
一株金黄色葡萄球菌噬菌体 SAK84P 的分离鉴定
2022-03-25 09:18黎晏辰刘自强黄志鹏马迎飞尤晓颜
集成技术 2022年2期
黎晏辰 刘自强 赵 敏 谭 新 黄志鹏 马迎飞* 尤晓颜
1(河南科技大学 河南省食品微生物工程技术研究中心 洛阳 471000)
2(中国科学院深圳先进技术研究院 深圳合成生物学创新研究院 中国科学院定量工程生物学重点实验室广东省合成基因组学重点实验室 深圳市合成基因组学重点实验室 深圳 518055)
1 引 言
金黄色葡萄球菌(Staphylococcus aureus,S. aureus)是一类革兰氏阳性致病菌,属于微葡萄球菌科(Micrococcaceae)葡萄球菌属(Staphylococcus)[1]。其主要存在于哺乳动物的前颈部和皮肤表面[2],是引起群体感染的主要病原菌之一,可引起轻微的皮肤感染,甚至导致严重的感染性休克,也是菌血症和感染性心内膜炎以及骨关节、皮肤和软组织、胸膜肺、脊髓硬膜外脓肿等的主要诱因[3-4]。此外,金黄色葡萄球菌易在异物装置表面形成难以根除的生物膜,从而引发器械感染[5]。20 世纪 40 年代中期,Kirby 发现部分金黄色葡萄球菌菌株产生了质粒编码的青霉素酶,该酶可水解青霉素的 β-内酰胺环,形成青霉素抗性,导致群体中由耐青霉素的金黄色葡萄球菌引起的感染比例增加[6]。1961年,Barber[7]发现了耐甲氧西林的金黄色葡萄球菌(Methicillin-resistantStaphylococcus aureus,MRSA)菌株。但是,目前耐甲氧西林金黄色葡萄球菌已对许多抗生素都产生了耐药性。金黄色葡萄球菌的临床感染不仅增加了微生物的耐药性,而且临床疾病的范围也在不断变化,对耐药菌感染的预防和治疗都带来了挑战[8]。
噬菌体是地球上数量最多、种类最多的生物实体[9],是一种可特异性地裂解细菌的病毒[10],1917 年,Félix d’Hérelle[11]将一种可裂解细菌的物质命名为噬菌体。噬菌体的生命周期主要有两种,一种是溶源性生命周期:噬菌体将其基因组整合到宿主基因组上,可在宿主细胞中稳定存在若干代,而非直接杀死宿主细胞[12];另一种是裂解性生命周期:噬菌体感染细菌后迅速杀死宿主细胞[13]。对于细菌感染中常存在的生物膜,由于其被多糖包裹,抗生素不易穿透,采用特异性强、裂解能力强的噬菌体更容易杀死细菌,且噬菌体对宿主生物及其他有益菌无害,副作用小[14],因此被视为治疗细菌感染的潜在药物。噬菌体用于治疗金黄色葡萄球菌的感染已有相关报道。