
慢性冷暴露对小鼠肝脏抗氧化功能的影响
2022-03-30 05:21刘媛媛薛念宇张博熙李士泽吕红明
动物营养学报 2022年3期
刘媛媛 薛念宇 张博熙 李士泽 吕红明
(黑龙江八一农垦大学动物科技学院,大庆163319)
1 材料与方法
1.1 试验材料
1.1.1 试验设计
体内慢性冷暴露模型建立:将12只3周龄C57BL/6雄性小鼠(购自中国人民解放军军事医学科学院试验动物中心),体重(22±1) g,随机分为2组(对照组和冷暴露组),每组6只。先将小鼠置于人工智能气候室中进行为期7 d的适应性饲养,环境温度设定为(28.0±0.5) ℃,相对湿度为(40±5)%,自由采食、饮水。日光灯照明,明暗周期比为1∶1(开灯08:00,关灯20:00)。然后将冷暴露组小鼠每日随机置于4 ℃环境中3 h,连续4周,冷暴露结束后将小鼠同时安乐死,并收集其血液和肝脏。将血液样本静置在4 ℃冰箱中12 h,然后3 500 r/min离心20 min,取上清液至1.5 mL离心管中保存。将肝脏放进1.5 mL离心管中,做好标记保存在-80 ℃冰箱。试验涉及的试验动物操作符合黑龙江八一农垦大学动物伦理委员会标准(20180328002)。
体外冷暴露模型建立:将小鼠肝细胞系AML12细胞(购自美国ATCC公司)随机分为4组(对照组、冷暴露1 h组、冷暴露3 h组、冷暴露6 h组),先在37 ℃培养箱中培养24 h,然后将冷暴露组细胞置于32 ℃培养箱中分别亚低温冷刺激0、12、24、36 h,冷刺激结束后将细胞刮取下来进行后续试验。
1.1.2 主要试剂
Trx-1(1∶1 000,A0053)抗体购自ABclonal公司(中国);TXNIP(1∶1 000,A0053)抗体购自ABclonal公司(中国);辣根过氧化物酶(HRP)山羊抗鼠免疫球蛋白G(IgG)(1∶10 000,#SA00001-1,Proteintech,美国);HRP山羊抗兔IgG(1∶10 000,#SA00001-2,Proteintech,美国)。DMEM培养基、胰蛋白酶由赛默飞世尔生物化学制品(美国)有限公司生产;Solarbio血清由北京Solarbio & Technology公司生产;磷酸盐缓冲液(PBS)由北京中杉金桥生物技术有限公司生产。酶联免疫吸附测定(enzyme linked immunosorbent assay,ELISA)试剂盒由南京建成生物工程研究所生产。
1.2 试验方法
1.2.1 细胞培养与传代
每天对小鼠肝细胞系进行换液,2 d进行1次传代。将DMEM高糖培养基,胎牛血清(FBS)和青链双抗分别按照90%、10%和1%的比例配制成细胞完全培养基。首先将冻存的细胞进行复苏,操作步骤:1)将完全培养基、PBS和胰酶提前在37 ℃水浴锅中预热30 min,同时将酒精灯、PE手套、15 mL离心管、50 mL离心管、移液枪、枪头等放进无菌操作台进行紫外灭菌30 min;2)将冻存的小鼠肝细胞系从液氮罐中取出,迅速放入37 ℃水浴锅中融化,约1 min后取出,放入无菌操作台中;3)从冻存管中吸出培养基放进15 mL离心管中,并缓慢加入1 mL完全培养基,用移液枪来回吹打混匀;4)然后1 000 r/min离心4 min,离心结束后吸弃上层培养基;5)加入2 mL PBS,1 000 r/min离心4 min,离心结束后吸弃上层PBS;6)再加入1 mL完全培养基,吹打混匀后转移到含有4 mL完全培养基的T25培养瓶中,“8”字混匀后转移到37 ℃恒温培养箱中培养。
