
九香虫形态及生物学特征研究
2022-03-24 08:41纪文香李红梅娄灯吉刘宪斌代晰慧施皓译
环境昆虫学报 2022年1期
纪文香,陈 祯,李红梅,娄灯吉,刘宪斌,代晰慧,施皓译
(玉溪师范学院化学生物与环境学院,云南玉溪 653100)
九香虫Coridiuschinensis(Dallas)隶属于半翅目Hemiptera蝽科Pentatomidae兜蝽属Coridius,是重要的药食两用昆虫,《本草纲目》《中药大辞典》《本草新编》和《中华人民共和国药典》等医药典著对其药理药效均有明确记载:“九香虫性味咸、温,归肝、脾、肾经,具理气止痛、温中助阳的功效”(候晓晖,2019)。随着中药现代化研究和虫药研究的不断深入,九香虫已成为治疗多种疾病的重要药物来源,如各类疼痛(包括痛经、心绞痛、风湿关节痛等)、阳痿不育、胃炎、肝炎、胃癌、肝癌、血管瘤、神经衰弱等(刘庆芳, 2002; 侯晓晖等, 2012)。在我国九香虫主要产区贵州、四川等省份民间素有食用九香虫的传统习俗,当地流传着“有钱吃鹿茸,无钱吃打屁虫”等俗谚。无论作为药用昆虫还是保健食品,九香虫都具有良好的应用前景(和韵苹, 2001; 张笠和郭建军, 2011)。目前,对九香虫的生物学、生理生态学以及人工繁育技术等研究不够深入,人工规模化养殖存在诸多技术瓶颈,致使九香虫资源供需矛盾突出,野生资源逐年下降,因此人工养殖九香虫具有良好的商业前景。
近年来,对九香虫的研究主要集中在营养成分分析与评价、药理药效等方面(刘伦沛和郁建平, 2008; 张颖等, 2009; 李俐和李晓飞, 2010; Luoetal., 2012; Lu and Wu, 2013; Shietal., 2014; 郭玉红和贾彬, 2018; Liuetal., 2019; Dingetal., 2020),其生物学研究最早见陈凤玉等对贵阳市花溪区的九香虫生物学进行了初步观察(陈凤玉和杨绪纲, 1985);肖铁光等对湖南地区的九香虫生物学特征进行了报道(肖铁光等, 1998);魏超等对贵州遵义习酒镇九香虫的形态和主要生物学特性进行了研究(魏超等, 2015);对云南省境内九香虫的研究尚未见任何报道。因此,为开展九香虫的规模化人工繁育,本文于2018-2020年间,对云南省玉溪市红塔区九香虫的形态特征及主要生物学生态学特征进行了较为系统的研究。
1 材料与方法
1.1 观察地点
野外观察地点主要位于云南省玉溪市红塔区北城街道后所社区(24°26′29.56″N, 102°32′33.39″E;海拔:1 667 m)和云南省玉溪市红塔区大营街州大河汇溪段旁菜地(24°20′58.58″N, 102°30′15.09″E;海拔:1 642 m),近自然实验种群观察地点位于云南省玉溪市红塔区玉溪师范学院东门苗圃(24°20′16.68″N, 102°33′49.42″E;海拔:1 694 m),在苗圃内树荫旁用尼龙防虫网搭建有一个5.0 m×4.0 m×3.0 m的网室,将盆栽的佛手瓜、南瓜等寄主植物移入网室,之后将野外采集的九香虫成虫接入网室,建立近自然实验种群。
1.2 实验材料
2018年3月15日-20日,自云南玉溪市红塔区采集野生九香虫雌雄成虫,释放于网室内,在网室内堆有柴垛、砖块、瓦片供其越冬,3月下旬始陆续将盆栽的南瓜、佛手瓜苗等寄主植物移入网室内,供其取食。