2018 年,杨航等[15]发现来自噬菌体裂解酶 PlyV12 的细胞壁结合结构域(CBD)的重组蛋白 V12CBD 可通过多种方式减弱金黄色葡萄球菌的毒力,并增强宿主的免疫防御能力。2019 年,Lehman 等[16]将 3 种肌尾噬菌体制成噬菌体制剂AB-SA01,经实验证明,该制剂在体外和小鼠模型中都能显著减少耐药金黄色葡萄球菌数目。Fabijan 等[17]给金黄色葡萄球菌感染的患者静脉注射噬菌体鸡尾酒制剂 AB-SA01,均无不良反应,证实了噬菌体治疗的安全性。噬菌体的独特属性给应用噬菌体治疗超级耐药菌感染提供了广阔的前景。
本研究以金黄色葡萄球菌 K84 为宿主,从环境污水中分离得到噬菌体 SAK84P,对其进行生物学特性及基因组学分析。裂解谱实验显示其可侵染耐甲氧西林金黄色葡萄球菌,具有临床价值,为探索噬菌体治疗耐药菌感染提供了理论基础,为应对超级耐药菌提供了新的抗菌药物和治疗方案。
2 材料与方法
2.1 菌株、样品来源和主要试剂和仪器
本研究所使用的菌株为金黄色葡萄球菌K84,SA 为实验室保存菌株(ATCC29213),SA1~SA11 为深圳市人民医院分离金黄色葡萄球菌;水样采自深圳市西丽农贸市场。其中,K84、SA、SA1~SA5 为甲氧西林敏感金黄色葡萄球菌(Methicillin-susceptibleStaphylococcus aureus,MSSA);SA6~SA11 为 MRSA。
LB 液体培养基:胰蛋白胨 10.0 g,酵母提取物 5.0 g,NaCl 5.0 g,1 L dd H2O,121 ℃,灭菌 30 min。LB 固体培养基:按上述液体培养基的配方,加入 1.2% 技术琼脂粉,作为下层琼脂平板;按上述液体培养基的配方,加入 0.7% 技术琼脂粉,作为上层琼脂平板。SM 缓冲液:NaCl 5.8 g,MgSO4•7H2O 2 g,50 mL 1 mol/L Tris-HCl,1 L dd H2O,121 ℃,灭菌 30 min。
0.22 μm 滤膜产自 Millipore 公司,2×Ex Taq酶产自 TaKaRa 公司,2000 DNA Marker 产自生工(上海)股份有限公司,落地离心机为 Thermo ST40R,超速离心机为 Himac CP 80NX,电泳仪为 BIO-RAD PowerPac,透射电镜为 JEM-F200。
2.2 噬菌体的分离纯化
噬菌体的分离纯化采用双层平板法[18]。将采集的污水与金黄色葡萄球菌 K84 共同培养 12 h 后8 000 rpm 离心 10 min,取上清液,作为第一次富集液,连续 3 次富集,将富集后的溶液离心收集上清液,过滤除菌,采用双层平板法进行噬菌体的分离纯化。将金黄色葡萄球菌 K84 与 0.7% 的LB 固体培养基混匀铺板,取 10 mL 滤液点板,放入 37 ℃ 培养箱,倒置培养,待有噬菌斑长出后,取单个噬菌斑浸泡于 SM 缓冲液中,加入K84 菌液与 0.7% LB 固体培养基混匀涂板,进行3 次纯化,直至噬菌斑形态单一。
