
ANA-12通过靶向阻断BDNF/TrkB信号通路降低大鼠的脊髓炎症和缓解病理性疼痛
2022-03-20 03:28:54赵佳佳杨荷雨王招娣朱海丽
南方医科大学学报 2022年2期
脑源性神经营养因子(BDNF)通过与其高亲和力受体—原肌球蛋白受体激酶B(TrkB)相互作用行使其生物学功能,包括细胞分化以及神经发生等。在脑中,BDNF具有多种生理功能,包括神经元的存活、新突触的形成、兴奋性和抑制性神经递质的调节以及突触可塑性的维持等。除了上述生理功能之外,BDNF还参与痛觉的调控,BDNF的异常表达与痛觉行为表现相关。BDNF在神经损伤或炎症相关的痛觉传导区域过度表达,即伤害性途径,主要分布区域包括背根神经节、三叉神经节、脊髓背角以及躯体感觉皮层。BDNF的过度表达对于神经性疼痛和炎症痛的发展是必需的。BDNF/TrkB信号在脊髓中枢敏化中的作用是痛觉维持的基础。BDNF通过激活TkrB而建立慢性痛觉,鞘内注射TrkB受体阻断剂能够阻抑外源性BDNF引起的异常性疼痛。研究显示,BDNF/TrkB信号的激活参与伤害信号传递的调控。目前发现与BDNF/TrkB信号途径相关的分子途径和调节因子包括PAR2-NF-κB、P2X4R-TLR4-p38 信号以及BDNF 转录调节因子CREB等。在阐述痛觉敏感性增加的机制方面,多数研究聚焦于胶质细胞。在多种信号途径中,已明确P2X4R-TLR4-p38信号在特定细胞类型,即小胶质细胞中发挥调节作用;对于星形胶质细胞,主要涉及炎症反应及炎症因子的表达调节。但目前对于小胶质细胞在骨癌痛相关的炎症反应中的作用及机制尚未完全明确。
在脊髓和脑中,各种受体系统的激活以及各种化合物的合成参与痛觉调控。脊髓小胶质细胞不仅参与了损伤引发的神经性疼痛的发生,并且也是维持超敏反应所必需的。BDNF在神经病理性疼痛晚期介导小胶质细胞-神经元相互作用的信号分子。在疼痛晚期,中枢炎症反应明显减弱,小胶质细胞主要依赖营养因子BDNF来延长疼痛超敏反应。目前研究主要通过调控BDNF/TrkB下游信号途径在胶质细胞的作用,抑制胶质细胞激活并减少炎症因子或神经营养因子的分泌,以达到改善动物痛觉行为的目的。
BDNF的异常过度表达与动物的痛觉行为表现密切相关,相关研究表明ANA-12可下调BDNF参与/诱导的神经炎症以及病理性疼痛;此外,ANA-12还参与慢性疼痛状态维持的调控。上述研究表明,ANA-12可通过抑制BDNF/TrkB信号参与多种痛觉行为的调控,但目前关于ANA-12在痛觉调控中的内在分子机制尚未明确。因此,为探究ANA-12在急性疼痛及慢性疼痛中的作用,小胶质细胞激活与动物痛觉行为的关系,本研究采用鞘内给药TrkB受体拮抗剂ANA-12靶向阻断BDNF/TrkB信号,进而分析BDNF/TrkB信号活性、小胶质细胞的激活状态以及炎症因子的表达情况,明确BDNF/TrkB信号在病理性疼痛中的作用及机制。
1 材料和方法
1.1 实验动物及分组
本实验使用42只雄性Sprague-Dawley(SD)大鼠(湖北省实验动物中心),体质量180~200 g,6~8周龄。大鼠饲养在室温为22±2 ℃的环境中,以12/12 h昼夜节律变换并提供充足的水和食物。将大鼠随机分为两大组,BDNF 处理组(4 小组,6 只/组):包括对照组(control)、ANA-12 处理组(ANA-12)、BDNF 处理组(BDNF)以及BDNF 和ANA-12 联合处理组(BDNF+ANA-12);炎症痛模型组(3小组,6只/组):包括对照组(control)、CFA处理组(CFA)以及CFA和ANA-12联合处理组(CFA+ANA-12)。本研究所有动物实验均经湖北科技学院实验动物伦理委员会批准(2020-01-900)。
1.2 主要试剂
