
长白山9种槭树的翅果扩散及种子萌发研究
2022-03-31 06:00:28张金峰葛树森李玉堂李俊清
生态学报 2022年4期
张金峰,葛树森,李玉堂,李俊清,*
1 北京林业大学森林资源生态系统过程北京市重点实验室, 北京 100083
2 吉林省林业调查规划院, 长春 130022
种子扩散是植物生活史的重要阶段,也是影响植物更新、群落组成和生态系统功能的重要生态学过程[1—2]。逃避假说(Janzen-Cannell假说)认为扩散有助于种子萌发和幼苗存活,因为扩散脱离了母体附近由于高密度而引起的高死亡率[3—4];种子扩散后或埋藏或贮存于适宜萌发和幼苗定居的环境,进而有助于植物种群占据新的生境,也可以影响植物的侵移、分布和物种形成等一系列进化过程[5—6]。
植物通常需要借助动物、风力、水流来扩散种子[5,7—8],其中,风力驱动是翅果类物种扩散的主要途径,而翅果的形态特征直接影响扩散能力和植物种群的分布范围[9]。例如,臭椿(Ailanthusaltissima)、白蜡树(Fraxinuschinensis)果实的沉降速度和水平扩散距离与翅果重量、种翅面积、翅载力显著相关[10—12]、原产于南美洲金虎尾科(Malpighiaceae)的许多物种依靠翅果扩散至非洲和亚洲热带地区等[13—14]。另外,自然情况下,风速对翅果的脱落及散布距离起重要作用[15—16],叙利亚马利筋(Asclepiassyriaca)种子在风速为2.2 m/s时才能脱落,且风速越大,越有利于增大其在水平方向上的扩散距离[15]。与之相似,强风条件下,绒毛白蜡(F.velutina)的种子能有效地脱离母株并扩散的更远[17]。然而,也有学者认为,风速增大可能会导致种子扩散速度增加,从而限制种子的远距离扩散[18]。
槭属(Acer)植物的许多物种是长白山针阔混交林的重要成分,在维持本地区森林群落稳定方面发挥着重要作用[19—20]。该属植物的果实为典型的双聚单翅果[21]。目前,许多研究工作都致力于理解翅果脱落和扩散的空气动力学[11—12,22—23],然而翅果的形态结构和传播特征在种间和种内均存在很大差异并对翅果的扩散能力有一定的影响[10,24]。此外,尽管一些研究对槭属植物种子的萌发机理进行过一些探讨,并认为种子的休眠特性是限制其萌发的重要因素[25],但槭属植物种子能否萌发不仅与自身的生物学特性有关,受扩散距离的影响也较大,因为对于适应风力扩散的物种而言,扩散距离的远近,往往能反映种子的优劣[1,26—27],种子被扩散的越远,重量可能越小,越不利于萌发[28]。
基于此,本研究以9种槭树的成熟果实为材料,对其翅果形态进行分析,在此基础上,通过模拟翅果从母株上垂直降落的速度及不同风速下的水平扩散距离,对不同槭树的扩散能力进行评估;并根据翅果的水平扩散距离,比较种子在不同扩散距离上萌发能力的差异,研究结果可为揭示翅果风力传播机制、槭属植物种子萌发对水平扩散距离的响应机制提供科学依据,对理解9个树种在长白山区的空间分布格局、种群更新和演替都有重要意义。