2.3 噬菌体的裂解谱测定
本文选取 SA、临床分离甲氧西林敏感金黄色葡萄球菌菌株(SA1~SA5)和耐甲氧西林金黄色葡萄球菌菌株(SA6~SA11),采用双层平板法进行噬菌体宿主范围测定。将 K84、SA和 SA1~SA11 等金黄色葡萄球菌培养至对数期(OD600=0.5)后,与 LB 固体培养基混匀铺板,然后将 SAK84P 噬菌体稀释不同的梯度进行点板,观察有无噬菌斑出现。
2.4 噬菌体温度敏感性测定
取 100 μL(起始量为 108PFU/mL)的噬菌体液加入 EP 管中,分别放入 37 ℃、50 ℃、60 ℃、70 ℃ 的水浴锅中孵育,在 20 min、40 min、60 min各时间点进行取样,测试噬菌体滴度。同时设置3 个平行实验。
2.5 噬菌体 pH 耐受度测定
取 10 μL 稀释过的噬菌体液,按 1∶9 的体积比分别加入 pH 为 1.0、3.0、5.0、7.0、9.0、11.0、13.0 的 SM 缓冲液中,然后放入 37 ℃ 的水浴锅中孵育 1 h,进行噬菌体滴度测试。每个实验同时设置 3 个平行实验。
2.6 噬菌体抑菌曲线测定
取对数期(OD600=0.5)的金黄色葡萄球菌加入 LB 培养基中,按感染复数(Multiplicity of Infection,MOI) 分别为 0.001、0.01、0.1、1、10、100 加入噬菌体,用 96 孔板放置 37 ℃ 酶标仪中连续震荡培养,间隔 10 min 测量 OD600的吸光值。用 LB 培养基作为空白对照,阳性对照为只加入金黄色葡萄球菌的 LB 培养基,每个实验设置 3 个平行实验。
2.7 噬菌体一步生长曲线测定
本实验采用的一步生长曲线的测定方法是基于 Pajunen 等[19]的方法,并对该方法略有改动。取100 μL 对数期(OD600=0.5)的金黄色葡萄球菌(约107PFU/mL),按 MOI=0.01 加入稀释后的噬菌体(约 105PFU/mL),混匀后取样作为第一次测量值,然后放入 37 ℃ 摇床中,转速为 220 rpm,每10 min 进行一次取样,共测量 80 min,采取双层平板测定各时间点的噬菌体滴度,设置 3 个平行实验,根据测试结果绘制出一步生长曲线。
2.8 噬菌体形态鉴定
