
淡水养殖池塘微型和超微型浮游植物的群落结构组成
2022-03-21 09:57卫鹏毕相东戴伟张达娟李宛津董少杰
大连海洋大学学报 2022年1期
卫鹏,毕相东,戴伟,张达娟,李宛津,董少杰
(天津农学院 水产学院,天津市水产生态及养殖重点实验室,天津 300384)
浮游植物是水生态系统中重要的初级生产者,直接影响着水域的生产性能,在水产养殖业中发挥着重要作用。浮游植物根据粒径大小可以分为3种类型,即小型浮游植物(microphytoplankton,粒径为20~200 μm)、微型浮游植物(nanophytoplankton,粒径为3~20 μm)和超微型浮游植物(picophytoplankton,粒径为0.2~3.0 μm)[1-2]。与小型和微型浮游植物相比,超微型浮游植物具有更小的营养吸收半饱和常数及更高的生长速率[3-4],在水生生态系统物质循环和能量流动中发挥着重要作用[5-6]。超微型浮游植物作为微食物环的起点,被异养鞭毛虫或纤毛虫等原生动物摄食后进入微食物环,再通过桡足类等后生浮游动物的捕食进入经典食物网,在较大程度上影响着渔业产量[7]。然而自20世纪70年代末被发现以来,有关超微型浮游植物的研究主要集中在海洋[8-9]、湖泊[3,5]和水库[10]中,关于养殖水体中超微型浮游植物的研究较少,仅限于养殖海区中少量报道[11],淡水养殖水体中超微型浮游植物研究仍处于空白状态。
加州鲈Micropterussalmoides是中国主要的养殖鱼类之一,具有适应性强、生长快、肉质鲜美且刺少等优点。加州鲈养殖方式以池塘养殖为主,主要通过调控水体中藻相和菌相管理养殖水环境,实现养殖水体物质和能量的良性循环,从而促进加州鲈健康快速生长并减少疾病发生[12]。目前,有关加州鲈养殖池塘藻相的研究[13]与调控[12]主要集中在微型浮游植物,尤其是蓝藻[14],往往忽略超微型浮游植物的影响。而超微型浮游植物大量繁殖亦能形成有害藻华,对水产养殖业造成巨大经济损失[15]。本研究中,对加州鲈养殖池塘微型和超微型浮游植物种群结构组成进行了研究,以期为池塘养殖水体中藻类调控提供参考依据。
1 材料与方法
1.1 材料