1.3 九香虫各虫期形态特征观察
在2018-2019年期间,将网室内饲养的九香虫(近自然实验种群)卵、不同龄期若虫、雌雄成虫带回实验室,在Leica M205C体式显微镜下观察和记述各虫期形态特征,并用该显微镜测量其卵的长、宽、高,1~5龄期若虫及雌雄成虫的体长和体宽。
1.4 生活史观察
自2018年1月始,野外观察点每隔5 d调查一次,网室内(近自然实验种群)每隔1 d调查1次,记录九香虫红塔区种群的越冬虫态、越冬起止时间、交配和产卵时间、不同龄期若虫的发育期及成虫羽化期等,连续观察3年,获得该虫在当地的年生活史。
1.5 寄主范围和取食行为观察
在2019年6-8月期间,每隔7 d到玉溪市红塔区野外生境调查1次,每次调查时间为2 d,分上午(7 ∶00 -12 ∶00)、下午(13 ∶00-18 ∶00)、晚上(20 ∶00-23 ∶00)3个时段,调查九香虫的寄主植物范围并观察成虫和若虫的取食行为。除此以外,在此期间,每隔1 d在网室内(近自然实验种群)按以上3个时段观察成虫和若虫的取食行为,作为补充观察。
1.6 取食偏好选择试验
1.6.1九香虫对多科寄主植物的取食偏好选择
2020年8月20日-30日,将先前准备好的7种盆栽寄主植物南瓜Cucurbitamoschat、四季豆Phaseolusvulgaris、番薯Ipomoeabatatas、番茄Solanumlycopersicum、茄子Solanummelongena、辣椒Capsicumannuum、柑橘Citrusreticulata移入网室(长×宽×高=2.0 m× 2.0 m×2.0 m)内,按圆形等距摆放,随后分别接入50头羽化后1日龄成虫和50头4~5龄初若虫,将其置于网室底部中央,从第2日起定时(18 ∶00-20 ∶00)统计不同寄主植物上取食的虫数,持续观察统计10日。
1.6.2九香虫对葫芦科寄主植物的取食偏好选择
2020年9月1日-11日,将预先准备好的6种葫芦科Cucurbitaceae盆栽寄主植物南瓜Cucurbitamoschat、葫芦Lagenariasiceraria、冬瓜Benincasahispida、苦瓜Momordicacharantia、黄瓜Cucumissativus、瓠瓜Lagenariasicerariavar.hispida移入网室(长×宽×高=2.0 m× 2.0 m×2.0 m)内,按圆形等距摆放,随后分别接入50头羽化后1日龄成虫和50头4~5龄初若虫,将其置于网室底部中央,从第2日起定时(18 ∶00-20 ∶00)统计不同寄主植物上取食的虫数,持续观察统计10日。
1.7 主要习性观察
在2018-2020年期间,在网室内及野外生境中观察和记录九香虫若虫及雌雄成虫的主要生物学习性及行为,主要包括取食、求偶、交配、产卵及越冬等行为习性。
1.8 数据分析
实验数据利用SPSS 21.0软件进行单因素方差分析,采用最小显著差数(LSD)法进行差异显著性检验,显著性水平α=0.05。
2 结果与分析
2.1 各虫期形态特征
2.1.1卵(图1-A~D)