噬菌体的形态可通过透射电子显微镜进行观察[20]。先将富集后的噬菌体用 CsCl 梯度密度离心,取纯化后的噬菌体滴于铜网上,吸附 10 min,用滤纸小心擦去多余液体,再在铜网上滴加 2%磷钨酸,染色 2 min,静置于阴凉干燥处,24 h后用透射电子显微镜观察。
2.9 噬菌体全基因组测序
将富集后的噬菌体基因组用天根基因组提取试剂盒进行提取,文库构建采用 Illumina Nextera DNA Flex Library Prep,全基因组测序采用Illumina 平台。序列组装采用 SOAPdenovo[21],开放阅读框预测采用 GeenMark[22],使用 Blastp工具进行序列相似性分析,序列注释主要使用 NCBI 数据库——NR(Non-Redundant Protein Sequence Database),tRNA 预测使用 tRNAscan-SE 工具[23],使用 CGView 在线软件对全基因组图谱进行绘制[24]。本文选择较为保守的主要衣壳蛋白、末端大亚基、DNA 聚合酶 3 种蛋白,用Clustal W 进行蛋白序列比对[25],并用 MEGA X软件[26]进行系统进化树的构建。
3 结果分析
3.1 噬菌体裂解谱
对于金黄色葡萄球菌临床分离菌株,SAK84P 呈现不同的裂解能力(表 1)。由表 1 可知,SAK84P 可侵染甲氧西林敏感金黄色葡萄球菌,也可侵染耐甲氧西林金黄色葡萄球菌,具有临床价值。
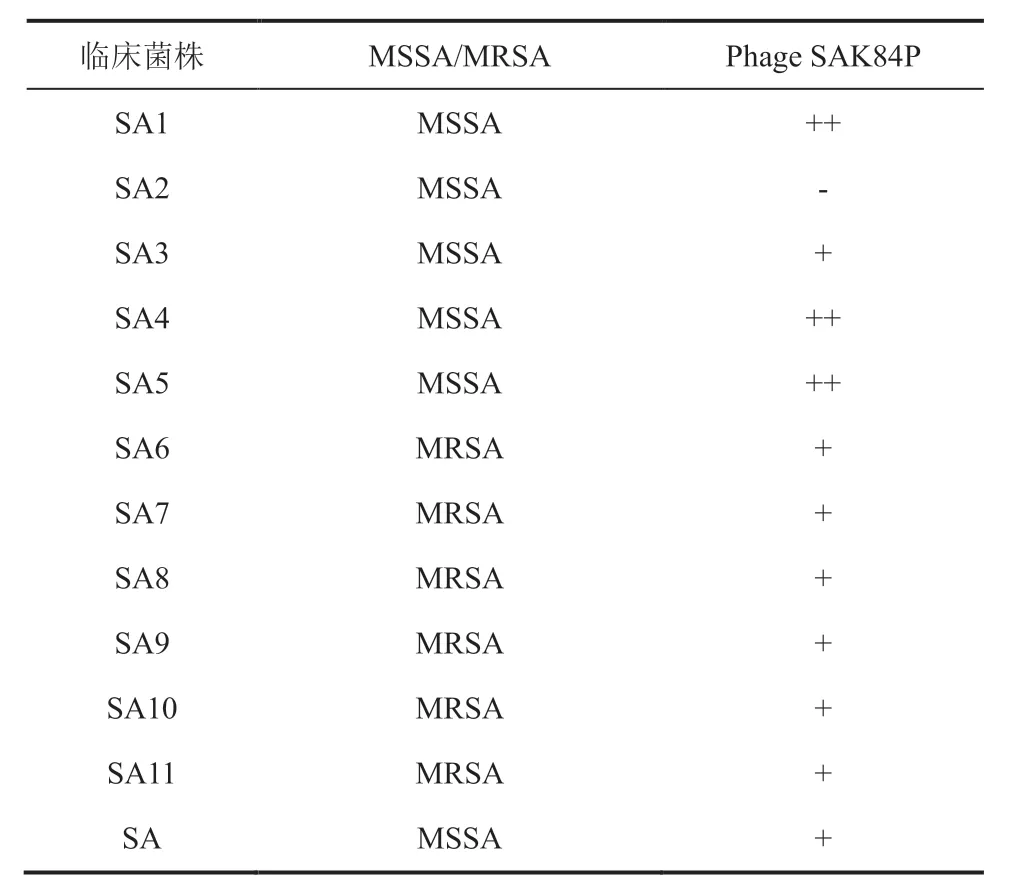
表1 噬菌体 SAK84P 的宿主谱Table 1 Host range of phage SAK84P
3.2 噬菌体温度稳定性
实验通过在不同温度的不同时间点对噬菌体的滴度进行测定,测定结果如图 1 所示。由图 1可知,金黄色葡萄球菌噬菌体 SAK84P 在 37 ℃时能保持很好的活性,具有繁殖能力;当温度为50 ℃ 时,噬菌体仍能存活且保持 108PFU/mL 滴度;但当温度高于 50 ℃时,噬菌体迅速失活。
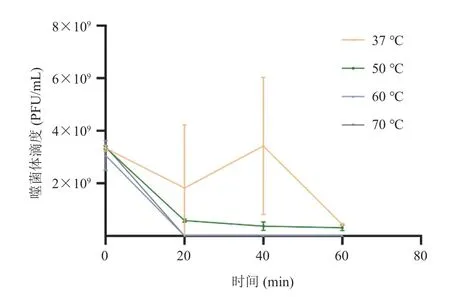
图1 噬菌体 SAK84P 的热稳定性Fig. 1 Temperature stability of phage SAK84P
3.3 酸碱耐受度
将噬菌体稀释液分别加入 pH 为 1.0、3.0、5.0、7.0、9.0、11.0、13.0 的 SM 缓冲液中,在 37 ℃ 的水浴锅中孵育 1 h 后对噬菌体进行滴度测定,测定结果如图 2 所示。由图 2 可知,SAK84P 在 pH 3.0~11.0 环境下具有生物学活性,其中,pH 为 5.0~7.0 是最适合噬菌体生长的环境;当 pH<5.0 或 pH>7.0 时,噬菌体活性急剧下降;在 pH<3.0 或 pH>11.0 时,噬菌体几乎无裂解能力。