1 材料和方法
1.1 翅果采集
试验材料于2020年9月采自长白山国家级自然保护区(127°40′—128°16′ E,41°35′—42°25′ N)针阔混交林内,随机挑选白牛槭(A.mandshuricum)、茶条槭(A.ginnala)、簇毛槭(A.barbinerve)、花楷槭(A.ukurunduense)、假色槭(A.pseudosieboldianum)、拧筋槭(A.triflorum)、青楷槭(A.tegmentosum)、色木槭(A.mono)和小楷槭(A.komarovii)结实植株各10株,用高枝剪或树剪采集每树种的成熟果实数个并带回实验室,置于室内自然通风一周,按4分法各随机挑选无虫蛀、种翅完整的果实90枚进行标记编号。
1.2 方法
1.2.1翅果形态性状测量
用游标卡尺(测量精度0.01 mm)测量翅果长、翅果宽、种子长、种子宽、种子厚度;万分之一电子天平测量90枚完整果实重量、90枚去翅果实重量;利用扫描仪(Canon Scan Lide 120)和图像分析仪(Image J)测定种翅面积。根据测得的数据计算狭长度和翅载力。
狭长度=翅果长/翅果宽[1]
翅载力=翅果重量/种翅面积(g/cm2)[29]

图1 9种槭树的翅果形态
1.2.2翅果沉降速度测定
采用并改进Anderson[30]和郝建华等[8]的方法测定翅果在稳定空气中的垂直沉降速度(以下简称沉降速度)。在高200 cm、内径20 cm的塑料透明圆柱筒中释放翅果以确定其在稳定空气中的沉降速度。为消除初始下降时对翅果沉降速度的影响,在筒的上方盖一个中空的硬塑料板,并于中空处连接一个倒扣的塑料杯,将翅果从塑料杯底部的小孔投下。测定时用镊子轻轻夹住翅果,自然松开镊子,使其自然沉降(图2 A)。用电子秒表测量翅果从顶部到底部的降落时间,并换算成沉降速度。每个物种随机取90枚翅果,每个翅果释放3次,每次的沉降时间由两个人同时观测和记录,取其平均值。沉降速度v由公式换算得出:v=h/t,h为翅果释放高度,t为沉降时间。

图2 翅果扩散能力测定装置示意图
1.2.3翅果水平扩散距离测定
翅果在室内风力条件下的水平扩散距离的测定采用郝建华等[8]的方法并略加改进。翅果释放高度200 cm(模拟这些槭属植物部分果实释放的一般高度),由电风扇在种子释放处提供横定水平风,实验时风向固定。分布于长白山区的槭树果实一般于当年9月至10月间脱落,此时,该地区盛行2—3级西北风,偶有1级软风,风速约为0.3—5.4 m/s,据此,本试验设置风速1 m/s、2 m/s、4 m/s。为更好地观察翅果的扩散距离,测定时在风扇前方翅果降落区域的地面上摆放并固定白色滤纸。用镊子轻轻夹住翅果,在200 cm高度处自然释放,观察并记录翅果在其水平方向上的扩散距离(图2 B)。每个种随机取90枚翅果,每个翅果释放3次。