不规则圆柱形,两端平截,中段较粗,似鼓状,横卧,长1.1~1.3 mm (n=10),宽1.1~1.2 mm (n=10),高1.1~1.2 mm (n=10)。卵有绿色和粉色两种生态型,绿色型卵壳呈淡墨绿色,粉色型卵壳呈淡粉色,无论哪种色型,中央部分由于卵壳内部的物质衬托颜色较深(图1-C~D)。卵壳质地坚硬,表面具网纹和刻点,近中部有一条由较粗颗粒斜绕而成的环圈(图1-A)。卵盖位于卵的正上方,近圆形,孵化时若虫从此处破壳而出。
2.1.2若虫(图1-E~J)
共5个龄期。1龄若虫(图1-E~F):长1.9~3.2 mm (n=23),宽1.2~2.1 mm (n=23)。头、胸部呈黑褐色,初孵时若虫腹部呈暗红褐色,后逐渐变为暗红色。头部呈梯形,前窄后宽,侧叶明显长于中叶;复眼突出,卵圆形,暗红褐色;触角5节,黑褐色;喙大部为暗褐色,端部为黑色。前胸背板侧缘向外延伸形成2对侧突,前侧突钝圆,有透明感,后侧突尖锐后弯呈刺状,黑色,边缘具微齿,中、后胸及第1腹节侧缘各具1对黑色后弯刺状突起;3对足及胸部腹板均为黑褐色,足末端具1对黄褐色弯钩状的爪和1对几近透明的卵圆形爪垫。腹背中央从基至端部具数块黑色斑块,臭腺孔成对位于斑块两侧;腹背中侧从第2腹节至第7腹节各具1对近圆形白色小斑点,对应腹部边侧各具1对近梯形的小斑块,斑块外缘黑色,内部乳白半透明。
2龄若虫(图1-G):长3.5~4.9 mm (n=21),宽2.0~3.1 mm (n=21)。头、胸部黑褐色,具金属光泽,腹部赭红色。触角第5节端部约4/5为淡黄色或橘黄色,其余各节均为黑褐色;喙大部为淡黄褐色,端部为黑色。前、中、后胸背板向两侧延伸呈黄白色薄片状,外缘呈波浪形;3对足均为黄褐色。腹背中侧从第2腹节至第7腹节各具1对近圆形黄白色小斑点,第1腹节至第9腹节侧缘各具1对半圆形小斑块,其上外缘黑色,其余部分黄白色。
3龄若虫(图1-H):长5.9~7.6 mm (n=13),宽3.9~4.9 mm (n=13)。腹部赭绿色。

图1 九香虫不同发育期形态及生物学特征图Fig.1 Morphology and biological characteristics of different development stage of Coridius chinensis注:A,卵形态;B,卵的排列方式;C,粉色型卵;D,绿色型卵;E,初孵1龄若虫;F,1龄末若虫;G,2龄若虫;H,3龄若虫;I,4龄若虫;J,5龄若虫;K,雌成虫正面观;L,雌成虫反面观;M,雄成虫正面观;N,雄成虫反面观。Note: A, Morphological characteristics of egg;B, Arrangement of eggs;C, Pink colored eggs;D, Green colored eggs;E, Newly-hatched 1st instar nymph;F, Terminal 1st instar nymph;G, 2nd instar nymph;H, 3rd instar nymph;I, 4th instar nymph;J, 5th instar nymph;K, Front of female adult of Coridius chinensis;L, Reverse side of female adult of Coridius chinensis;M, Front of male adult of Coridius chinensis;N, Reverse side of male adult of Coridius chinensis.