采样池塘为天津市蓟州区蓝科水产有限公司加州鲈覆膜养殖Ⅰ号塘和Ⅱ号塘,平均面积约为4 000 m2,养殖密度约为3尾/m2。每2 000 m2池塘面积安装1.5 kW增氧机1台,维持水体溶解氧浓度大于4 mg/L。养殖池塘以持续微流水方式换水,每天交换量为池塘水体的50%。在加州鲈养殖期间,6—7月以投喂冰鲜杂鱼为主,8—9月以投喂鲈配合饲料为主,总体上按照鱼体质量8%投喂冰鲜杂鱼,按照鱼体质量4%投喂配合饲料。
1.2 方法
1.2.1 浮游植物样品的采集 2018年6—9月每月12日采集浮游植物样品一次,每个养殖池塘设3个平行采样点。使用 5 L采水器采集表层(水面以下0.5 m)水样,将采集的水样混匀。
1.2.2 叶绿素a含量的测定 取混匀后1 L水样用3.0、0.2 μm滤膜进行逐级过滤,分别得到微型(粒径>3 μm)和超微型(0.2 μm≤粒径≤3.0 μm)浮游植物,用体积分数95%的乙醇溶液萃取叶绿素a后,采用分光光度计测定叶绿素a含量。
1.2.3 微型浮游植物的定性与定量 取1 L混匀后的水样加入体积分数15%的鲁哥氏试剂进行现场固定,沉淀48 h后,浓缩至50 mL的定量瓶中,之后在光学显微镜下进行浮游植物鉴定和显微计数(参考《中国淡水藻类》[16]和《淡水浮游生物研究方法》[17])。
1.2.4 超微型浮游植物群落结构分析 取混匀后2 L水样分别采用3.0、0.2 μm的滤膜进行逐级过滤,以获得0.2 μm≤粒径≤3.0 μm超微型浮游植物,用于超微型浮游植物总DNA提取。
1.2.5 超微型浮游植物DNA的提取 利用试剂盒(PowerWater®DNA Isolation Kit)提取纯化粒径为0.2~3.0 μm超微浮游植物基因组DNA。每个池塘设3个平行样,共计24个样品。将提取的基因组DNA用10 g/L琼脂糖凝胶电泳进行检测后,置于-20 ℃冰箱中保存待用。
1.2.6 16S rDNA基因高通量测序 按指定测序区域16S rDNA V4区合成特异引物,上游引物为5′AYTGGGYDTAAAGNG 3′,下游引物为5′TACNVGGGTATCTAATCC 3′[18]。PCR体系为25 μL。