
选择性雌激素受体调节剂抑制变异链球菌的作用探究
2022-03-31 09:45廖盛楠吕炜桐唐权马玗玟刘力嘉王亮彭显
华西口腔医学杂志 2022年2期
廖盛楠吕炜桐唐权马玗玟刘力嘉王亮彭显
1.口腔疾病研究国家重点实验室 国家口腔疾病临床医学研究中心四川大学华西口腔医院正畸科,成都61004;
2.口腔疾病研究国家重点实验室 国家口腔疾病临床医学研究中心四川大学华西口腔医院,成都610041
变异链球菌(Streptococcusmutans,S.mutans)是引起龋齿的主要病原菌,在人的牙齿表面生长并分泌葡萄糖基转移酶、葡聚糖结合蛋白等,以形成牙菌斑[1-2]。长期以来,氟、氯己定和其他抗菌药物是龋齿治疗的常规方法[3-4]。然而,变异链球菌对这些抗菌药物产生的耐药性已经成为全球公共卫生的挑战[5-6]。开发一种新药需要大量的劳动力、资金和时间成本,药物再利用是指在临床上已经广泛使用的药物基础上开发新的治疗效果,是临床上开发新药的的替代选择。近年来,小分子药物防龋研究引起广泛关注。通过对大量小分子化合物、天然产物等化合物库的筛选,再通过靶点分析,寻求更能结合其靶蛋白活性位点的小分子,最终找到具有选择性、特异性抑制致龋菌及牙菌斑生物膜形成的小分子药物,达到开发新型抗龋药物的目的。
选择性雌激素受体调节剂(selective estrogen receptor modulators,SERMs)是一类美国食品药物管理局(Food and Drug Administration,FDA)批准的作用于雌激素受体的药物。与纯雌激素受体激动剂和拮抗剂不同,其因在不同组织中的作用不同而具有选择性地抑制或刺激雌激素样作用。临床常用的SERMs包括他莫昔芬、托瑞米芬、克罗米芬和雷洛昔芬。他莫昔芬是雌激素受体阳性转移性乳腺癌的一线激素治疗药物;托瑞米芬是一种氯化他莫昔芬衍生物,与他莫昔芬相比,在肝脏中产生的DNA加合物更少,并且被开发用于避免肝癌;克罗米芬是一种用于治疗未排卵妇女(包括多囊卵巢综合征妇女)不育症的药物;雷洛昔芬用于预防和治疗高危妇女中的绝经后骨质疏松和乳腺癌[7-9]。SERMs还具有抗菌、抗病毒和神经保护作用,已经有少数研究[8,10-12]显示了SERMs对革兰阴性细菌具有有效的抗菌性能。但目前仍然缺乏对SERMs药物系统的筛选,并且较少研究它们对口腔致龋菌,如变异链球菌的抗菌作用,且具体的作用机制需要进一步的探索。
转座子插入为细菌基因组的随机突变提供了一种有效方法,已被广泛用于细菌发病机制和生物学研究中。课题组前期基于Himar1的转座子的自杀载体pMagellan6,开发了变异链球菌突变体库。Himar1是mariner家族的转座子,利用Himar1转座酶的点突变体增加了该酶的转座率,使其能被开发成为基因操作的工具。Himar1转座子转座的唯一序列要求是TA二核苷酸序列。通过对转座子插入的确切位置的鉴定即可找到突变基因位点。一种对Himar1转座子非常有效的方法是在转座子的反向重复序列中引入限制性内切酶MmeⅠ识别位点。用它消化基因组时,MmeⅠ会从识别位点侧向消化20 bp序列,产生一个包含转座子和反向重复序列的片段。通过特定的Illumina兼容序列连接到转座子两侧的突出端碱基上,然后进行聚合酶链反应(polymerase chain reaction,PCR)和Illumina深度测序,即可精确定位插入位点。