续图1 九香虫不同发育期形态及生物学特征图Fig.1 Morphology and biological characteristics of different development stage of Coridius chinensis注:O,雌成虫展翅形态;P,雄成虫展翅形态;Q,若虫的群居性;R,成虫的群居性;S,正在交配的成虫;T,正在产卵的雌成虫;U~X,不同类型的越冬场所。Note: O, Wings spread morphological characteristics of female adult of Coridius chinensis;P, Wings spread morphological characteristics of male adult of Coridius chinensis;Q, Gregarious characteristics of nymphs of Coridius chinensis;R, Gregarious characteristics of adults of Coridius chinensis;S, Mating adults of Coridius chinensis;T, Oviposting adults of Coridius chinensis;U~X, Different hibernaculum of Coridius chinensis.
前、中胸背板稍向两侧延伸,边缘黄白色较平直;3对足从基节至胫节基半段均为黄白色,其余部分为黑褐色。其余特征与2龄若虫相似。
4龄若虫(图1-I):长8.1~11.1 mm (n=27),宽5.0~7.2 mm (n=27)。翅芽微露。其余特征与3龄若虫相似。
5龄若虫(图1-J):长11.2~15.6 mm (n=23),宽7.0~10.0 mm (n=23)。前、中、后胸侧缘平直,呈淡黄或橘黄色。翅芽明显外露,常伸达第3腹节。其余特征与4龄若虫相似。
2.1.3成虫(图1-K~P)
雄虫长15.8~18.4 mm (n=74),宽9.1~10.5 mm (n=74),雌虫长17.8~19.8 mm (n=64),宽10.0~11.5 mm (n=64)。形似六边形,背面较平,腹面显著隆起,多为紫褐色,少见黑褐色,微有铜色和油性光泽。头部略呈梯形,基部宽于端部,头部侧叶长于中叶;复眼突出,卵圆形,黄褐色或深褐色,位于近基部两侧;单眼1对,淡黄色或橘黄色,位于复眼内后侧;喙4节,较短,可伸达前中足基中间;触角线状,5节,约为体长的2/5;第5节端部约4/5多为淡黄色,部分为橘黄色,少为深褐色,其余各节均为黑褐色;第1节较粗短,圆筒形,其余4节较细长而扁,第2节长于第3节,第4节最长。