属[80]、矿物类型[80]、温度[81]、沉淀速率(有争议)[82-83]、无定型前驱的转化[84]、生物利用镁元素的方式[85]等因素都可能会影响生化过程中的镁同位素分馏。
3 微生物诱导白云石沉淀面临的挑战及其局限性
3.1 特殊形貌并不是判识微生物成因的可靠标志
虽然,微生物诱导的白云石往往呈特殊的微观形貌,但大量的证据表明,这些特殊形貌并不是微生物活动的特有产物。Kirklandet al.[86]指出,非生物过程也可以形成与微生物成因白云石类似的形貌特征。Velimirov[87]基于生物学角度,认为部分特殊形貌缺乏生物成因的有效支撑,原因在于其与微生物的尺寸和生理响应等方面存在诸多矛盾。此外,Gallagheret al.[88]发现,多数微生物培养皿中的磷酸盐含量远高于自然环境中的常值,其可能导致结晶产物呈不规则形态。因此,所谓的生物成因的特殊形貌完全有可能是溶液中高浓度磷酸盐形成的人为假象[88]。
实际上,诸如球粒、哑铃状等形貌更多地反应了晶体结晶过程的多阶段性,即经历了由不定型前驱体以球晶生长(Spherulitic growth)的结晶过程[89-90](图4),而与是否存在生物的参与没有必然联系。球晶生长过程多见于高分子和仿生领域,是一种典型的由动力学控制的非经典结晶路径[91]。对碳酸盐岩体系而言,当溶液呈高饱和度,或无定型前驱体与结晶相之间存在很大的溶解度差异时,往往会促进球晶生长[90]。截至目前,已有大量碳酸盐岩矿物可以在相关条件下由球晶生长过程产出,如球霰石[92]、文石[93]、方解石[94]、单水方解石[95]、白云石[96]和水菱钇矿等[97]。

图4 由无定型碳酸钙经球晶生长结晶生成方解石过程的扫描电镜照片[89](a)无定型碳酸钙;(b)无定型碳酸钙和球霰石纳米集合体;(c)无定型碳酸钙和球霰石纳米集合体;(d)球霰石纳米集合体;(e)球霰石纳米集合体;(f)球霰石纳米集合体,方解石晶体首现;(g)球霰石纳米集合体和方解石表面的生长阶步;(h)方解石表面的生长阶步;(i)球霰石纳米集合体和方解石表面的生长阶步Fig.4 SEM images of calcite transformed from amorphous calcium carbonate through spherulite growth and crystallization[89](a) amorphous calcium carbonate (ACC); (b) ACC and vaterite nanoaggregates; (c) ACC and vaterite nanoaggregates; (d) vaterite nanoaggregates; (e) vaterite nanoaggregates;(f)vaterite nanoaggregates and first calcite crystals;(g)calcite crystals attached to vaterite spheres with the development of growth steps on the calcite surface;(h)calcite growth steps;and(i)calcite crystals with vaterite casts
3.2 地球化学特征缺乏有力约束
针对微生物白云石,尽管学者们做了大量的地球化学工作,并尝试利用非传统同位素示踪其形成过程。但总体而言,微生物白云石的地球化学特征依然缺乏强有力的约束。前文已经述及,一方面,微生物白云石并不一定都呈现出异常的碳同位素特征。另一方面,Mg、Ca、Fe等非传统同位素在微生物白云石形成过程中应用尚不成熟,其分馏机制和相关细节还存在争议,且缺乏确定性的研究实例。此外,古老地层的白云岩样品必然要面临成岩作用对目标地球化学信号的破坏与覆盖,这极大地限制了地球化学手段在地质历史时期的微生物白云石研究中的运用。
3.3 “白云石”并不是微生物诱导的唯一产物