扩增条件为:94 ℃下变性30 s,50 ℃下退火30 s,72 ℃下延伸30 s,将样本的PCR产物用20 g/L琼脂糖凝胶电泳进行检测,使用AMPure XP Beads (Beckman Coulter)进行产物纯化。得到纯化产物进行2次PCR,连接“Y”字形接头及引物序列编码。利用AMPureXP磁珠纯化筛选目的条带,并通过琼脂糖凝胶电泳对纯化产物进行质控。随后采用Hiseq测序平台对PCR产物进行测序。
1.2.7 18S rDNA基因高通量测序 按指定测序区域18S rDNA V4合成基因扩增引物,上游引物为5′GCGGTAATTCCAGCTCCAATA 3′,下游引物为5′GATCCCCHWACTTTCGTTCTTGA 3′[19]。PCR反应体系(共50 μL):Buffer 5 μL,dNTP Mixture 8 μL,上、下游引物(10 μmol/L)各2 μL,模板DNA 2 μL,Taq DNA聚合酶 2.5 U,加适量灭菌水补足至50 μL。反应条件:94 ℃下预变性3 min;94 ℃下变性30 s,58 ℃下退火复性45 s,72 ℃下延伸45 s,共进行33个循环;最后在72 ℃下再延伸5 min。随后参照与16S rDNA相似的试验流程对最终的PCR产物进行测序。
1.2.8 生物多样性分析 微型浮游植物多样性:基于显微计数后各种微型浮游植物数量进行统计分析。超微型浮游植物多样性:将Hiseq测序得到的原始双端序列数据经过拼接、质控、过滤、去嵌合体,最终得到有效序列,然后根据有效序列数据将97%相似性的非重复序列(不含单序列)进行OTU聚类,采用RDP Classifier贝叶斯算法对97%相似水平的OTU代表序列进行分类学分析,并基于各分类水平中OTU数量统计各样本中的超微真核浮游植物和超微原核浮游植物的群落结构组成。
1.3 数据处理
试验数据采用SPSS Statistics 22.0软件进行相关统计,采用Qiime 2进行扩增子测序数据的处理及相关生物多样性统计分析。
2 结果与分析
2.1 微型和超微型浮游植物对总浮游植物生物量的贡献
从图1可见:加州鲈养殖池塘水体中,微型浮游植物的叶绿素a含量为5.25 ~9.79 μg/L,对总浮游植物叶绿素a的贡献率为82.8%~90.2%,超微型浮游植物的叶绿素含量为0.62~2.03 μg/L,对总浮游植物叶绿素a的贡献率为9.8%~17.2%;微型浮游植物和超微型浮游植物的叶绿素a含量最高值均出现在6月份的Ⅰ号塘和Ⅱ号塘。