前胸背板和小盾片表面布有许多刻点和不规则的横皱,似波纹状,且在前胸背板和小盾片中央有一条笔直的纵纹贯穿其中;前胸背板发达,前窄后宽,中后部横向隆起,前半部斜向前方,前缘凹进,略呈“V”字形,侧缘较直,略向上卷曲,后缘略拱出,中部横直,侧角、后角显著,后胸腹部近前缘区具1对臭腺孔,位于中后足基间外侧,受刺激时臭腺产生的臭气由此处释放;小盾片发达,显著超过翅爪区长度,长约2/5前翅长度,前段平直,末端钝圆,具明显的横纹;前翅革区位于前3/5,为紫褐色,膜区位于后2/5,为暗黄褐色,具密集纵脉,后翅膜质,基部黄褐色,端部暗褐色,具稀疏纵脉;3对足后足最长,中足次之,前、中、后足跗节均为3节,且第1跗节腹面密被浅黄色毛丛,跗节端部具1对深褐色弯钩状的爪和1对卵圆形的黄色肉质爪垫;雌虫后足胫节内侧具一长椭圆形灰乳白色凹陷,长约2/5胫节长度。翅展开后可见腹部背面基中大部为橘黄色,后端部为紫红色,两侧缘具一窄边,腹部侧接缘及下方侧缘每节中央具有一浅黄色至橘色的斑纹,但小于侧接缘各节的黑色部分;腹部腹面密布刻点及灰白色粉状物,两侧颜色较深,中央部位常为深红色;雄虫腹部具5对气门,雌虫具6对气门。雄虫尾器端缘弧形,中央弓凸。雌虫生殖节末端两侧突起,中央为凹陷状结构。
2.2 生活史
通过长期对玉溪市红塔区九香虫进行野外观察和建立近自然实验种群,获得玉溪市红塔区九香虫的年生活史(表1)。自然条件下,九香虫在该区1年发生1代,越冬成虫翌年5月上旬开始活动,陆续迁至佛手瓜、南瓜等作物上取食,5月上旬开始交配,5月中旬至7月上旬为交配产卵的高峰期,交配产卵会持续到8月上旬,6月上旬至8月下旬陆续孵化出若虫。越冬成虫于5月下旬至8月下旬陆续死亡,新一代若虫于7月下旬至9月下旬羽化为成虫,新一代成虫于9月中旬至10月上旬陆续从佛手瓜、南瓜等寄主植物上迁至越冬场所,进入冬眠状态。
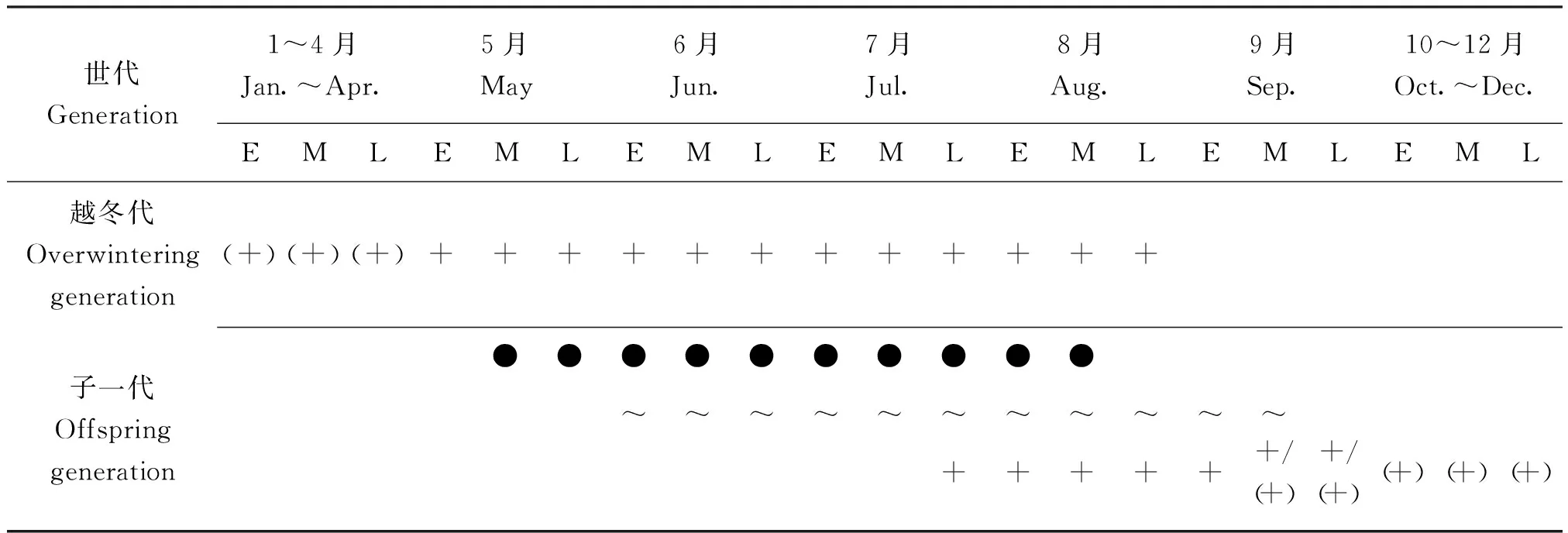
表1 九香虫的生活史(玉溪市红塔区,2018-2020年)
2.3 寄主范围和取食行为
野外调查发现,玉溪市红塔区九香虫可取食的寄主植物有葫芦科佛手瓜Sechiumedule、南瓜、三叶瓜Cucurbitapepo、葫芦、黄瓜、苦瓜、丝瓜Luffacylindrica,旋花科Convolvulaceae番薯,豆科Leguminosae豇豆Vignaunguiculata,茄科Solanaceae茄子和辣椒。据田间观察,九香虫成虫和若虫对佛手瓜和南瓜有明显的偏嗜性,偶见在其他寄主植物上取食。成虫和若虫均有聚集取食的习性,尤其是1~3龄期若虫通常以数头至几十头聚集在一处取食(图1-Q~R)。成虫喜欢在瓜茎上吸食,偶尔会在瓜叶上吸食,若虫则不然,喜欢在瓜叶腹面吸食,少见在瓜茎上吸食,偶尔也会在瓜的藤蔓、花冠和果实上吸食。被口针刺入的瓜茎部位通常会有组织液渗出,伤口处通常还会形成胶质颗粒物。被吸食为害的瓜叶会逐渐变黄,虫量较大时,整株植物会在数日萎蔫枯死。晴天时成虫取食的时段主要集中在上午、下午16 ∶00以后和晚上,阴天可全天取食。取食过程中若遇到惊扰,则立即停止取食,在气温较低时成虫和若虫均会迅速爬离或假死坠落,在气温较高时,遇到惊扰的成虫多会展翅飞逸,但通常只飞行数米便降落。
2.4 九香虫的取食偏好选择
2.4.1九香虫对多科寄主植物的取食偏好选择