1.2.4种子萌发
根据上述测得的9种槭树水平扩散距离(仅探讨风速为4 m/s时扩散距离对种子萌发的影响),将每个种的90枚种子分为3组,每组30枚,分别视为较短距离、中等距离和较远距离扩散。每组的30枚种子分为3小组作为3个重复,每小组10枚种子。萌发试验在室内智能人工气候箱(RXZ- 280C型,宁波江南仪器厂)控制的25℃恒温条件下进行。槭树种子具有休眠性,试验前,将各小组的种子于清水中浸泡4 d并于500 mg/L的赤霉素溶液中浸种24 h[25]。随后将每小组的10枚种子播于直径为120 mm的培养皿中,以双层滤纸为萌发基质,用蒸馏水沿滤纸边缘注入培养皿底部,待浸透滤纸后将种子整齐排列于滤纸上。播种后将培养皿置于人工气候箱中开始试验(光照条件设置为14 h光照/10 h黑暗,光强约8800 lx)。每24 h观测记录种子萌发一次,以胚根突出种皮界定为种子萌发[31],观察记录至连续2周无种子萌发为止。试验期间每48 h更换1次滤纸并衡量补充蒸馏水。萌发率计算公式如下:

1.3 数据分析
采用Excel 2007和SPSS 13.0对数据进行统计分析,采用单因素方差分析法(one-way ANOVA)和最小显著差异法(LSD)及Person相关系数法分析比较不同数据组间的差异显著性和相关性。用SigmaPlot 12.0软件作图。图表中数据为平均值±标准差。
2 结果与分析
2.1 9种槭树翅果形态特征分析
9种槭树翅果的形态统计分析见表1。翅果重量排序为:拧筋槭>白牛槭>簇毛槭、色木槭、假色槭、青楷槭>小楷槭、茶条槭、花楷槭(注:符号前后差异显著,P<0.05,下同);翅果长排序为:拧筋槭>白牛槭>簇毛槭>青楷槭、茶条槭、小楷槭、色木槭>假色槭>花楷槭;9种槭树翅果宽在不同种间差异不显著;拧筋槭种子的长和宽均显著大于其它8种植物(P<0.05),花楷槭和茶条槭分别具有最小的种子长和宽;种子厚度排序为:白牛槭>拧筋槭>假色槭>簇毛槭>青楷槭>花楷槭、小楷槭>色木槭、茶条槭;拧筋槭种子重量最大(0.170 g)、白牛槭次之(0.078 g)、花楷槭最小(0.013 g);种翅面积排序为:拧筋槭>白牛槭>簇毛槭>青楷槭、色木槭、小楷槭>假色槭>茶条槭、花楷槭。以上结果表明,9种槭树翅果和种子的8个表型性状在种间差异较大,其中,拧筋槭、白牛槭各性状值均较大,花楷槭、小楷槭和茶条槭各性状值均较小。

表1 9种槭树翅果形态特征统计分析(平均值±标准差)
2.2 9种槭树翅果传播特性分析
2.2.1翅果狭长度和翅载力
翅果狭长度能够大致反映种翅的形状,白牛槭和茶条槭狭长度较大,二者也显著大于其它7种槭树(P<0.05);翅载力是风力传播结构常用的参数,其排序为:拧筋槭>白牛槭>假色槭>色木槭、簇毛槭、青楷槭>茶条槭、小楷槭、花楷槭(表2)。
2.2.2沉降速度