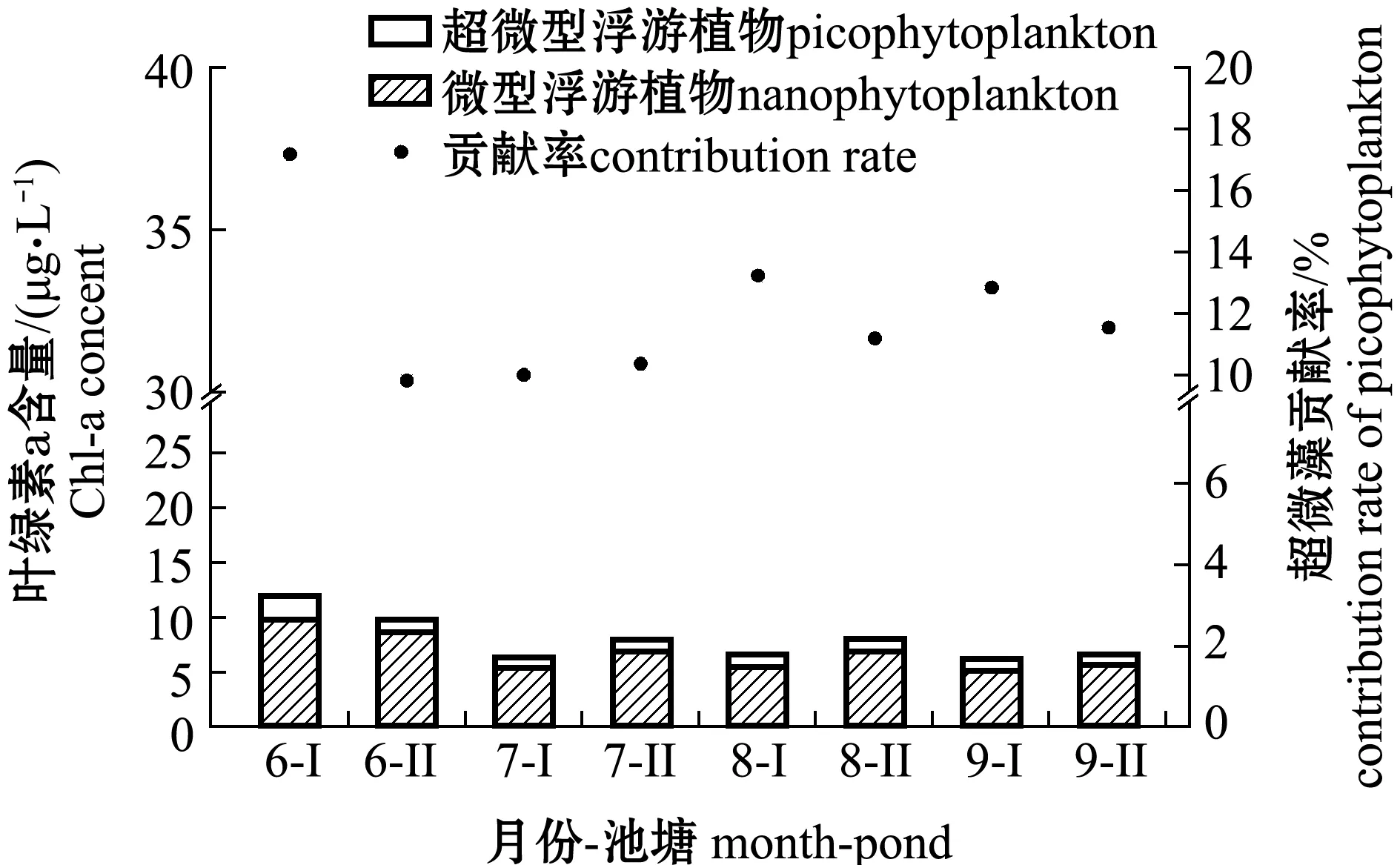
图1 微型和超微型浮游植物叶绿素a含量及对总浮游植物的贡献率Fig.1 Chl-a concentrations and contributions rate to total phytoplankton in nannophytoplankton and picophytoplankton
2.2 微型浮游植物群落结构分析
调查结果显示,Ⅰ号和Ⅱ号养殖池塘的微型浮游植物共有4门24属,主要为蓝藻门Cyanophyta、硅藻门Bacillariophyta、绿藻门Chlorophyta和裸藻门Euglenophyta,其中,蓝藻门包括席藻属Phormidium、颤藻属Oscillatoria等11种,硅藻门包括直链藻属Melosira、针杆藻属Synedra等5种,绿藻门包括新月藻属Closterium、纤维藻属Ankistrodesmus等6种,裸藻门包括裸藻属Euglena和扁裸藻属Phacus2种(表1)。

表1 加州鲈养殖池塘微型浮游植物的种类组成Tab.1 Nanophytoplankton species composition in culture ponds of largemouth bass Micropterus salmoides
进一步分析显示:Ⅰ、Ⅱ号养殖池塘微型浮游植物中,主要以蓝藻门为主,在6—9月,随着时间的推移,其占比逐渐减少,但与其他微型浮游植物相比,其占比始终最大;绿藻门占比则呈逐渐升高趋势,最大占比出现在9月份;硅藻门占比在8月份达到最高值;而裸藻门浮游植物在整个调查期间所占比例变化较小(图2)。
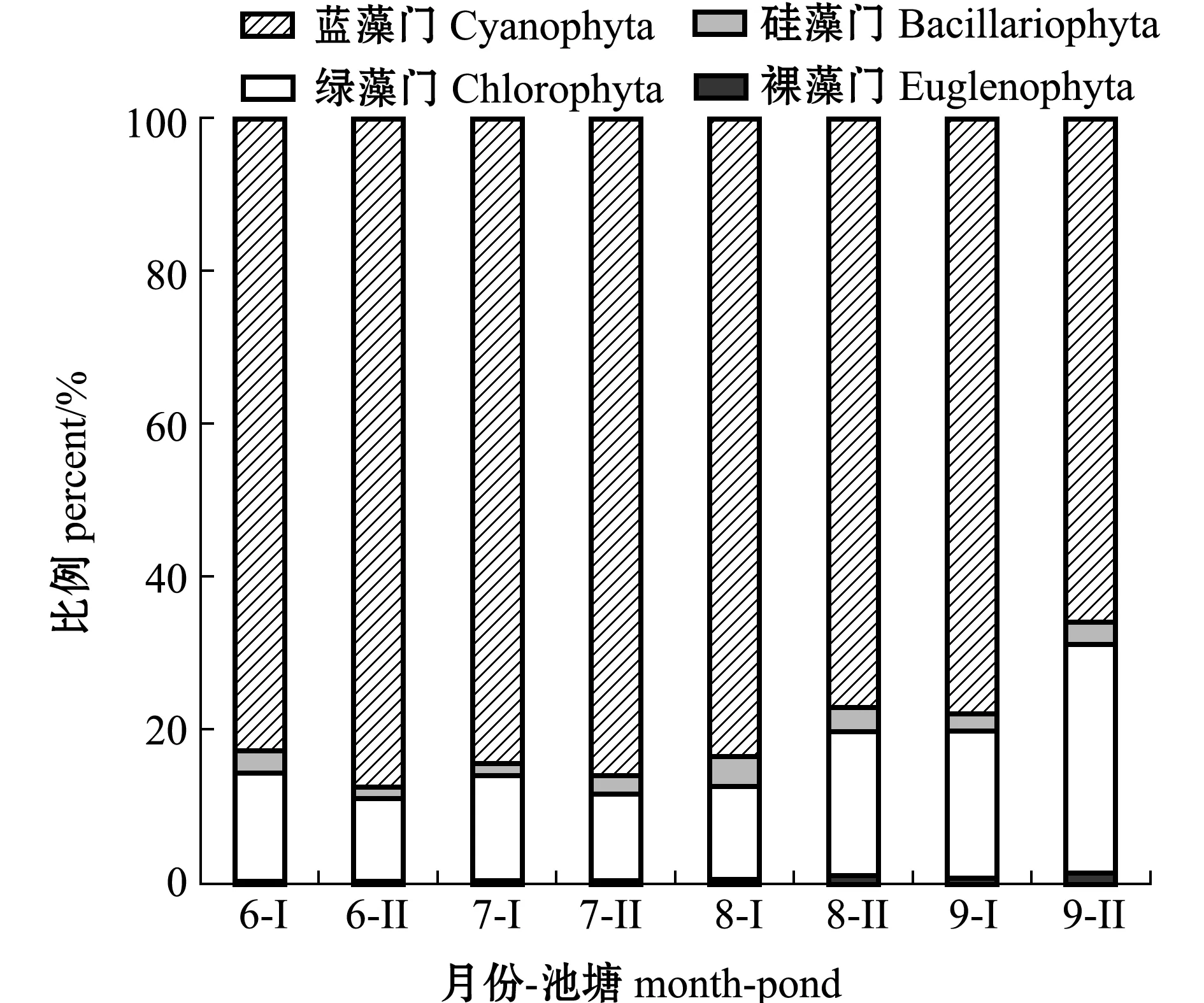
图2 各门微型浮游植物在全部微型浮游植物中的占比Fig.2 Percent of each phylum of nanophytoplankton in the whole nanophytoplankton