蛋白酶抑制剂、Radio Immunoprecipitation Assay裂解液、SDS-PAGE凝胶快速配制试剂盒、BCA蛋白浓度测定试剂盒(上海碧云天生物技术有限公司);BDNF(北京义翘神州科技股份有限公司);ANA-12(Selleck);完全弗氏佐剂(CFA)(Sigma-Aldrich)。本实验所用I抗如 下:anti-BDNF(A4873)、anti-TrkB(A19832)、phospho-TrkB-Y705(AP0423)、anti-Iba1(A1527)和anti-TNF-α(A0277)(武汉爱博泰克生物科技有限公司)。所用Ⅱ抗为HRP标记山羊抗兔IgG(武汉爱博泰克生物科技有限公司)。
1.3 模型建立及分组处理
BDNF-急性痛大鼠模型:大鼠随机分为对照组(control)和模型组。首先大鼠背部注射部位进行剃毛和皮肤消毒处理,然后用无菌微量注射器于L5和L6脊椎棘突间隙垂直进针,进入蛛网膜下腔,将注射器缓慢倾斜45°方向。模型组大鼠注入10 ng/10 μL BDNF溶液,对照组大鼠注射相同体积(10 μL)的生理盐水。给药处理如下:模型组大鼠随机分为溶剂对照组(BDNF)和ANA-12给药组(BDNF+ANA-12)。BDNF注入30 min后,溶剂对照组鞘内注射10 μL体积的DMSO+生理盐水(体积比1∶1),ANA-12组注射200 μg/10 μL体积的ANA-12溶液。
将大鼠置于30 cm×30 cm×30 cm的透明有机玻璃盒内,实验平台高50 cm。实验过程中保持环境安静。实验开始前,将大鼠放入玻璃盒中适应30 min,观察并记录每5 min内大鼠自发性缩足(或舔爪)反射次数,记录3次,5 min/次。脊髓鞘内注射BDNF后,将大鼠放回盒内适应30 min后,记录每5 min内缩足(或舔爪)次数,记录3次。继续鞘内注射溶剂或ANA-12溶液,放回盒内适应30 min后,记录每5 min内缩足(或舔爪)次数,记录3次。
1.4 大鼠自发性缩足反射次数记录
CFA-慢性炎症痛大鼠模型:大鼠随机分为对照组(control)和模型组。模型组大鼠采用水合氯醛麻醉,将CFA按1∶1的比例与生理盐水混匀为悬浊液,将该悬浊液皮下注入大鼠左后肢足底,使其局部产生红肿热痛等炎性反应。对照组注射相同剂量的生理盐水。给药处理:模型组大鼠随机分为溶剂对照组(CFA)和ANA-12给药组(CFA+ANA-12)。CFA模型构建7 d后,溶剂对照组鞘内注射10 μL体积DMSO+生理盐水(体积比1∶1),ANA-12组注射200 μg/10 μL体积的ANA-12溶液。
1.文章标题分析。优质推文的标题总体上具有以下特点:一是情感倾向明显,语气强烈。其中标题中含有感叹号的推文有32篇,占25.6%,有4篇推文的标题包含多个感叹号,以使感情表达更为强烈,如河北民建发布的《公职人员请注意!这些饭局不能去!!》;标题中含有问号的推文有7篇,更容易吸引读者的注意和兴趣,如河北民建发布的《都是共产党的官员,差距咋这大?》。二是成句形式活泼多样,多使用网络流行语。如九三学社之声发布的《确认过眼神!都是九三青年人!》,中国农工民主党发布的《【福利来袭】@所有人 喜迎新春,海量红包等拿!》。
BDNF与TrkB 的细胞外结构域结合,诱导受体二聚化,导致细胞内结构域内的酪氨酸(Y705)激酶活性和自磷酸化增加,依次触发下游信号通路的激活。通过检测TrkB在Y705位点的磷酸化水平以表征其活性变化情况。蛋白质免疫印迹结果显示,BDNF和CFA处理均诱导脊髓组织中磷酸化TrkB(Y705)水平增加(<0.05);经ANA-12处理后BDNF或CFA组TrkB(Y705)磷酸化水平降低(<0.05,图3)。
笔者结合多年心理咨询案例、历年的学生就业压力调查以及学生管理工作中特殊学生的问题处理,发现高职生的心理健康问题不仅需要学校高度重视,而且需要家长充分发挥作用,需要家校合作才能取得成绩。通过查阅文献资料发现,家校合作方面的实践性研究并不多。
1.5 50%缩足阈值(PWT)检测
与对照组相比,BDNF 处理后缩足次数上升(<0.05)。进一步进行鞘内给药ANA-12并记录大鼠自发性缩足(或舔爪)反射次数,结果显示,与BDNF组大鼠相比,ANA-12处理降低大鼠自发缩足次数(<0.05)。ANA-12处理对正常大鼠的行为学影响无统计学意义(>0.05,图1)。