本研究旨在建立基于Himar1转座子的变异链球菌突变体库,可以通过大量突变体筛选探索抗菌药物的作用位点。
1 材料和方法
1.1 细菌复苏和培养
取-80℃冻存的变异链球菌UA159标准菌株,室温下解冻后接种于脑心浸出液(brain heart infusion agar,BHI)琼脂培养基中,37℃恒温厌氧箱(80%N2,10%CO2,10%H2)内复苏24 h,挑取单克隆菌株继续培养24 h后制备细菌悬液,用PBS将细菌浓度调整为1×106CFU·mL-1,4℃备用。携带质粒的大肠杆菌菌株在含有红霉素(12μg·mL-1)的Luria-Bertani(LB;Difco)培养基中37℃培养。
1.2 药物的准备
实验中所有使用的FDA批准的小分子化合物都购买于成都宝科生物科技有限公司。其中SERMs包括托瑞米芬、他莫昔芬、克罗米芬和雷洛昔芬。将药物制备了一系列浓度的稀释液:将所有药物分别溶于DMSO中,稀释至50、25、12.5、6.25、3.125、1.56μmol·L-1的浓度。
1.3 最低抑菌浓度(minimum inhibitory concentration,MIC)值的测定
使用微量稀释法测定小分子化合物对变异链球菌UA159的MIC值。将稀释后不同浓度的小分子药物溶液分别加到灭菌的96孔板中,第1至第11孔加药液,每孔10μL,第12孔加入等量无菌PBS作为阴性对照。将单克隆的UA159菌落在BHI液体培养基中复苏一夜,用BHI将菌液稀释20倍后再生长至OD470为0.6,并每孔加入2μL到96孔细胞培养板中。然后在5%CO2下厌氧培养16 h,并通过酶标仪记录每个孔在OD600的光密度(optical density,OD)值。以在孔内完全抑制细菌生长的最低药物浓度为MIC,当对照孔(即PBS组)内细菌明显生长实验才有意义。
1.4 变异链球菌突变体库的构建
利用mini-Himar1转座子构建变异链球菌的突变体库[13]。从异丙基硫代半乳糖苷(isopropyl thiogalactoside,IPTG)诱导的大肠杆菌中分离出含有MarC9转座酶的粗提取物,然后通过亲和色谱法使用淀粉树脂柱纯化该酶。MarC9转座酶纯化后,用8%的十二烷基硫酸钠聚丙烯酰胺凝胶电泳(sodium dodecyl sulfate polyacrylamide gel electrophoresis,SDS-PAGE)分析测定最终产物的纯度。使用1μg细菌DNA和1μg Magellan6在2 mL培养基中进行体外转座。在转座酶MarC9介导下,含有壮观霉素和红霉素抗性的Magellan6通过剪贴机制被引入线性的细菌基因组DNA片段中。转座后,每个转座子和线性基因组DNA的连接处存在单链的间隙,用T4DNA聚合酶和大肠杆菌DNA连接酶进行修复。
随后,包含Magellan6插入片段的线性基因组DNA被转化入感受态细胞中。即在BHI中过夜培养的变异链球菌的菌株被1∶10/1∶20/1∶30稀释,并在37℃中厌氧培养至OD600值达到0.2~0.3(约2~3 h)。培养物在室温下静置10 min。在1.5 mL离心管中,将1μg的用于转化的DNA加入500~1 000μL细胞培养液中,并以30℃厌氧额外培养2~3 h。在培养之后稀释细胞,将一部分涂板于含有抗生素的BHI选择培养基上,在37℃下培养,直到可见菌落。