2.3 超微型浮游植物群落结构分析
2.3.1 超微原核浮游植物群落组成 对6—9月2个池塘的24个样品进行高通量测序,将分析后得到的235个OTUs进行群落组成分析,结果显示,加州鲈养殖池塘中超微原核浮游植物种类并不局限于聚球藻Synechococcus,且聚球藻丰度占比较小,Ⅰ号和Ⅱ号池塘中聚球藻的相对丰度在6、7月较高,但两池塘聚球藻的相对丰度差异较大,其中,6月Ⅰ号和Ⅱ号池塘中聚球藻的相对丰度分别为2.7%和6.3%,7月分别为3.1%和5.9%,在8、9月其丰度均有下降的趋势(图3)。将占比不足1%的物种归入其他类,除聚球藻属外,还发现微囊藻属Microcystis、浮丝藻属Planktothrix、席藻属、拟柱胞藻Cylindrospermopsis、长孢藻Dolichospermum和节球藻属Nodularia6个藻属,其中,微囊藻属、浮丝藻属和席藻属在整个超微原核浮游植物中的相对丰度均较高,而其余4 个藻属所占比例均较小;微囊藻属和席藻属的相对丰度在8、9月相对较高,而浮丝藻属在6、7月的相对丰度明显高于8、9月;从总体上来看,聚球藻属的相对丰度较其他藻属仍处在较低水平(图3)。
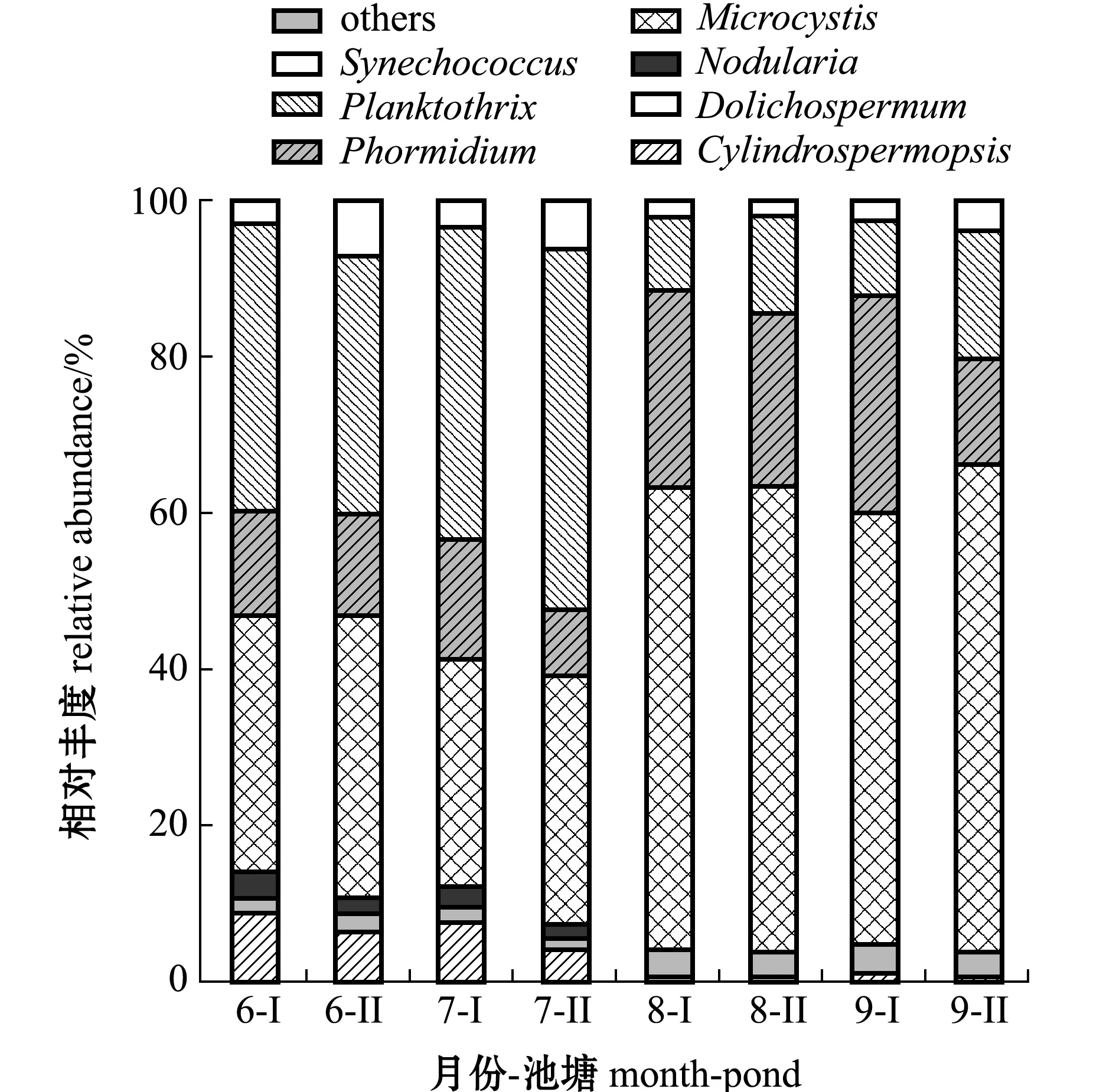
图3 超微原核浮游植物的群落结构Fig.3 Community structure of prokaryotic picophytoplankton
2.3.2 超微真核浮游植物的群落组成 超微真核浮游植物的物种组成非常复杂,多样性极高,因此,本研究中选择在纲级水平上对其进行研究。对养殖池塘6—9月的24个样品进行18S rDNA高通量测序,将分析后得到的896个OTUs进行群落组成分析(图4),其中,将占比不足1%的物种归入其他类,共发现4门7纲,分别为不等鞭毛类Stramenopiles淡色藻门Ochrophyta中的硅藻纲Bacillariophyceae、间藻纲Mediophyceae、真眼点藻纲Eustigmatophyceae 和金藻纲Chrysophyceae,囊泡虫类Alveolata双鞭毛虫门Dinoflagellata中的甲藻纲Dinophyceae,隐藻门Cryptophyta中的隐藻纲Cryptophyceae,以及绿藻门Chlorophyta中的绿藻纲Chlorophyceae。