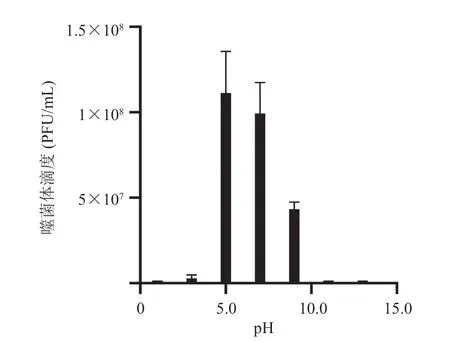
图2 噬菌体 SAK84P 的酸碱稳定性Fig. 2 pH stability of phage SAK84P
3.4 抑菌曲线
将噬菌体与宿主菌以不同的 MOI 比例混合后共同培养,噬菌体 SAK84P 的抑菌曲线如图 3所示。当 MOI 为 0.001 时,加入 SAK84P 与只加入宿主菌 K84 的生长趋势几乎一致,表明若按此MOI 加入噬菌体,则无法抑制宿主菌的生长;当MOI 为 0.01 和 0.1 时,对金黄色葡萄球菌 K84 的裂解能力较强,在 OD600大于 1 时,即菌液较浓时仍可以显著抑制宿主菌的生长;当 MOI 为 1、10、100 时,可对宿主菌产生一段时间的抑制,但在 360 min 后,抗性菌出现。因此,当 MOI 为0.01 和 0.1 时,可有效起到杀菌作用,并抑制细菌不再生长,所以其为最佳感染复数。
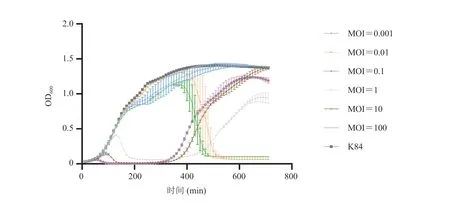
图3 噬菌体 SAK84P 抑菌曲线Fig. 3 Killing curve of phage SAK84P
3.5 一步生长曲线
对 SAK84P 进行一步生长曲线测定。在各时间点测得噬菌体计数除以起始点计数,即为平均每个噬菌体产生的子代数目(Burst Size)[27],用Burst Size 绘制出噬菌体 SAK84P 的一步生长曲线图如图 4 所示。由图 4 可知,SAK84P 的潜伏期为 20 min,裂解时间为 40~70 min,Burst Size 为100 PFU/cell。该噬菌体裂解能力较强,平台期较短,在 70 min 后噬菌体便进入下一阶段的裂解。

图4 噬菌体 SAK84P 一步生长曲线Fig. 4 One-step growth curve of phage SAK84P
3.6 噬菌体形态学鉴定
本实验以金黄色葡萄球菌 K84 为宿主菌,从污水中分离纯化得到一株烈性噬菌体 SAK84P,双层平板显示该噬菌体呈现透明的噬菌斑,直径约为 1~1.5 mm,边界清晰(图 5(a))。
由透射电镜结果(图 5(b))可观察到,噬菌体 SAK84P 的头部为多边形(近似六边形),其直径为 112 nm,尾巴长 200 nm,宽为 29 nm,且尾巴粗壮。根据国际病毒分类委员会的新国际病毒分类方法,该噬菌体属于有尾噬菌体目(Caudovirales),螺旋噬菌体科(Herelleviridae)[28]。