翅果沉降速度涵盖了两个数量级(表2),从沉降速度最快的拧筋槭(143.219 cm/s)到沉降速度最慢的花楷槭(74.960 cm/s)。对种子的沉降速度均值进行多重比较的结果显示,拧筋槭的沉降速度分别显著高于白牛槭、茶条槭、簇毛槭、花楷槭、假色槭、青楷槭、色木槭、小楷槭的24.709%、85.794%、61.773%、91.061%、25.670%、60.654%、27.185%、86.566%,其中,沉降速度在白牛槭、假色槭、色木槭之间无显著差异,青楷槭和簇毛槭以及茶条槭和小楷槭间也无显著差异,花楷槭的沉降速度最小。

表2 9种槭树翅果风扩散特性统计分析(平均值±标准差)
2.2.3水平扩散距离
由表2可以看出,沉降速度最慢的花楷槭在不同风速(1 m/s、2 m/s、4 m/s)下的水平扩散距离均最远(分别为167.378 cm、215.744 cm、275.000 cm),尤其风速为1m/s时,它与其它种的扩散距离之间差异显著(P<0.05);风速为1 m/s和2 m/s时,沉降速度最快的拧筋槭水平扩散距离最短(分别为39.878 cm和157.133 cm),风速为4 m/s时,色木槭的水平扩散距离最短(207.289 cm)。此外,风速为1 m/s时,白牛槭、簇毛槭、假色槭、青楷槭、小楷槭之间的扩散距离在彼此之间无显著差异;风速为2 m/s时,茶条槭、簇毛槭、假色槭、青楷槭、小楷槭之间的扩散距离在彼此之间也未达显著水平;风速为4 m/s时,尽管花楷槭的扩散距离最远,但它与小楷槭和青楷槭之间差异不显著,此风速下,扩散距离最短的色木槭与簇毛槭、拧筋槭、假色槭和白牛槭间也无显著差异。
2.3 翅果形态性状与扩散能力之间的相关性
由表3可以看出,翅果形态特征之间相互关联。例如,翅果重量与翅果长、种子长、种子宽、种子厚度、种子重量呈正相关性;种子长与翅果长、种子宽、种子厚度、种子重量、种翅面积呈正相关性等。翅果形态特征与翅载力之间多呈正相关性。翅果沉降速度除翅载力与其呈正相关外,与其它所有翅果形态特征呈负相关关系;翅果水平扩散距离与沉降速度呈负相关性。
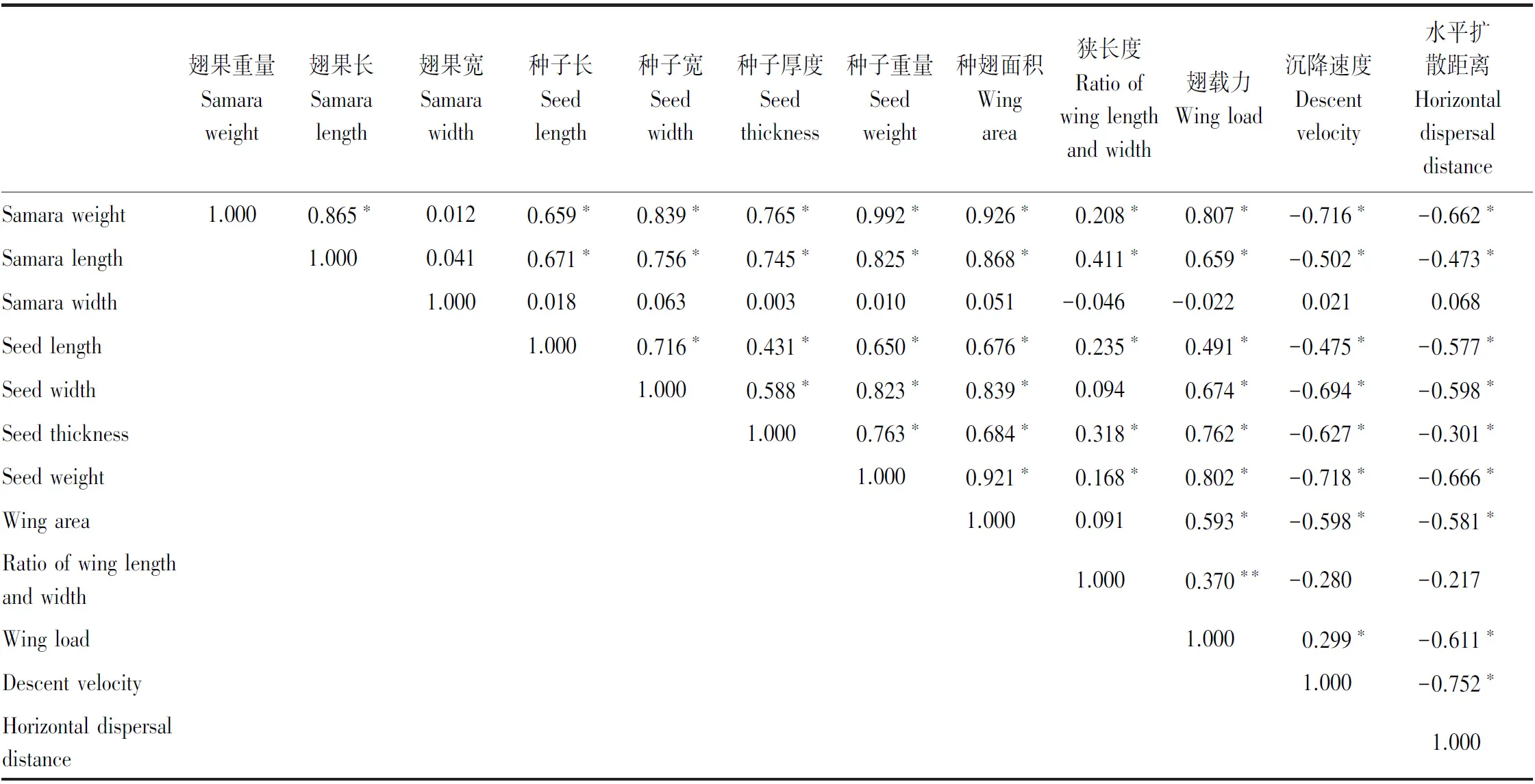
表3 翅果形态特征与风扩散特征的相关性分析
2.4 种子萌发与水平扩散距离间的关系
根据测得的9种槭树水平扩散距离,将种子分为3组,分别视为较短距离、中距离和较远距离扩散(图3)。可以看出,不同槭属植物种子萌发均不相同,且随扩散距离增加,所有树种种子萌发率均不同程度降低。茶条槭和青楷槭种子萌发率在不同扩散距离间差异显著(P<0.05);色木槭种子萌发率在较短和中等距离无显著差异,但二者分别显著高于较远距离的82.835%和66.845%;簇毛槭、假色槭和拧筋槭种子萌发率在中等距离显著低于较短距离的37.187%、40.000%、50.035%,在较远距离显著低于较短距离的42.523%、50.157%和62.197%,但中等距离和较远距离间无显著差异;尽管白牛槭、花楷槭和小楷槭种子的萌发率在不同扩散距离间无显著差异,但仍呈下降趋势。