各月份Ⅰ号塘和Ⅱ号塘中均以绿藻纲真核浮游植物占绝对优势,相对丰度为33.4%~56.5%,其次为间藻纲,相对丰度为10.3%~37.9%,硅藻纲、甲藻纲和真眼点藻纲真核浮游植物相对丰度在6、7月相对较高,而金藻纲在8、9月相对丰度明显高于6、7月(图4)。
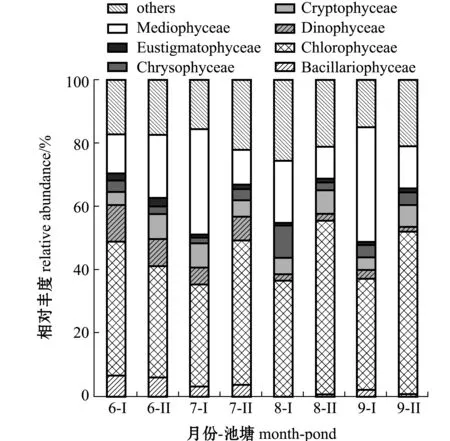
图4 超微真核浮游植物的群落结构Fig.4 Community structure of eukaryotic picophytoplankton
3 讨论
3.1 养殖池塘超微型浮游植物对总浮游植物生物量的贡献
本研究显示,加州鲈养殖池塘中超微型浮游植物对总浮游植物生物量的贡献率为9.8%~17.2%,平均贡献率为12.0%,与中营养湖泊相吻合[20],而微型浮游植物对总浮游植物生物量的贡献率(82.8%~90.2%)明显高于超微型浮游植物。考虑到过滤中的阻滞作用,随着大孔径滤膜孔径逐渐变小,小粒径微藻也可能被截留在大孔径滤膜上。另外,因过滤挤压某些大粒径细胞通过了小孔径滤膜,或某些杆状等大粒径微藻短轴通过孔径也可能偶然截留在小孔径滤膜上[21],滤膜分级过程中无法避免的过滤阻滞作用和过滤挤压作用可能会造成超微型浮游植物对总浮游植物生物量的贡献被低估或高估。浮游植物的生长速率和种群丰度与水环境中营养盐的供应情况及自身对营养盐的吸收速率有关[22]。浮游植物对营养盐吸收的半饱和常数(Ks)与细胞大小成正比,随着浮游植物粒径增大,其Ks也随之增大。在营养盐含量较高情况下,Ks大的浮游植物对营养盐的吸收速率高于Ks小的浮游植物,而在营养盐缺乏情况下,则Ks小的浮游植物对营养盐的吸收速率更高,因此,在贫营养水环境中,个体较小的超微型浮游植物较个体较大的具有更高的营养盐吸收速率和较高的生长速率,并能够在浮游植物群落中占据优势[23-24]。但在营养盐含量丰富的养殖池塘水环境中,微型浮游植物不受营养盐缺乏的抑制,生长速率正常,虽然超微型浮游植物保持同样或更高的营养盐吸收速率和生长速率,但因个体微小故对浮游植物群落的贡献率明显低于个体较大的微型浮游植物。