待细胞状态稳定后进行传代培养,操作步骤:1)待细胞长满70%~80%后,先吸弃细胞瓶中的培养基,加入PBS 2 mL,对细胞进行清洗,之后再吸弃PBS;2)加入1 mL胰酶,放入37 ℃恒温培养箱中约1 min,待贴壁细胞完全消化后,拿到无菌操作台中,加入2 mL完全培养基进行终止反应,并轻轻吹打细胞瓶底部,以保证细胞完全脱离;3)将细胞瓶中含有的完全培养基转移到15 mL离心管中,1 000 r/min离心4 min,离心结束后吸弃上层培养基;4)然后加入2 mL PBS,轻轻地将细胞吹散,1 000 r/min离心4 min,离心结束后吸弃上层PBS;5)加入1 mL完全培养基,吹打混匀后转移到含有4 mL完全培养基的T25培养瓶中,“8”字混匀后转移到37 ℃恒温培养箱中培养,待细胞系完全稳定后开始试验。
本文提出了对于接入海上风电场的区域电网的多风电场无功优化算法,在MATLAB中验证所提出算法的有效性和正确性。
1.2.2 细胞计数
先将肝细胞用胰酶消化下来,然后往消化下来的细胞中加入完全培养基制成细胞悬液,充分混合后,吸取10 μL细胞悬液与10 μL台盼蓝染色液混合,混合均匀后吸取10 μL滴加在血细胞计数板上计数并记录。然后将细胞平均铺到8个35 mm细胞培养皿中,待细胞在培养皿中培养24 h后,将细胞放入32 ℃培养箱中冷刺激12、24、36 h,冷刺激结束后将细胞消化下来,再次进行计数并记录。
1.2.3 血清生化指标
小鼠血清中MDA和GSH含量分别按照碧云天的MDA和GSH检测试剂盒的说明书进行操作。
1.2.4 Western Blot检测抗氧化相关蛋白表达
先用NP40裂解液(碧云天,中国)分别提取细胞和肝脏组织的总蛋白,用BCA蛋白浓度测定试剂盒(碧云天,中国)进行蛋白浓度的测定,然后把蛋白浓度调成一致。首先配制10%或者12%的分离胶、5%浓缩胶,按每孔20 μg蛋白量上样进行十二烷基硫酸钠聚丙烯酰胺凝胶电泳(sodium dodecyl sulfate polyacrylamide gel electrophoresis,SDS-PAGE)。其次根据“三明治”结构进行转膜。转膜结束后用5%脱脂奶粉-三羟甲基氨基甲烷(Tris)盐溶液和吐温-20(TBST)溶液室温封闭1 h,然后一抗4 ℃过夜。第2天先用TBST洗涤5次,每次5 min,然后按1∶10 000稀释HRP标记山羊抗兔IgG和HRP标记山羊抗鼠IgG配制成二抗,室温孵育1 h。最后再用TBST洗涤5次,每次5 min。滴加显影剂后,在化学放光检测器中显影,用Image Pro Plus 5.0软件进行图像分析。
1.3 数据的统计分析