图5 噬菌体 SAK84P 的形态Fig. 5 Morphology of phage SAK84P
3.7 噬菌体全基因组测序
SAK84P 全基因组大小为 141 535 bp,GC含量为 30%,Blastn 结果显示与 Staphylococcus phage VB_ScoM-PSC1 (GenBank: MZ573923.1)最为相似,其中,98% 的覆盖度中有 99.96% 的序列一致性。SAK84P 全基因组的预测结果为,共有 224 个开放阅读框(Open Recording Frame,ORF)和 4 个 tRNA,且含有多个末端重复序列编码的蛋白。
3.7.1 基因功能预测
使用 GeenMark 对噬菌体基因组 ORF 进行预测,使用 NCBI 数据库——NR 进行 ORF 注释,注释结果用 CGView 软件进行噬菌体全基因组图谱绘制(图 6)。其中,62 个 ORF 在反义链上,162 个 ORF 为正义链编码的蛋白质。将预测结果中已知蛋白功能的 ORF 分为:DNA 复制和代谢相关的蛋白、结构相关的蛋白、裂解相关的蛋白、DNA 包装相关的蛋白,以及未知功能的蛋白。
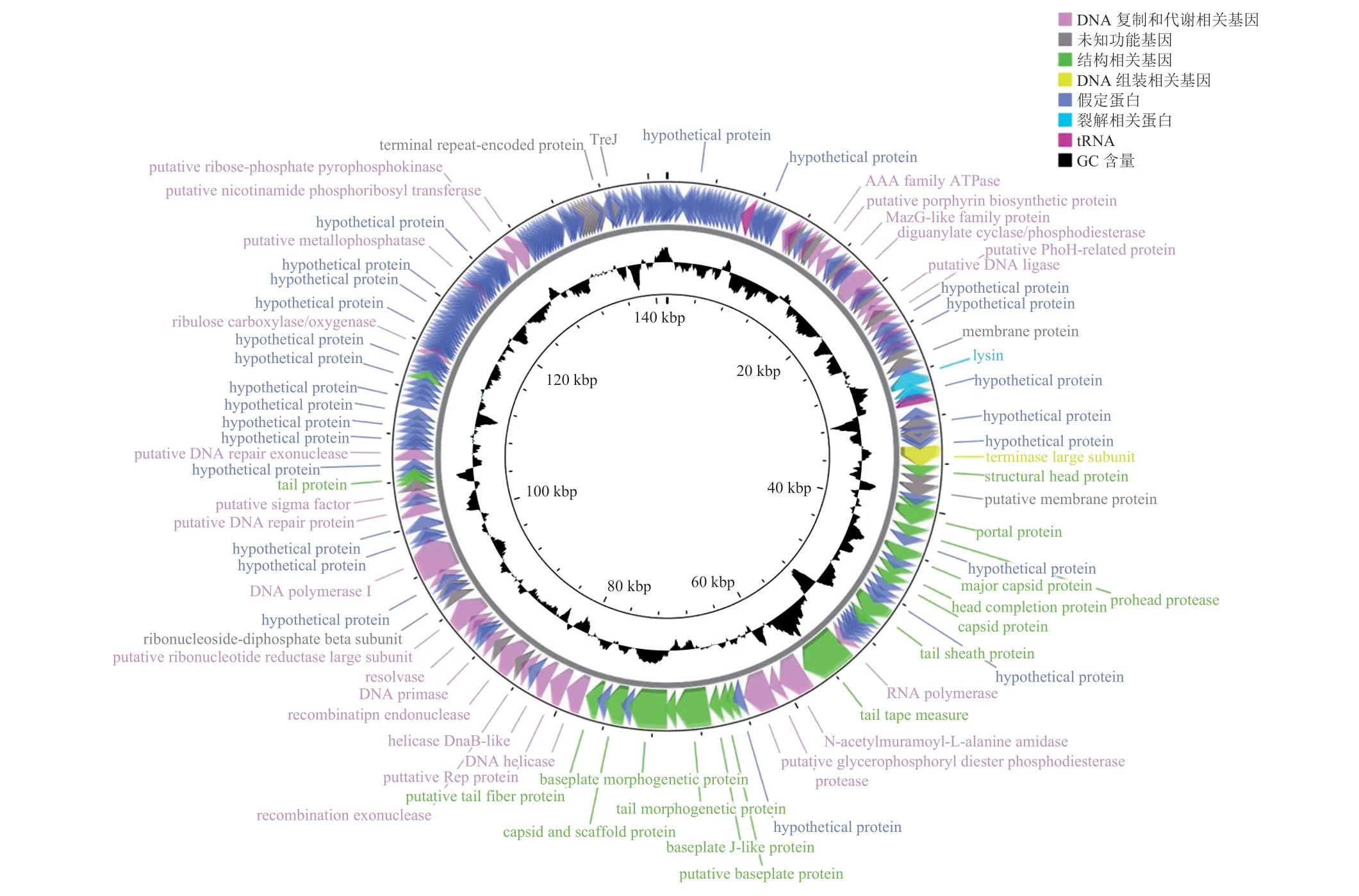
图6 噬菌体 SAK84P 全基因组图谱Fig. 6 Genome map of phage SAK84P
在该噬菌体基因组中,与 DNA 复制和代谢相关的基因有 ORF46(DNA 连接酶)、ORF92(RNA聚合酶)、ORF127(DNA 聚合酶 I)、ORF112(重组外切酶)、ORF139(DNA 修复外切酶)[29];与结构相关的蛋白包括 ORF78(主要衣壳蛋白)、ORF85(尾鞘蛋白)、ORF86(尾管蛋白)、ORF99(基板蛋白)、ORF135(尾丝蛋白)等,负责噬菌体形态相关的蛋白组装;裂解相关的蛋白主要有ORF58(裂解酶),其作为噬菌体感染宿主后的晚期基因表达的水解酶,通过破坏细菌细胞壁肽聚糖,帮助子代噬菌体的释放[30];DNA 包装相关的蛋白主要有 ORF69(末端大亚基蛋白),其负责将病毒 DNA 包装到空衣壳中[31],该蛋白多为保守蛋白,可利用该蛋白构建系统进化树。
3.7.2 噬菌体 SAK84P 系统进化树构建