1.5 插入位点的鉴定
从选择培养基上挑选出成功转座的突变体菌株后,识别转座子的插入位点。转座子左右两端的反向重复序列里设计有MmeⅠ识别序列,用ⅡS型限制酶MmeⅠ对转座子插入的突变体的基因组DNA进行消化。MmeⅠ在其识别位点的下游切割了20个bp的序列。产生的一部分DNA片段包含了转座子的反向重复序列和20 bp的侧翼DNA。预先制备一种特定的Illumina兼容序列,并存储在
-20℃。
用DNA连接酶在转座子两侧的突出端碱基连接上Illumina兼容序列,并在16℃培养一夜,即产生120 bp的插入位点片段。应用PFU超缓冲液、dNTPs、引物P1_M6_MmeI、Gex PCR引物2和PFU超聚合酶等进行PCR以扩增序列。通过Illumina深度测序,即可精确定位插入位点。
2 结果
2.1 抑制变异链球菌的小分子化合物的筛选
作为FDA批准的药物,这些药物的生物安全性已经得到了测试和保证。用微量稀释法测定了426个FDA批准的小分子化合物对变异链球菌UA159的MIC。研究发现,这些小分子化合物中有18种小分子抑制剂对变异链球菌具有抗菌效果,分别属于以下类别(表1)。除了传统抗生素外,还发现了以往认为与抗菌活性无关的药物。在确定的具有抗菌特性的18种小分子化合物中,有4种属于选择性雌激素受体调节剂,结构存在差异(图1),展现出良好的抗菌能力。
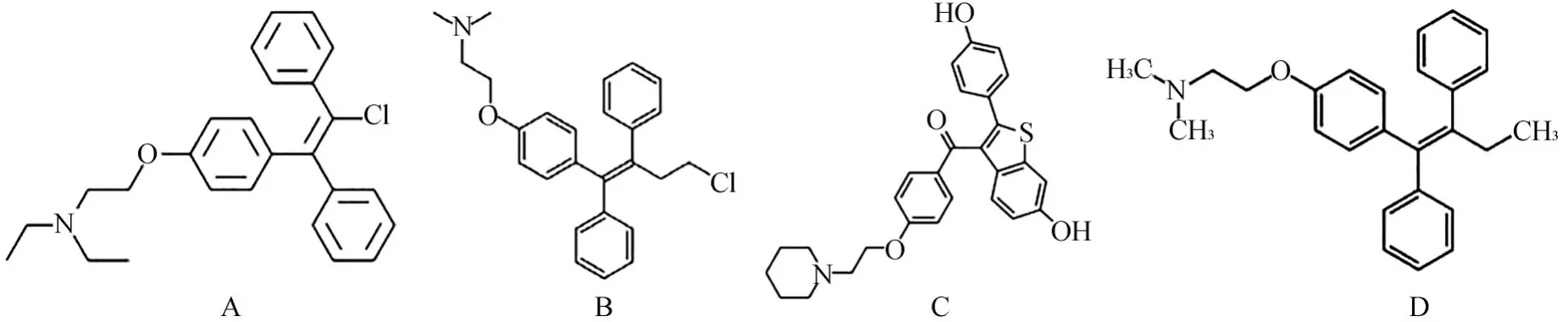
图1 筛选出的4种SERMs的化学结构Fig 1 Chemical structuresof four selected SERMs

表1 筛选出的具有抑制变异链球菌活性的小分子化合物Tab 1 Selected small molecular inhibitors which have anti-S.mutans activities
2.2 变异链球菌突变体库的成功构建
为了研究SERMs的抗菌机制,以克罗米芬为例进行探索。利用Himar1 Mariner转座子系统构建了变异链球菌的突变体库。其中包括一系列的实验:MarC9酶的纯化、Magellan6的转化和突变株的筛选。为了在体外转化Magellan6,成功地构建了质粒PVPT作为载体。从IPTG诱导的大肠杆菌中分离并纯化MarC9转座酶后,用于之后的转座子系统中。在诱导细菌进入感受态后,含有转座子的DNA被转化入变异链球菌中。采用含有壮观霉素的培养基验证变异链球菌是否转化成功。Magellan6上携带有壮观霉素抗性,因此没有成功转化DNA的细菌将被抑制生长。对于成功转化的细菌,在37℃的选择性BHI平板上可以看到菌落,得到了成功转化的突变体(图2A)。为了确认转座子是否成功插入菌株,在提取细菌DNA后,通过PCR扩增了120 bp的插入位点片段。10个随机挑选的单个菌落均出现了120 bp序列片段对应的条带(图2B)。

图2 变异链球菌突变体的成功转化Fig 2 Thesuccessful transformation of S.mutans