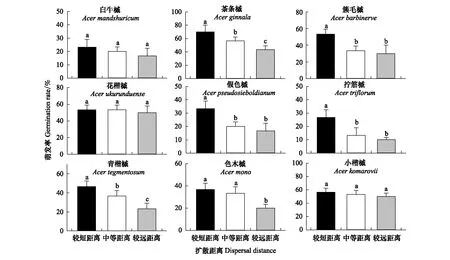
图3 9种槭树种子在不同扩散距离上的萌发率
3 讨论
3.1 翅果形态与扩散能力
风力扩散的繁殖体形态对物种的扩散能力影响显著。一般而言,重量很轻并有“飞行器官”的果实可以延长飞行时间,如种翅、冠毛[22,26],有学者甚至根据风播植物的形态特征进行物种扩散能力的分类[8]。然而,本文9种槭树的翅果重量、翅果长、翅果宽和翅果面积与沉降速度和水平扩散距离呈负相关性,种翅较长或较宽的拧筋槭和白牛槭也不具有最强的扩散能力,表明对于具有明显不同扩散能力的物种而言,翅果形态并不是最佳分类指标,这可能与翅果形态性状之间的协同变化有关[16]。例如,种翅面积较大的物种通常具有较高的种子重量,而种子重量往往是限制风播植物远距离扩散的重要因子[10—12]。上述现象也在本试验得到验证。例如,9种槭树的翅果长、宽和种翅面积与种子重量、种子长和宽及种子厚度均呈正相关,植物表型的协同变化是同种植物对环境长期适应的结果[32],槭树果实的翅果长、宽和种翅面积越大,其支撑的种子重量、种子厚度以及种子长和宽亦越大,翅果及种子性状间的“权衡”可能是槭属植物果实适应风力扩散的重要机制[1,24]。同时,本研究中,种子重量与沉降速度和水平扩散距离呈负相关,这是因为种子重量越大,其在空中停留的时间越短,下降速度越快,扩散距离也越近[16,29]。
翅果长度与宽度的比值,即狭长度能大致反映翅果的形状[1],狭长度越大,翅果越细长,反之越短粗,翅果形状的变化通常与空气动力学有关[22]。本研究中,狭长度与翅果扩散能力间无显著性关性,这可能是因为本试验所用的材料均为双聚单翅果,此类型翅果的沉降和远距离扩散依靠螺旋形向下旋转完成[10,22],众多研究表明,与翅果的长和宽相比,翅果的旋转更多地与旋转轴、种翅卷曲度、回转半径、力矩中心以及沉降过程中产生的前缘涡流等有关[33—34]。翅载力常用来表征物种的扩散能力[1],是指种翅单位面积上所承受的翅果重量(种翅重量和种子重量),本研究中,翅载力与沉降速度呈显著正相关,说明种翅单位面积上的负荷越大,种子沉降速度越快[24],这一结果与Peroni[21]对欧洲红枫(A.rubrum)的研究结论一致,对周位翅果、棱翅果和翼状萼翅果的研究也得出类似结论[35],然而,该研究结果与黑松(Pinusthunbergii)种子的“翅载力与沉降速度负相关”的结论相反[36],这可能与不同种翅后缘卷曲度引起的空气阻力有关,另一方面,尽管黑松种子具有较高的翅载力,但由于旋转速度快、锥角低,从而降低了其沉降速度[36]。
3.2 9种槭树的扩散能力比较