微生物席(microbial mat)是由微生物群落(多以蓝细菌为主)及其黏结—代谢产物构成的层状生态系统[98],其厚度介于数毫米至数厘米[99-100](图5)。微生物席垂向分异明显:1)多呈双层结构,上部以丝状菌为主,具有较强的韧性,起保护作用,下部主要为球状菌,较为致密,起稳固作用[101];2)微生物群落常垂向分布,从上至下,依次为光合自养细菌、硫酸盐还原细菌、化能自养细菌、硫化细菌等[102]。正是如此,微生物席内部的生化环境梯度变化非常剧烈(图2),相应的生物矿化产物类型多达数十种[38,103]。在这样的背景下,“白云石”仅是微生物诱导沉淀矿物的类型之一,这必然引发学者们对微生物诱导沉淀“白云石”的数量问题的思考,以及该模式能否解释地质历史时期规模性白云岩的疑问。当然,微生物白云石绝不仅限于微生物席内,正是目前缺乏更有效的识别标志,且后期成岩作用的改造与覆盖,很多古老微生物白云石可能未被识别[104]。这使得微生物作用对于古老白云岩的贡献还难以得到有效评估。

图5 巴西LagoaVermelha 潟湖半固结的微生物席[100],其中白色亮层为碳酸盐矿物层Fig.5 Semi-solid microbial mat from the LagoaVermelhalagoon in Brazil[100](white bright layers consist of carbonate)
3.4 现今浅水白云石主要分布在盐度较高的温暖环境
虽然深海沉积物中也有白云石分布的报道[105],但现代浅水白云石主要分布在温暖(主要分布在纬度40°S~40°N)、较高盐度的受限环境中(如潟湖、海湾和咸湖等)[106-125](图6),而在微生物席更为发育的潮坪环境中白云石却相对匮乏[126]。这可能暗示,水化学条件和水文学条件才是影响白云石沉淀的关键。
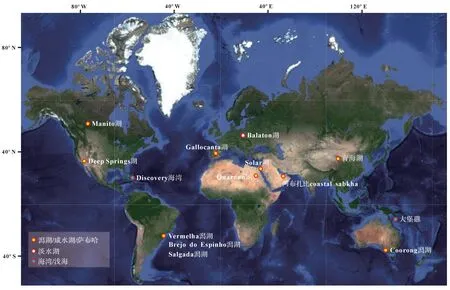
图6 现代白云石的全球主要分布(注洞穴堆积物和深海环境中的白云石并未统计)白云石统计数据:Manito湖[106-107],Deep Springs湖[108],Discovery海湾[109],Vermelha潟湖和Brejo do Espinho潟湖[110-111],Gallocanta湖[112-113],Quaroun湖[114],Balaton湖[115-116],Solar湖[117],阿布扎比coastal sabkha[118-120],青海湖[121-122],Coorong潟湖[123-124],大堡礁[125]Fig.6 Global distribution of modern dolomite (note: dolomite in cave deposits and deep sea is not counted)
实际上,较高的盐度对微生物诱导白云石沉淀有显著的促进作用[121]。微生物适应高盐环境的关键是维持细胞内外的渗透平衡[127]。研究表明,受高渗透压胁迫,微生物的EPS生成量将会加强,这是因为EPS作为细胞和环境间的过渡屏障,其表面的多种官能团能捕获阳离子,减缓外部离子向细胞内流入[128]。这种现象已经在多种微生物中得以证实[129-131]。此外,相对于其他蛋白质,富羧基的蛋白质在活性和稳定性都更具有优势[132],因此合成更多富羧基大分子是微生物适应高盐环境另一个重要途径。前文已述及,羧基可以有效降低Mg2+的水合作用,促进白云石生成。综上,较高盐度环境不仅能促进微生物分泌更多的EPS,也能增加细胞表面的羧基密度[121],进而大大促进了白云石在低温下成核。
3.5 地质历史时期,白云岩丰度的变化与微生物的分布并不完全匹配
白云岩的地史分布不均衡,其主要分布于元古宙,尤其是新元古代[133]。但在地质历史时期的演变中,白云岩的丰度变化与微生物席的分布并不完全匹配,即在微生物最为发育的古元古代和中元古代,白云岩的丰度并不大,相反在微生物已经受到一定胁迫的新元古代白云石丰度达到最大(图7)[133-134]。值得注意的是,白云岩的丰度在新元古代达到顶峰,恰好与古海洋学家提出的“白云石海”(dolomite sea,海水的Mg∕Ca比可能大于10)的时代分布具有很好的耦合性[135-137]。最近,Dayeet al.[26]利用厌氧光合细菌成功诱导出低有序度的白云石,但上述过程必须要求一定数量Mn2+的存在,而海水富Mn正是新元古代白云石海的典型特征之一[137-138],这也表明水化学性质的长周期变化可能对于白云石在地质历史时期中的不均衡分布有重要贡献。
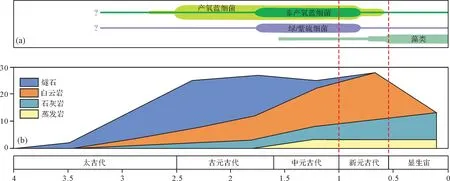
图7 地质历史时期的初级生产者演化[134]和非硅酸盐[133]的相对丰度变化Fig.7 Evolution of primary producers[134]and the relative abundance of non-siliciclastic sediments[133]through geologic history