通过对这120 bp的片段的测序可以确定转座子插入的位置。因此,对片段进行了测序,并在变异链球菌的基因组进行了比对,证实了突变体库中不同位点的随机插入。在以上10个被测序的菌落中,9个呈阳性插入,插入位点的识别如下(表2)。

表2 变异链球菌突变体的插入位点的鉴定Tab 2 Identification of the insertion sites of S.mutans
通过菌落计数统计,从4个独立的转化实验中收集了大约45 000个转座子插入突变体。而变异链球菌UA159共有约2 160个基因,数目庞大的随机突变足以覆盖其全部非必须基因,此突变体库可以作为研究药物作用靶点的有效工具。
2.3 SERMs抗菌机制的探索
由于DNA片段的体外转座和随后质粒的转化都是低频事件,绝大多数的突变菌株都只有单个转座子插入。为了筛选出具有克罗米芬抗性的突变体,将突变体库的不同菌株添加到含克罗米芬的BHI琼脂平板上。在37℃下培养48~72 h后,最终获得了2个抗克罗米芬的突变菌株。在没有克罗米芬的条件下培养至少30代后,2个突变体的菌株仍然可以表现出稳定的抗克罗米芬的表型,结果表明这2种突变体的耐药性与转座子插入密切相关。对DNA序列的测序分析结果表明,转座子插入位点位于smu_546、smu_874的基因中(图3)。
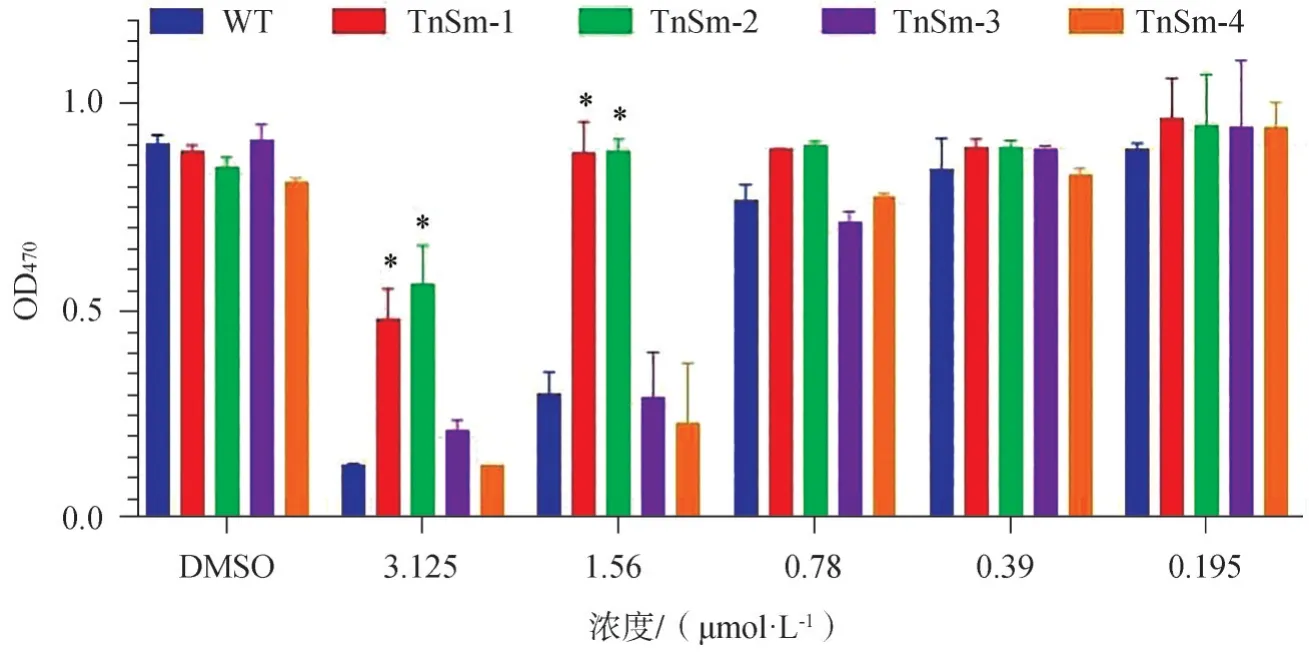
图3 不同的变异链球菌突变株在克罗米芬中的生长情况。其中TnSm-1(smu-546基因突变)和TnSm-2(smu-874基因突变)两个菌株具有克罗米芬抗性Fig 3 The growth condition of different mutant strains in clomiphene.Two mutant strains,TnSm-1(Smu-546 gene mutation)and TnSm-2(Smu-874 genemutation),werefound to beresistant to clomiphene
3 讨论
龋病是一种慢性细菌感染性疾病,氟化物、氯己定和其他抗菌的药物长期使用导致了细菌耐药性的出现,促使去探寻更有效地对抗变异链球菌的药物,以实现龋病的防治。从FDA已经批准的药物中开发新的抑菌功能是一个很好的方向。本实验从FDA批准的药物中发现了SERMs这类药物具有良好的抑菌效果,并系统地探究其抗菌作用。尽管近年来SERMs的抑菌能力受到了一定关注,但对口腔致龋菌——变异链球菌的抑制作用探究较少。本实验通过建立变异链球菌的突变体库,进一步探究SERMs的作用机制。将SERMs和临床中最常见、最有效的口服抗菌剂相比,SERMs比临床中使用的氯己定具有更低的抑菌浓度[14]。氯己定通过改变细菌生物膜的渗透性来抑制细菌的生长,且具有一定的不良反应。由于抑菌机制的不同,SERMs可能解决变异链球菌产生的耐药性的问题。此外,与氯己定一样,SERMs在其表面具有正电荷,这有助于药物与牙菌斑结合,并稳定地释放药物[15]。