针对噬菌体上的主要衣壳蛋白、末端大亚基、DNA 聚合酶等保守蛋白,构建系统进化树,进一步确定噬菌体 SAK84P 的分类,噬菌体 SAK84P 系统发育树如图 7 所示。由图 7 可知,SAK84P 噬菌体与Kayvirus属的噬菌体聚在一个分支上,且 SAK84P 与金黄色葡萄球菌噬菌体Staphylococcusphage G15 在这 3 种保守蛋白的氨基酸序列上较一致,与Staphylococcusphage LSA2308、Staphylococcusphage philPLAC1C 相距较远。与 SAK84P 噬菌体基因组核酸序列最为相似的是Staphylococcusphage VB_SavM_JYL01,其末端大亚基和主要衣壳蛋白都与 SAK84P 较相似,但 DNA 聚合酶存在差异。因此,SAK84P 的分类和Staphylococcusphage G15 一致,属于有尾噬菌体目(Caudovirales),螺旋噬菌体科(Herelleviridae),Twortvirinae亚科,Kayvirus属。该螺旋噬菌体家族由国际病毒分类委员会命名,具有肌尾噬菌体的形态[28]。
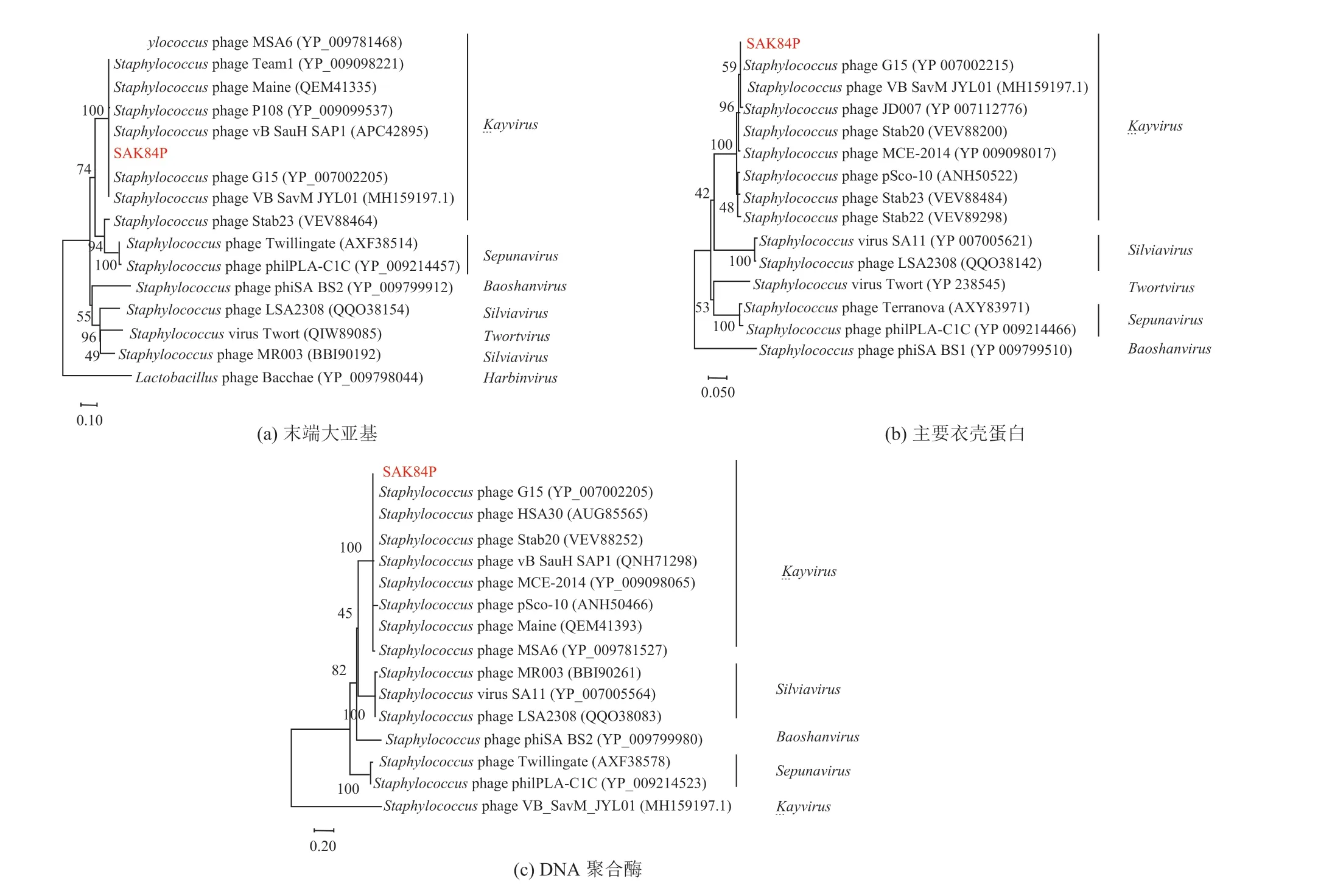
图7 噬菌体 SAK84P 系统发育树Fig. 7 Phylogeny tree of phage SAK84P
4 讨论与结论
抗菌素耐药性是一个全球关注的复杂问题,其可能会对人民健康和社会经济造成巨大影响。而金黄色葡萄球菌作为最棘手的细菌病原体之一,不仅对青霉素、甲氧西林不敏感,而且对其他抗生素(大环内酯等)和万古霉素也出现了耐药性。
噬菌体疗法在应对金黄色葡萄球菌感染中扮演了重要角色。Dickey 等[32]发现噬菌体辅助治疗可增强低浓度抗生素对金黄色葡萄球菌生物膜的杀菌作用;在金黄色葡萄球菌足部感染的小鼠模型中,Albac 等[33]使用 3 种噬菌体联合利奈唑胺,在小鼠后爪单次注射中,噬菌体均显示出良好的抗菌功效;Doub 等[34]使用 PM448 噬菌体成功治疗了顽固性葡萄球菌表皮假肢膝关节感染;Chhibber 等[35]用脂质体包裹噬菌体,治疗金黄色葡萄球菌引起的糖尿病伤口感染小鼠模型,可显著提高感染的治愈率、加快伤口愈合,且在包裹脂质体的伤口部位上,检测到噬菌体的存在更加持久。这些发现进一步支持噬菌体疗法的临床前和临床研究。由于噬菌体对宿主的高度专一性,所以应用噬菌体治疗耐药菌感染,不会引起菌群紊乱。噬菌体疗法应用已有 100 多年历史[36],临床上也未发现有明显的毒副作用[17]。
常规的噬菌体疗法依赖于天然的噬菌体对感染部位的菌株进行裂解,这需要针对不同耐药菌株分离相应的噬菌体,因此,治疗效果受噬菌体实体库容量的影响。而生物学特性是噬菌体能否作为生物制剂的先决条件,也是噬菌体未来应用于临床或生产的理论基础。本研究发现,噬菌体 SAK84P 能侵染多株临床分离的耐甲氧西林金黄色葡萄球菌,且该噬菌体裂解能力极强。