地质学家已经认识到仅仅凭借成岩过程中的白云岩化作用似乎很难解释地质历史时期白云岩的非均衡分布状况[139]。在长周期尺度上,白云岩的规模性发育与全球环境的长期变化存在联系[140]。大量的统计资料和有关模拟结果显示,地质历史时期,白云岩主要分布在全球海平面上升背景下的浅水碳酸盐岩台地繁盛期,且大多集中在海水相对贫氧的时期[140-142]。目前,越来越多的证据表明,前寒武时期(尤其是新元古代)微生物白云石发育的环境可能比现今广泛的多,其微生物诱导白云石的规模也可能相当巨大[103]。但毫无疑问,上述推断尚处于探索阶段,还需要更多的证据和深入的研究来予以支持。
3.6 绝大部分微生物诱导沉淀的白云石,依然是严格无序的
从矿物学角度而言,所谓理想有序的白云石要满足以下两个基本条件:1)元素成分上,Ca2+∶Mg2+≈1;2)晶体结构上,阳离子完全有序占位,Ca2+层与Mg2+层以CO2-3层为间隔,依次交替排列,互不干扰,呈现典型的R-3空间群对称型[7,143]。基于上述特征,理想有序的白云石在X射线衍射图谱上理应具备两个重要的鉴别标志(图8):1)d104=2.88Å,这意味着Mg2+含量为50%;2)出现(101)、(015)和(021)等超结构衍射线,又以(015)峰最具说服力,标志着阳离子的有序排列。
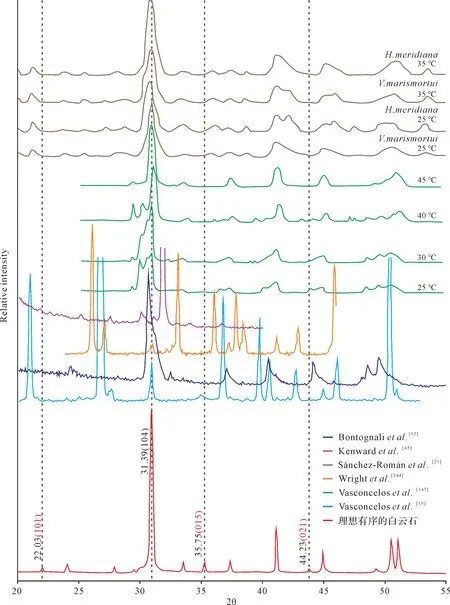
图8 理想有序白云石与微生物诱导沉淀“白云石”的XRD 衍射图谱特征对比Fig.8 Comparison of X-ray diffraction patterns between ordered dolomite and microbial dolomite
然而,Gregget al.[7]对前人数据加以重新评估,发现微生物诱导沉淀形成的所谓“白云石”并不是有序的,其XRD 衍射图谱中白云石超结构衍射线并不完整或可靠(图8)。严格地讲,如果上述三个主要的超结构衍射线缺失两个及以上,就基本表明晶体里的Ca2+与Mg2+的排列并不完全有序,该矿物应该更接近方解石族而不是白云石族[9]。鉴于此,大部分实验模拟生成的微生物“白云石”本质上更可能为超高镁方解石(VHMC)或原白云石(proto-dolomite)[7]。事实上,大量的实验数据本身也表明,官能团对阳离子的吸附粘结趋势存在差异[30,44]。由于Mg2+的水合能力大于Ca2+,打破Mg2+水合壳层需要更多能量,因此,EPS 优先吸附Ca2+而不是Mg2+[41],这必然导致其矿物成分更富钙,更趋于沉淀(超)高镁方解石而不是白云石。但值得注意的是,由于原白云石和超高镁方解石被认为是白云石形成过程中的前驱相或中间产物[9,96,146],微生物活动可以在一定程度上缩短中间过程[147],进而在合适的环境因素下,促进白云石的形成[43,148]。
4 结论与展望
微生物诱导沉淀是低温条件下形成“白云石”的重要路径,是对“白云岩问题”的重大补充。依靠矿物的特殊形貌来判识白云石的成因时,应该谨慎,并结合其他证据加以佐证。此外,由于目前缺乏有力的手段予以成因上约束,加之成岩改造对原始的形貌和地化特征的覆盖,使得判识古老白云石是否为微生物诱导成因依然是困难的。在这样的背景下,微生物作用对于古老白云岩的贡献尚不能得到有效评估,使得该模式目前难以独立地解释地质历史时期厚度达数百米甚至上千米的规模性白云岩。虽然存在上述挑战,但由于微生物活动能促进超高镁方解石和原白云石等中间产物的生成,显然微生物活动有利于白云石的形成。当然,地质历史(尤其是前寒武时期)微生物的种类、数量和生理代谢方式可能远远超过我们目前的认知范围,微生物活动在白云石形成中的作用还需要进一步深入的工作。
致谢 感谢沈俊芳老师(洛阳理工学院)对理解“球晶生长”提供的帮助,感谢审稿专家的宝贵意见。
猜你喜欢
中国高新科技(2022年16期)2022-12-07
河北地质(2022年2期)2022-08-22
河南冶金(2021年1期)2021-04-27
黑龙江水利科技(2020年8期)2021-01-21
矿产勘查(2020年5期)2020-12-25
石油与天然气地质(2020年1期)2020-03-06
同位素(2018年1期)2018-01-18
安徽冶金科技职业学院学报(2015年3期)2015-12-02
同位素(2014年3期)2014-06-13
同位素(2014年2期)2014-04-16