虽然超微型浮游植物在总浮游植物中的比例随着水体营养盐水平的增加而降低,但其生物量与营养条件呈正相关[25]。所以在淡水养殖系统中,当水体贫营养时超微藻对初级生产力和生物量的作用比微型浮游植物更重要。而当水体富营养化后虽然其贡献率下降,但因其自身的生物量和微型浮游植物一样急剧增加,亦会由于强烈的夜间呼吸作用导致养殖水体缺氧,给养殖业带来危害。
目前,淡水池塘养殖加州鲈主要采用冰鲜鱼配合饲料投喂的养殖模式,因冰鲜鱼的投放会导致养殖水体水质有机污染和富营养化。本研究中发现,加州鲈养殖池塘微型浮游植物中,蓝藻一直占绝对优势,在6—9月,其含量逐渐减少,与6—7月以投喂冰鲜杂鱼为主而8—9月以投喂鲈配合饲料为主的投喂方式相符。蓝藻不仅影响加州鲈摄食,同时因其能够产生藻毒素,还会造成加州鲈肝脏受损,影响其生长。另外,蓝藻大量增殖,夜晚呼吸作用过强而大量耗氧,容易诱发加州鲈浮头,而白天由于光合作用过强,溶解氧含量过高,又容易造成鱼体发生气泡病。因此,蓝藻防控一直是养殖池塘加州鲈水质管理的重点。
3.2 养殖池塘超微型原核和真核浮游植物群落结构特征
本研究中,对粒径为0.2~3.0 μm超微型浮游植物16 S扩增子进行高通量测序,结果显示,除聚球藻外还发现包括微囊藻属在内的其他6个藻属。基于池塘养殖水体自身的特殊性,分析其原因可能有以下几方面:(1)进行抽滤的水样中可能存在一些游离蓝藻的环状DNA,因其比一般线性DNA结合更紧密和稳定,致使其最后截留在了0.2 μm滤膜上;(2)某些长孢藻其藻丝呈直线状[26],浮丝藻属也是一种丝状蓝藻[27],这些丝状的大粒径蓝藻短轴可能通过3.0 μm孔径但被截留在0.2 μm滤膜上;(3)本研究中所调查的加州鲈养殖系统采用的是微流水循环系统,因为某些蓝藻可产生内生孢子或外生孢子以进行无性生殖,混在水中的这些孢子随着抽滤过程的进行截留在0.2 μm滤膜上,导致产生误差;(4)由于水体营养盐的变化,相对体积较小的自养型生物吸收和利用营养盐的速率更快,故粒径更小的浮游植物往往具有竞争性优势[28],导致某些藻类出现粒径小型化的发展趋势。因此,可能出于以上几种原因,最终导致测序分析后出现了本不属于超微原核浮游植物的藻属。