当用 K84 作为宿主时,扩大培养后噬菌体滴度能达到 1010PFU/mL,可作为治疗 MRSA 的备用噬菌体。SAK84P 噬菌体在 50 ℃ 以下可稳定存在,37 ℃ 时生物学活性强,可适用于治疗人体耐药菌感染。此外,该噬菌体对 pH 有较大程度的耐受范围,在 pH 为 3.0~11.0 内都有裂解能力,在pH 为 5.0~7.0 时活性最高,说明该噬菌体对酸碱的耐受度广,可治疗细胞外感染和胞内感染,有较强的应用价值。本研究通过对 SAK84P 噬菌体进行抑菌曲线和一步生长曲线测定,得到最佳感染复数(MOI)为 0.01 或 0.1,可作为噬菌体鸡尾酒疗法中的添加比例,能更有效地抑制菌株生长[37];该噬菌体潜伏期为 20 min,Burst Size 为100 PFU/cell,高于陈冈等[38]报道的金黄色葡萄球菌噬菌体 SP-2(80),也显著高于金黄色葡萄球菌噬菌体 qdsa001(32.2)[39]。该噬菌体具有较短的裂解周期,平台期较短,在 70 min 完成第一阶段裂解后迅速进入下一个裂解周期,噬菌体的生长周期在临床应用中有重要意义。因此,该噬菌体在侵染宿主菌过程中可迅速产生大量子代,具有作为噬菌体生物制剂的潜力,也可用作消杀材料[38],后续将继续分离 MRSA 噬菌体,扩大金黄色葡萄球菌噬菌体实体库,通过噬菌体鸡尾酒疗法治疗更多临床中的耐药细菌感染。
利用噬菌体进行治疗的另一途径是利用生物技术等扩大治疗制剂范围,包括使用工程化噬菌体或纯化噬菌体裂解蛋白等策略[40]。Gu 等[41]使用纯化的葡萄球菌噬菌体 GH15 的裂解酶在体外杀死多株金黄色葡萄球菌,包括 MRSA,该酶还可有效保护小鼠免于菌血症。噬菌体 SAK84P 的基因组注释显示,该噬菌体中有编码裂解酶的基因,且在基因组注释中尚未发现已知的毒力和耐药基因,后续可纯化该噬菌体的裂解酶蛋白,进行细菌生物膜降解实验[42],为后续体外治疗MRSA 感染奠定基础,也可通过噬菌体改造技术,赋予该噬菌体更多功能,使其更好地在治疗胞内细菌感染中发挥作用,以及预防群体感染和其他超级耐药细菌感染。
猜你喜欢
昆明医科大学学报(2022年2期)2022-03-29
植物保护(2021年4期)2021-11-12
食品安全导刊(2021年20期)2021-08-30
中华养生保健(2020年3期)2020-11-16
科学24小时(2020年4期)2020-05-14
科研成果与传播(2019年3期)2019-09-10
创新作文(1-2年级)(2018年2期)2018-09-13
学习报·教育研究(2017年33期)2017-08-31
食品工业科技(2014年23期)2014-03-11