1.6 免疫印迹法检测脊髓组织蛋白质表达
冠心病作为常见多发疾病,可见血液液体成分逐渐减少,且固体成分不断增多,造成血液黏度升高。此疾病好发于中老年人群,患者血管壁逐渐失去弹性,管腔越来越狭窄,进一步加重血粘度升高程度[2-3],血液流速随之减慢,引发心肌缺氧,出现冠状动脉血栓、心绞痛,故而,研究血液流变学变化情况对冠心病治疗、预后评估等均有重要意义。
2016年1月~2018年8月,本院骨科共收治年龄≥60岁患者1 381例,其中1 336例符合上述标准,纳入本研究。男520例,女774例,年龄60~98岁,平均(68.86±7.58)岁。
1.7 统计学方法
采用SPSS 26.0软件进行统计学分析。行为学数据资料以均数±标准误表示,免疫印迹数据资料以均数±标准差表示。组间比较采用单因素方差分析,<0.05为差异具有统计学意义。
2 结果
2.1 BDNF/TrkB信号活性抑制缓解大鼠BDNF-急性痛
各组大鼠给药处理后0、2、4和6 h,根据up-down法检测50%PWT。方法如下:将动物放置于行为测试笼内,待大鼠适应30 min后,用Von Frey纤维(0.4~26 g)垂直剌激大鼠左侧后肢足底正中,持续时间≤5 s,出现舔足或者抬足等行为视为阳性反应,反之则为阴性反应。记录6 次数据后结束检测。根据公式50%PWT(g)=(10)/10000计算各组大鼠的缩足阈值。(Xf为本次行为学测试中使用的最后一个Von Frey纤维的阶数,K为痛觉阈矫正系数,δ为相邻不同纤维刺激之间的差异。)
行为学记录结束后,给予过量麻醉剂处死大鼠,分离脊髓组织于预冷的PBS中,取腰段脊髓置于含有蛋白酶抑制剂的RIPA裂解液中在冰上进行组织匀浆。收集组织匀浆液于离心管中,4 ℃条件下12 000 r/min离心15 min后取上清,加入1/3体积的4×上样缓冲液,沸水中加热10 min变性。样品的蛋白质浓度用BCA蛋白分析试剂盒进行检测。SDS-PAGE电泳分离蛋白质样品,将蛋白质电转至PVDF膜上,5%脱脂奶粉室温封闭l h,Ⅰ抗4 ℃孵育过夜。所用Ⅰ抗及比例如下:anti-BDNF(1∶500)、anti-TrkB(1∶500)、phospho-TrkB-Y705(1∶500)、anti-Iba1(1∶500)和anti-TNF-α(1∶500)。Ⅰ抗孵育之后,洗膜3 次,并用Ⅱ抗室温孵育1 h,然后用LAS500凝胶成像系统扫描观察,ImageJ软件分析条带灰度值。
通过足底注射CFA建立炎症疼痛大鼠模型。注射CFA导致大鼠同侧后爪对机械刺激的敏感性增加,痛觉阈值降低(<0.05)。与CFA组比,ANA-12处理2、4、6 h后,大鼠缩足阈值升高(<0.05,图2A)。CFA处理对大鼠对侧后爪的影响无统计学意义,机械痛阈值在ANA-12处理前后的差异也无统计学意义(>0.05,图2B)。

2.2 BDNF/TrkB信号活性抑制缓解大鼠CFA-慢性炎症痛
为检验该清洁机器人能否满足实际光伏电站的使用需要,利用学校和合作单位提供的现场试验条件,对样机进行了运动功能和清洗作业实验。
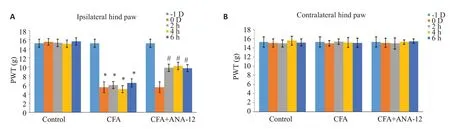
2.3 ANA-12鞘内给药阻断BDNF/TrkB信号
Suggestions on Xuzhou constructing science and education center

2.4 BDNF/TrkB信号活性抑制减轻脊髓小胶质细胞活化并抑制脊髓炎症
免疫印迹分析显示BDNF和CFA诱导脊髓组织中小胶质细胞的特异性标记物Iba1和促炎细胞因子TNF-α表达上调(<0.05,图4),同时炎症小体NLRP3 和caspase-1 以及炎症组分IL-1β表达增加(<0.05)。ANA-12处理后,Iba1、TNF-α、IL-β、NLRP3和caspase-1表达水平下降(<0.05,图5)。