所有统计分析均通过统计软件Graphpad Prism 5.0完成。对照组和冷暴露组数据采用单因素方差分析(one-way ANOVA),P<0.05为差异显著。
2 结果与分析
2.1 慢性冷暴露对小鼠血清生化指标的影响
如图1所示,与对照组相比,冷暴露组小鼠血清中的GSH含量被显著抑制(P<0.05),而MDA含量显著增加(P<0.05)。

与对照组相比,NS表示差异不显著(P>0.05),*表示差异显著(P<0.05)。下图同。
2.2 体外冷刺激对AML12细胞活力的影响
如表1所示,与对照组相比,随着冷刺激时间的延长,活细胞数量逐渐下降(P<0.05),并表现出一定的时间依赖性,说明冷刺激可能对细胞生长增殖产生一定的抑制作用。

表1 体外冷刺激对AML12细胞活力的影响
2.3 不同冷暴露时间对小鼠肝细胞系AML12细胞抗氧化蛋白表达的影响
如图2所示,与对照组相比,冷暴露3和6 h后,抗氧化蛋白Trx-1的表达量显著减少(P<0.05)。而冷暴露6 h后,TXNIP蛋白的表达量显著增加(P<0.05)。结果说明冷暴露会导致小鼠肝细胞AML12的抗氧化功能受到抑制。

TXNIP:硫氧还蛋白互作蛋白 thioredoxin interacting protein;Trx-1:抗氧化蛋白硫氧还蛋白-1 thioredoxin-1;β-actin:β-肌动蛋白。
2.4 慢性冷暴露对小鼠肝脏组织中抗氧化蛋白表达的影响
如图3所示,与对照组相比,冷暴露小鼠的肝脏组织中抗氧化蛋白Trx-1的蛋白表达量显著降低(P<0.05),TXNIP蛋白的表达量显著增加(P<0.05),与体外试验结果一致,进一步说明了冷刺激会抑制抗氧化蛋白的表达,降低抗氧化功能。
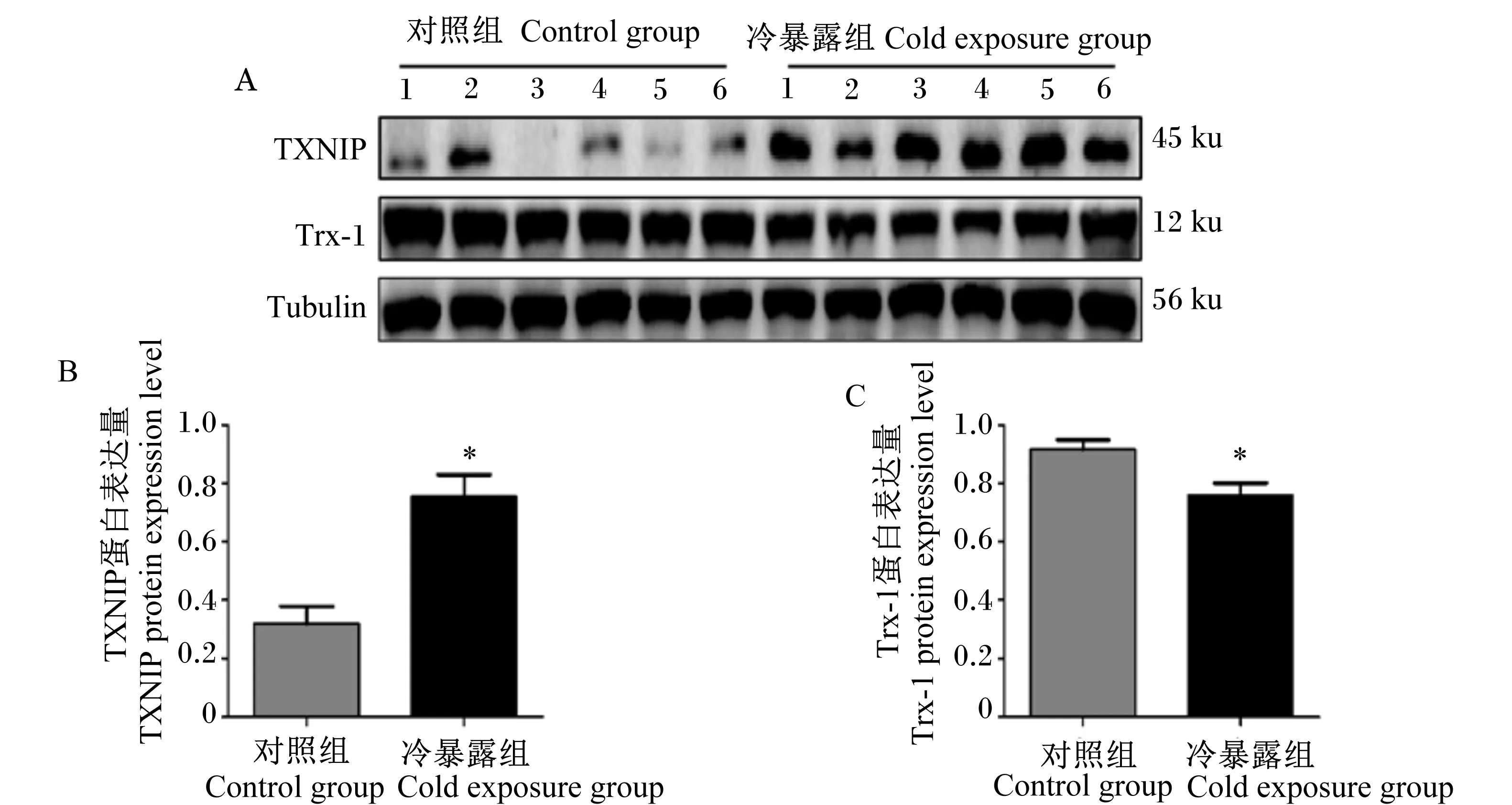
TXNIP:硫氧还蛋白互作蛋白 thioredoxin interacting protein;Trx-1:抗氧化蛋白硫氧还蛋白-1 thioredoxin-1;Tubulin:微管蛋白。
3 讨 论
3.1 慢性冷暴露对小鼠血清生化指标的影响
王金涛[5]的研究表明,冷应激会造成机体的抗氧化功能受到不同程度的影响。在生理状态下,细胞内氧化动态平衡受到活性氧(ROS)的产生和细胞内防御机制,以及抗氧化酶的严格调控。比如GSH是哺乳动物细胞中含量最丰富的非蛋白硫醇,它所在的GSH系统能够与Trx-1交叉提供电子,并作为彼此的备份系统,一起控制哺乳动物细胞中的氧化还原环境[6],其活性的高低间接反映了机体清除氧自由基的能力;MDA是脂质发生过氧化反应后产生的氧化终产物,具有细胞毒性,其含量是反映机体抗氧化潜力的重要指标。本试验结果表明,4 ℃慢性冷暴露后会导致小鼠血清中GSH含量显著减少,MDA含量显著增加,说明机体在暴露于寒冷环境中会使抗氧化酶的活性降低,氧化产物增多,从而导致抗氧化能力下降。
3.2 慢性冷暴露对小鼠肝细胞活性的影响