九香虫对多科寄主植物的取食偏好选择详见表2。结果表明,若虫和成虫对多科寄主植物的取食偏好存在差异。在设置的7种寄主植物中,若虫最喜取食南瓜,其次分别是:番茄>番薯>四季豆>茄子>辣椒,但不取食柑橘,取食南瓜的若虫数显著大于其它处理组(F=10.984;P=0.000);成虫最喜取食四季豆,其次分别是:南瓜>番薯>番茄>辣椒>茄子>柑橘,取食四季豆、南瓜和番薯的成虫数之间不存在显著性差异(P>0.05),但均与其它4个处理组(番茄、茄子、辣椒、柑橘)存在显著性差异(F=9.422;P=0.000)。从取食的总虫数(若虫+成虫)来看,取食南瓜的虫数最多,其次分别是:四季豆>番薯>番茄>茄子>辣椒>柑橘,取食南瓜的总虫数显著大于其它处理组(F=14.930;P=0.000)。
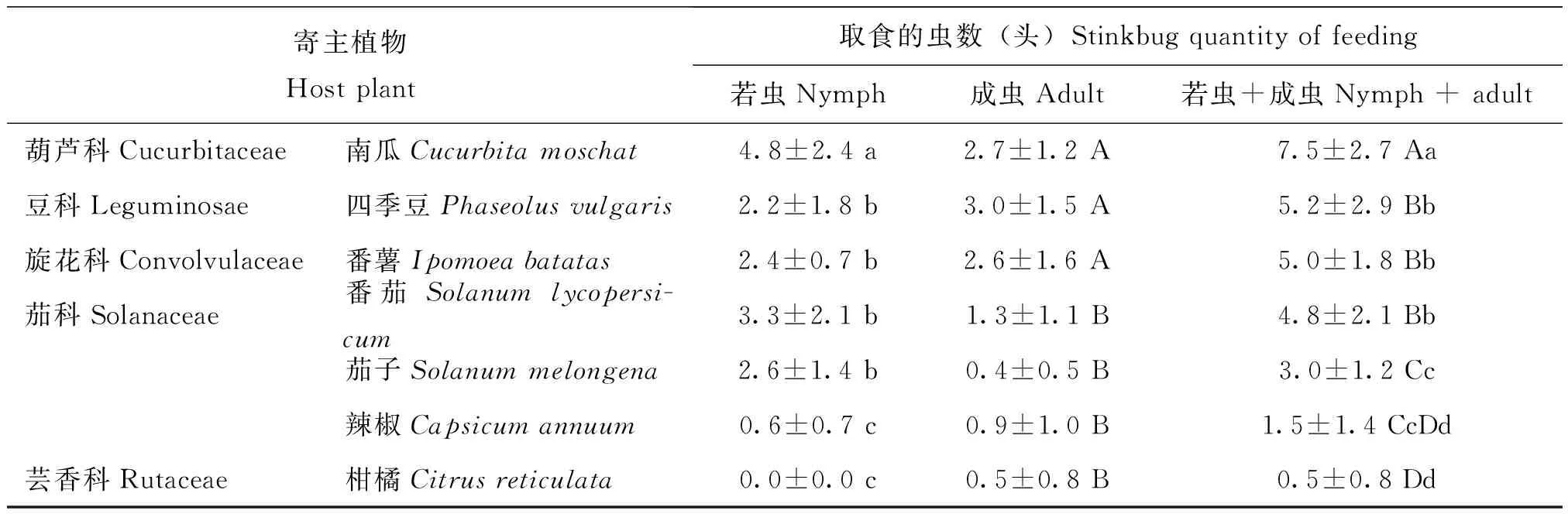
表2 九香虫对多科寄主植物的取食偏好选择
2.4.2九香虫对葫芦科寄主植物的取食偏好选择
九香虫对葫芦科寄主植物的取食偏好选择详见表3。结果表明,若虫和成虫对葫芦科寄主植物的取食偏好也存在差异。在设置的6种葫芦科植物中,若虫最喜取食葫芦,其次分别是:南瓜>冬瓜>苦瓜>黄瓜>瓠瓜,取食葫芦的若虫数显著大于其它处理组(F=62.524;P=0.000);成虫也最喜取食葫芦,其次分别是:冬瓜>南瓜>瓠瓜>苦瓜>黄瓜,取食葫芦的成虫数也显著大于其它处理组(F=99.949;P=0.000)。从取食的总虫数(若虫+成虫)来看,取食葫芦的虫数最多,其次分别是:冬瓜>南瓜>苦瓜>黄瓜>瓠瓜,取食葫芦的总虫数显著大于其它处理组(F=139.654;P=0.000)。

表3 九香虫对葫芦科寄主植物的取食偏好选择
2.5 主要习性
2.5.1求偶和交配
雄成虫主动追逐雌成虫进行求偶。当雄虫找到求偶对象后,会反复用触角触碰雌虫的腹末,整个过程一般会持续数分钟。若雌虫无交配意向,则会短暂停留后迅速爬离,但雄虫通常不会轻易放弃,继续追求,尝试数次未果后才会离去,寻找新的求偶对象;若雌虫有交配意向,则待在原地不动,等待交配的开始。开始时,雄虫会爬上雌虫背部,慢慢调整体向,腹末端伸出阳具,尾部对准雌虫的腹部末端,尝试将阳具伸入雌虫体内,但通常很难一次成功。交配时,雌虫尾部在上,雄虫尾部在下(图1-S),雌雄成虫均具有多次交配的习性。交配从5月上旬开始,5月中旬至7月上旬为交配高峰期,全天可见交配成虫,但通常在早、晚时段交配数量较多,12 ∶00~16 ∶00点之间交配数量较少。交配持续时间长短不一,约1.1~42.0 h,多数选择在瓜茎上边取食边交配,少数会选择在瓜叶上交配,偶尔也会有极少数选择在瓜下的杂草丛和土块等隐蔽处交配。交配过程中若遇到惊扰,雌虫常会迅速拖动雄虫躲到隐蔽处继续交配,但有时也会立即分开,各自逃避或假死坠落。
2.5.2产卵
在自然条件下,九香虫喜将卵产在佛手瓜和南瓜叶腹面和茎干上,在室外建立的近自然种群中,大多数卵也会被产在盆栽南瓜叶腹面和茎干上,但也有少数卵会被产在纱网、杂草和其他杂物上(图1-T)。产卵时雌虫将尾部靠近产卵物体表面,大约每2~4 min产下1粒卵,每产完1粒卵雌虫都会先用后足跗节拍打另一后足胫节内侧的长椭圆形斑块3~7次,之后再同时用1对后足跗节左右拍打卵的两侧数次,然后再换另一后足跗节拍打另一侧后足胫节内侧的长椭圆形斑块,如此交替重复拍打数次。九香虫的卵呈单行排列,单次产卵量可为数粒至30余粒,单次平均产卵量为16.3±6.0粒(n=20),一般随产卵物体纵轴排列。九香虫的产卵具有一定的节律性,产卵一般在16 ∶00-20 ∶00之间进行,单次产卵会持续1 h左右。