翅果的沉降速度是决定扩散距离远近的一个重要因素[17]。本研究中,对9种槭树翅果沉降速度的均值进行多重比较,从小到大可划分为5个子集:Ⅰ花楷槭;Ⅱ小楷槭和茶条槭;Ⅲ青楷槭和簇毛槭;Ⅳ假色槭、白牛槭、色木槭;Ⅴ拧筋槭,且各子集间差异显著(P<0.05)。一般而言,翅果的沉降速度越慢,其在空气中漂浮的时间越长,扩散距离越远,风力传播能力也越强[18]。然而,在自然环境中,翅果很少在静止的空气中漂浮,树冠上方的风速在确定翅果的散布距离中起着重要作用[15—16],且同属的不同种间由于具有不同的形态结构,扩散能力也不相同[8]。研究发现,翅果沉降速度和扩散距离间呈负相关性,且沉降速度最慢的花楷槭在不同风速(1 m/s、2 m/s、4 m/s)下的水平扩散距离均最远(分别为167.378 cm、215.744 cm、275.000 cm),尤其风速为1 m/s时,它与其它种的扩散距离之间均存在显著差异(P<0.05),因此,实现种子长距离扩散并扩大其分布范围的概率更高。此风速下,沉降速度最快的拧筋槭水平扩散距离仅为39.878 cm,究其原因可能与其具有较大的种子重量有关。风速增大时,所有槭树的水平扩散距离均不同程度地增大,其中,拧筋槭扩散距离变化最明显,表明强风有助于提高物种的扩散能力[15,24]。相关研究也表明,强风条件下,钻形紫菀(Astersubulatus)和小飞蓬(Conyzacanadensis)的翅果能有效地脱离母株,并被传播的更远[37]。
尽管翅果适合于风传播,但初始扩散距离仍较低。本试验中,翅果的释放高度为2 m,能代表野外环境中茶条槭、假色槭、簇毛槭、小楷槭果实释放的一般高度,然而四个物种扩散的最远距离却不足3 m,与此类似,高大乔木欧洲白榆(Ulmuslaevis)成熟果实的散布距离小于30 m[38],如此小的散布距离可能有助于区域内种群的维持,但并不能解释物种的远距离扩散。野外调查发现,假色槭和小楷槭孤立木近20 m内仍有扩散后的种子,这可能跟二次扩散有关,例如,被风散后,欧亚槭(A.pseudoplatanus)的种子仍吸引着啮齿类动物捕食[24,39]。二次扩散使果实被运送到较远或更加适宜建苗的地方[40],因此,有翅种子的二次传播机制可能是翅果类物种种群传播和拓植的一个重要因素。
3.3 扩散距离与种子萌发
通常来说,具翅物种中重量较轻的种子扩散更远。因此,扩散距离的远近,往往能体现种子的优劣[26—27]。本研究根据9种槭树的水平扩散距离,将种子分为3组,分别模拟较近距离、中等距离和较远距离,发现随着扩散距离的增加,种子萌发率逐渐降低,其中,茶条槭和青楷槭种子萌发率在不同距离间差异显著(P<0.05),Pearson相关分析也表明,种子重量与扩散距离呈显著负相关。上述结果表明扩散距离对种子萌发具有重要作用,即扩散距离越远,种子重量越小,越不利于种子萌发。Morse & Schmitt[15]和McAlpine & Jesson[26]分别以入侵草本植物叙利亚马利筋(A.syriaca)和灌木物种达尔文小檗(Berberisdarwinii)种子为材料的研究结论与上述结果一致,有关其他乔木树种的研究也有类似报道[1,24]。der Weduwen & Ruxton认为风播植物在初始扩散过程中的种子可能较轻,萌发率也较低,而那些能实现远距离扩散并在新环境中成功定植的物种,其种子质量通常较高,且受多次扩散的影响较大[24],例如,初始扩散后的挪威槭(A.platanoides)、梣叶槭(A.negundo)和臭椿(A.altissima)种子在食草动物或水流介导下的二次扩散可能会增加成功发芽的几率[11,41—42]。