在之前的研究中,克罗米芬通过抑制十一异戊烯二磷酸合酶(Undecaprenyl-diphosphate synthase,UPPS)来抑制金黄色葡萄球菌,该合成酶负责合成的十一异戊烯二磷酸是脂质载体Und-P的直接前体,从而降低了用于合成磷壁酸(wall teichoic acid,WTA)的Und-P,对细胞壁产生作用[16]。克罗米芬与β-内酰胺在抑制金黄色葡萄球菌方面具有协同作用,这可能归因于UPPS的抑制降低了脂质Ⅱ,而脂质Ⅱ被用作PBPs的转糖基作用和转肽作用的底物。然而,克罗米芬对不同的细菌种类可能具有不同的抗菌机制。变异链球菌的突变体库提供了2个对克罗米芬耐药的菌株。对这两个菌株的DNA序列分析表明,转座子插入位点位于smu_546、smu_874的基因中。smu_546编码一种推定的三磷酸鸟苷(guanosine triphosphate,GTP)结合蛋白,它是一种参与应激反应的膜GTP酶。smu_874编码一种双功能的同型半胱氨酸S甲基转移酶/5,10-亚甲基四氢叶酸还原酶蛋白。然而,需要进一步的实验来验证是否是这2个基因的突变导致了变异链球菌对克罗米芬的耐药性的产生。其他SERMs药物,例如托瑞米芬和他莫西芬,通过将细菌的细胞膜去极化和渗透作用,诱导离子流出,导致跨膜电位的丧失,严重破坏膜的形成和导致细菌细胞壁的膨胀,也作用于脂质和蛋白质,导致细胞裂解[17-20]。作为同一类药物,克罗米芬与它们具有相似的化学结构和性质,可能具有相似的机制影响变异链球菌。
柠檬酸克罗米芬已被用于临床治疗,其安全性问题不容忽视。许多临床试验和Meta分析系统地研究和介绍了与它的安全性有关的问题。临床上,使用大剂量的克罗米芬治疗与多胎妊娠、先天性血管瘤和卵巢癌有关[21-22]。在本研究中,克罗米芬的MIC仅为3.125μmol·L-1,治疗口腔感染所需的剂量远远小于安全范围[23-25]。在细菌生物膜的影响下,不同药物在体内的实际抗菌效果可能不同。大量的研究[26]表明,由于生物膜的保护作用,变异链球菌在生物膜中对抗菌剂的敏感度远低于体外的浮游状态,因此,应进行进一步的体内实验来研究SERMs在体内的抗菌作用及其防龋效果。口腔的生态环境是一个由多种细菌相互作用的复杂环境,SERMs药物是否只特定地针对变异链球菌发挥抗菌作用,或者对其他口腔细菌也产生相应的影响,进而调控口腔微生态的平衡,也需要进一步的研究。
不同的SERMs抑菌作用明显不同,可能是由于其化学结构的不同。克罗米芬、他莫昔芬和托瑞米芬对变异链球菌有良好的抗菌活性。它们都是三苯乙烯类衍生物。虽然目前尚不能确定三苯乙烯的结构是否与其抗菌活性有关,但据报道[27],三苯乙烯结构的变化会显著影响其抗感染活性。也有研究[28]记录了三苯乙烯通过干扰钙的稳态达到了抗真菌作用。需要进一步的实验确定这些药物的抗菌机制与此结构有关,也可能通过修饰三苯基乙烯类药物上的基团来合成更适合抑制细菌生长的药物。
利益冲突声明:作者声明本文无利益冲突。
猜你喜欢
中国感染与化疗杂志(2022年3期)2022-12-12
亚热带农业研究(2022年1期)2022-08-08
江苏卫生保健(2022年5期)2022-05-24
家庭医学(2021年11期)2021-12-29
皮肤病与性病(2021年3期)2021-07-30
农业科技通讯(2021年1期)2021-03-06
趣味(数学)(2020年4期)2020-07-27
支部建设(2020年15期)2020-07-08
中国农业科技导报(2020年3期)2020-03-15
山西农业科学(2020年2期)2020-02-29