2.5.3越冬
在玉溪市红塔区,九香虫以成虫群聚越冬。羽化成虫每年9月中旬至10月上旬(中秋节前后)陆续从佛手瓜、南瓜等寄主植物上迁至越冬场所,进入冬眠状态,翌年4月下旬陆续解除休眠,迁至佛手瓜、南瓜等作物上取食,补充营养。该区的九香虫通常选择在枯树皮下、柴堆、土块、砖(瓦、石)缝、佛手瓜枯藤堆下等隐蔽场所越冬(图1-U~X)。野外调查发现,九香虫对越冬场所的选择具有以下特点:1)就近原则。越冬成虫一般会在原先取食的寄主植物附近寻找合适的越冬场所。同类型的越冬场所,离原先栽种寄主植物的位置越近,越容易找到越冬成虫。2)喜选择能遮风挡雨、干燥弱光的越冬场所环境。越冬期间若天气转暖,偶有成虫会爬出越冬场所在其附近活动。
3 结论与讨论
3.1 九香虫卵的颜色多态性
在近自然实验条件下及红塔区野外,九香虫的卵有绿色和粉色两种色型。通过进一步解剖产卵期的雌成虫卵巢发现,卵的颜色早在其发育期就已形成,且1头雌成虫一生只产一种色型的卵,或为绿色或为粉色,两种色型卵的比例、色型调控机制以及进化适应意义尚不清楚。
昆虫卵的颜色多态性在自然界较普遍存在,尤其在鳞翅目和半翅目昆虫中较为常见。卵的颜色多态性通常被认为是一种进化适应结果,可能与防御捕食者和抵御不良环境有关,其适应功能主要表现在隐匿(crypsis)、警戒色(aposematism)、体温调节(thermoregulation)、光保护(photoprotection)和保水性(water retention)等方面(Guerra-Grenier, 2019)。在自然界,有些昆虫卵的颜色与其周围的背景颜色高度匹配,从而隐藏在环境中不易被天敌(捕食者或寄生者)发现。如螟蛾科Pyralidae和小卷叶蛾科Olethreutidae的一些类群的卵壳呈透明状,卵壳周围背景的反射光使得卵的颜色与附着卵的背景颜色一致,从而使卵不易被捕食者察觉(Peterson, 1962)。Stallings(1986)报道硕大弄蝶Agathymusestelleae的卵有绿色和米黄色两种色型,这两种色型的卵通常都会散落在植物和地面上,绿色型的卵能隐匿在植物上,而米黄色的卵能隐匿在地面上,使得总有一定比例的卵会隐匿在与背景相匹配的环境中,这或许更有利于物种的生存和延续。
而有些昆虫的卵的颜色相比背景环境显得格外鲜艳醒目,这类卵通常具有一定的毒性或不适口性(unpalatability),在适应功能上表现为对捕食者的一种警告信号(warning signals)。Winters等(2014)发现七星瓢虫Coccinellaseptempunctata的卵有黄/橙色型,这两种色型的卵内均含有有毒物质生物碱(alkaloids),生物碱的含量与卵的颜色度量呈正相关。Eisner等(2000)观察到一种草蛉幼虫对灯蛾Utetheisaornatrix产的无生物碱的卵块的捕食率显著高于含生物碱的卵块。本研究认为九香虫卵的颜色二态性(color dimorphism)可能是一种典型的两头下注(Bet-hedging)、风险分散(Risk spreading)的生活史对策:绿色型卵具有与寄主植物颜色背景匹配的隐匿功能,而粉色型卵则具有因含有一定有毒物质成分或不适口性的警戒功能,这需要进一步的相关实验验证。