对加州鲈养殖池塘24个样品进行18S rDNA高通量测序获得的896个OTUs中,隐藻纲的多性最高,具有16个OTUs,其次是甲藻纲,有8个OTUs,但其相对丰度均低于绿藻纲,而绿藻纲的多样性却较低。谢薇薇等[29]对太湖超微真核藻遗传多样性的研究发现,太湖超微真核藻主要为隐藻门,其次为硅藻门、金藻门和定鞭藻门。吴凡等[3]对抚仙湖超微真核藻的群落结构特征进行研究发现,该湖主要为金藻纲、硅藻纲、甲藻纲等,其中金藻纲占绝对优势。雷瑾等[30]研究发现,白洋淀超微真核藻主要以绿藻纲、硅藻纲、中心硅藻纲、金藻纲为主。而本研究中发现,加州鲈养殖池塘中绿藻纲超微真核浮游植物占绝对优势,其次为间藻纲和隐藻纲,而金藻纲的相对丰度处于较低水平。在加州鲈养殖池塘中未发现常见的定鞭藻门超微真核藻[29],却发现了从黄藻纲中分出来的一个新纲——真眼点藻纲[31]。虽然淡水系统中的超微真核藻近些年才逐渐被关注,但已发现其组成十分复杂,不仅具有环境异质性,且其多样性与水环境营养水平密切相关,此外,其他非生物因素对其群落组成也可能存在重要影响[32]。
4 结论
1)在加州鲈养殖池塘中,微型浮游植物对总浮游植物的贡献率显著高于超微型浮游植物。因为水质有机污染和富营养化,导致微型浮游植物中的蓝藻占绝对优势,随着投喂模式的改变,超微型浮游植物的贡献率有所提升。微型和超微型浮游植物在养殖池塘中起到的作用不容忽视。
2)养殖池塘中除含有典型超微原核藻聚球藻外,还存在有其他6 属超微原核藻,藻类可能出现粒径小型化的发展趋势。
3)超微真核浮游植物相对于超微原核浮游植物,其组成更复杂,多样性更高。
猜你喜欢
沈阳建筑大学学报(自然科学版)(2022年4期)2022-11-15
南京航空航天大学学报(2022年4期)2022-08-30
汽车实用技术(2022年15期)2022-08-19
辽宁化工(2022年3期)2022-04-06
发明与创新·中学生(2022年3期)2022-03-14
西部交通科技(2021年1期)2021-11-09
实验流体力学(2021年3期)2021-07-15
科学家(2021年24期)2021-04-25
建筑建材装饰(2019年5期)2019-09-16
科技与企业(2015年20期)2015-10-21