本研究还发现,尽管9种槭树种子经清水及赤霉素溶液浸泡,但萌发率仍较低,甚至个别种的萌发率不足20%,且9种槭树的种子萌发率也均有所不同;另外,即使拧筋槭、白牛槭种子重量较大,但其萌发率较其它种的小,上述结果表明槭树种子的萌发能力在不同种间差异较大,究其原因可能与种子本身的休眠程度、种皮厚度等生物学特性有关[25]。此外,外界环境也是影响种子萌发的重要因素,野外调查发现,长白山区的花楷槭和小楷槭多分布于高海拔地段,其余槭树多分布于低海拔地段,海拔梯度上的温度差异可能对其萌发及种群分布具有重要影响,本试验是在智能人工气候箱控制的25℃恒温条件下进行,这一温度条件可能不利于部分槭树种子萌发,且植物种不同,种子萌发所需的最适温度也不同[25]。另据报道变温或贮藏(干贮藏和冷层积)是槭树种子萌发的必要条件[25]。因此,生物学特性、温度以及贮藏条件如何影响不同槭树种子萌发还需进一步研究。
4 结论
本研究以9种槭树的成熟果实为材料,通过模拟翅果从母株上垂直降落的速度及不同风速下的水平扩散距离,得到以下结论:
(1) 翅果形态性状间的“权衡”可是槭属植物果实适应风力扩散的重要机制,但翅果形态并不是最佳分类指标,而翅载力能较好地反映物种的风散播能力。
(2) 翅果在空气中停留的时间越长,其沉降速度越小,被风吹得越远,有助于植物物种广泛散布其后代,且强风有助于提高翅果的扩散能力。
(3) 沉降速度最慢的花楷槭在不同风速下的水平扩散距离均最远,其实现翅果远距离扩散和扩大分布范围的概率也更高;沉降速度最快的拧筋槭水平扩散距离最短;二次扩散可能是翅果类物种种群传播和拓植的重要机制。
(4) 水平方向上扩散距离体现了种子的优劣。即扩散距离越远,种子重量越低,越不利于种子萌发。
猜你喜欢
矿冶工程(2022年6期)2023-01-12 02:14:56
——曾来德小楷展
中华书画家(2022年6期)2022-11-04 11:21:45
草堂(2022年7期)2022-10-22 02:00:20
海洋通报(2020年4期)2020-12-11 01:56:44
中国油脂(2020年9期)2020-09-16 03:15:14
中国油脂(2019年7期)2019-08-22 09:40:32
中国水土保持科学(2019年6期)2019-04-26 05:14:24
丹青少年(2018年5期)2018-10-24 01:34:42
书画艺术(2016年1期)2016-04-21 22:02:34
生态毒理学报(2014年5期)2014-09-21 01:59:33