此外,卵的颜色还与光保护性和保水性有关。Abram等(2015)观察到益蝽Podisusmaculiventris的卵具有从淡黄白色到深褐色/黑色的颜色多态性,有趣的是,浅色型的卵通常被产在植物叶片的腹面,而深色型的卵通常被产在植物叶片的背面,进一步的实验观察发现,当不同色型的卵同时暴露在紫外光下,黑色型的卵比浅色型的卵具有更长的存活时间。昆虫体内的黑色素(melanin)除了具有调节体温和光保护的特性以外,还在昆虫保水性方面扮演重要角色。例如,Farnesi等(2017)发现四斑按蚊Anophelesquadrimaculatus卵壳的黑色素沉积量与其抗脱水性呈正相关。综上,九香虫卵的颜色二态性是否同时具有以上多种适应功能需进一步研究。
3.2 玉溪红塔区九香虫种群的寄主范围和取食偏好
现有文献报道中所记载的九香虫寄主植物主要为葫芦科的各种瓜类作物(陈凤玉和杨绪纲, 1985; 肖铁光等, 1998;侯晓晖, 2019),这与本项研究野外调查以及近自然实验种群取食选择偏好结果基本吻合。陈凤玉和杨绪纲(1985)报道贵阳花溪以及贵州石降、威宁等地的九香虫除主要取食南瓜、丝瓜、短藤瓜等瓜类作物以外, 尚可为害豇豆、菜豆、茄子、烟草、花生、柑桔、苎麻等多种作物。肖铁光等(1998)报道湖南长沙地区的九香虫对南瓜、冬瓜有明显的偏好性,大量田间调查和饲养发现,在南瓜、丝瓜混栽的情况下,能见其低龄若虫在丝瓜上取食,但半月后调查死亡率高达80%,且均很难存活发育到成虫,而逐渐转移到南瓜上取食。本研究对玉溪市红塔区周边的九香虫及其寄主植物进行了调查,在佛手瓜、南瓜、三叶瓜、葫芦、黄瓜、苦瓜、丝瓜、番薯、豇豆、茄子和辣椒藤叶上均发现栖息和取食的九香虫,但其成虫和若虫几乎都只聚集在佛手瓜和南瓜上取食,在以上其它几种寄主植物上偶见取食。而文献报道中所记载的其它寄主植物上未采集到九香虫。地理种群间利用不同的寄主植物,在昆虫中并不少见。有时,即便多种寄主植物同时分布在不同地区,地理种群间也可能利用各不相同的寄主(Singer and McBride, 2010)。不同地理种群间的寄主差异可能是当地综合选择压力的结果,也可能是寄主种类的区域分布差异造成的(Carroll,2008)。
本研究的取食偏好选择实验结果表明,若虫和成虫对寄主植物的取食偏好存在差异。若虫最喜取食南瓜,而成虫最喜取食四季豆,但从取食的总虫数(若虫+成虫)来看,取食南瓜的虫数仍最多,且显著大于其它处理组。显然,在食物匮乏的情况下,若虫和成虫的食性分化可有效避免种内竞争,更有利于种群的生存与繁衍。
猜你喜欢
农业工程学报(2022年10期)2022-08-22
天津农业科学(2022年5期)2022-05-31
植物保护(2021年4期)2021-11-12
饲料博览(2020年11期)2020-12-30
江苏农业科学(2017年12期)2017-09-16
江苏农业科学(2017年7期)2017-05-23
百科知识(2015年7期)2015-04-08
山东农业科学(2014年9期)2015-01-07
江苏农业科学(2014年3期)2014-07-16