李宝贵等[7]的研究表明,大鼠在低温刺激后,肝细胞间隙变小,肝脏血窦结构突出明显,肝细胞会发生变形,并且随着低温刺激时间的延长,这些变化更加明显。苏莹莹[8]通过苏木精-伊红(H&E)染色观察到,大鼠暴露于低温后,肝脏细胞会发生水肿,有丝分裂增加并且出现大量的双核细胞和少量的凋亡细胞;电镜结果发现肝细胞糖原减少,线粒体发生轻微肿胀。本试验结果表明,小鼠肝细胞AML12用32 ℃亚低温处理后,细胞数量显著减少,并呈现出一定的时间依赖性减少趋势,说明冷暴露会对细胞造成损伤,产生一定的负面影响。
3.3 慢性冷暴露对小鼠肝细胞及肝脏组织中抗氧化蛋白表达的影响
肝脏作为体内重要的代谢器官,也是体内最大的糖原储存器官和静息状态时主要的产热器官,在受到冷应激后,会产生不同程度的损伤[9]。此外,当动物暴露于低温环境中,会导致肝糖原被大量消耗,分解为葡萄糖为机体提供能量底物,但此时肝脏通过偶联作用产热,这样会使肝脏出现氧化应激损伤[10]。同时,机体其他组织和器官的大量能量代谢中间产物可以通过血液运输到肝脏,在肝脏内进行生物转化,形成新的能量底物或被肝脏代谢,同样导致肝脏代谢率增加和ROS的产量增加[11]。在体内,Trx系统由烟酰胺腺嘌呤二核苷酸磷酸(NADPH)、TrxR和TXNIP组成,通过其二硫键还原酶活性调节蛋白质二硫醇/二硫键平衡,从而成为抵御氧化应激的关键抗氧化系统,是氧化蛋白的还原剂[12]。Trx-1的抗氧化功能还表现在通过减少核苷酸还原酶、蛋氨酸亚砜还原酶和调节许多氧化还原敏感转录因子的活性来参与DNA和蛋白质修复[13-14]。TXNIP是Trx-1的负性调节因子,可以阻碍其保护作用[15]。因此,Trx-1通过与TXNIP相互作用蛋白在免疫反应、病毒感染和细胞死亡中发挥重要作用[16-17]。本试验结果发现慢性冷暴露会导致小鼠肝脏和AML12细胞中抗氧化蛋白Trx-1的表达量显著减少,TXNIP的蛋白表达量显著增加,进一步说明了慢性冷暴露会导致小鼠肝脏的抗氧化能力下降。
目前,畜禽场主要通过以下几个方面防控冷应激,比如加强动物房舍系统的防风和保暖功能;通过改善饲粮的配方加强畜禽的御寒能力;或者繁殖出抗寒的品种等[18-20]。本研究涉及相关指标的检测与整合将有助于拓展冷应激对小鼠肝脏抗氧化功能的认识,对提升畜牧抗寒能力、保证畜牧业持续稳定发展具有重要意义。
4 结 论
① 32 ℃亚低温处理后,AML12细胞数量显著减少,抗氧化蛋白Trx-1的表达量显著降低,说明冷刺激一段时间后会对小鼠肝细胞造成一定的氧化损伤。
② 4 ℃慢性冷暴露4周后,小鼠肝脏中抗氧化蛋白Trx-1的表达量显著降低以及血清中GSH和MDA的含量显著增加,进一步说明了冷暴露会导致小鼠肝脏抗氧化功能受损。
猜你喜欢
中国典型病例大全(2022年13期)2022-05-10
昆明医科大学学报(2022年2期)2022-03-29
昆明医科大学学报(2022年1期)2022-02-28
现代仪器与医疗(2021年6期)2022-01-18
临床与实验病理学杂志(2021年7期)2021-09-06
昆明医科大学学报(2021年3期)2021-07-22
科技视界(2020年26期)2020-09-24
科技视界(2020年17期)2020-07-30
数码世界(2020年4期)2020-06-18
食品工程(2020年3期)2020-01-05