3 讨论
疼痛刺激以外周痛觉感受器的激活以及将“痛觉信号”传递到躯体感觉皮层为主要特征,“痛觉信号”传递需要至少3种不同类型的神经元。BDNF是痛觉信息传递中的重要神经调质,BDNF参与了痛觉途径的初始敏化,在损伤的早期阶段的增加可能是适应性;然而,BDNF的持续释放会触发一种“不适应的”和有害的影响,从而维持或导致慢性疼痛。本研究运用BDNF和CFA处理建立动物痛觉模型,从而探究其中BDNF信号在动物痛觉行为调控中的作用及机制。
BDNF在痛觉传导通路中的异常过度表达与动物痛觉行为表现呈正相关性。在外周炎症中,BDNF在DRG神经元水平的增加与脊髓背角释放增加相关,表明BDNF信号在炎症性疼痛中发挥调节作用。本研究中,BDNF鞘内给药处理可显著诱导正常大鼠(即对照组大鼠)痛觉敏感性增加,表现为痛觉过敏。另外,本研究在BDNF 和CFA-诱导的痛觉模型中均观察到TrkB信号的表达和活性均上调,而ANA-12处理可下调TrkB受体的表达及活性且降低动物的痛觉敏感性。这表明抑制痛觉传导通路中的BDNF-TrkB信号的活性可改善动物的痛觉行为。
神经胶质细胞,包括小胶质细胞和星形胶质细胞以及它们的相互作用参与神经炎症过程的调控。小胶质细胞被认为是神经炎症反应的主要来源,星形胶质细胞可以被小胶质细胞通过补体级联反应激活并进一步引起神经毒性,因此,星形胶质细胞的激活也被认为是神经炎症的一个重要调控因素。生理状态下小胶质细胞处于静息态,感知和监测周围环境的变化,这使小胶质细胞能够有效地控制微环境,清除积累的代谢产物和恶化的组织成分;外周神经损伤后,脊髓中小胶质细胞活化,大量合成释放诸多疼痛因子(白介素-6、肿瘤坏死因子、γ-干扰素、BDNF等)参与神经病理性疼痛的发生及维持。有炎症脊髓水平研究显示,CFA诱导的动物模型中,BDNF和促炎细胞因子TNF-α、IL-1β、IL-6的表达均上调。本研究中,脊髓组织蛋白质水平分析表明BDNF 和CFA 处理均可激活小胶质细胞,上调NLRP3炎症小体的表达,同时增加炎症因子的表达。结合上述研究提示,不论在正常动物还是炎症痛模型动物中,BDNF的过度表达均参与痛觉的诱导、形成和维持过程。BDNF可通过激活多种信号通路参与小胶质细胞介导的炎症性疼痛反应。BDNF可诱导脊髓小胶质细胞中PI3K和ERK的激活,该效应是神经性疼痛发展的基础。另外,BDNF调节PKCλ和PKMζ的形成以及PKMζ的磷酸化,BDNF/aPKC信号形成了维持集中慢性疼痛状态所需的信号轴。提示本研究中BDNF和CFA诱导的痛觉模型中,小胶质细胞的激活以及炎症因子的表达可能由上述激酶途径介导。BDNF通过与其高亲和力受体TrkB结合介导并激活下游信号通路参与疼痛调节,TrkB抑制剂可显著下调痛觉模型动物的痛觉敏感性并改善痛觉行为。本实验通过对已经存在神经病理性疼痛的大鼠鞘内应用ANA-12,有效的拮抗脊髓背角BDNF/TrkB信号的过度激活,即TrkB活性位点(Y705)磷酸化水平下调,大鼠痛觉过敏症状得到改善,进一步说明,BDNF在疼痛维持过程中起重要作用以及ANA-12的镇痛作用。
综上所述,BDNF/TrkB信号调节脊髓痛觉信息传递,TrkB受体拮抗剂ANA-12下调BDNF/TrkB信号从而抑制脊髓小胶质细胞的激活并下调炎症因子的表达水平,进而缓解动物痛觉,发挥镇痛作用。本研究作用于痛觉信号的源头BDNF/TrkB,探究小胶质细胞与炎症信号的相关性在痛觉行为学中的调节作用及机制,为相关疾病治疗靶标的选择提供了新的方向,但具体的分子机制还有待深入研究。
猜你喜欢
军事文摘(2022年8期)2022-11-03 14:22:01
昆明医科大学学报(2020年12期)2021-01-26 00:44:30
神经损伤与功能重建(2020年11期)2020-12-01 05:01:54
湖南中医药大学学报(2016年1期)2016-12-01 04:08:21
中国运动医学杂志(2016年3期)2016-07-10 12:07:23
癌症进展(2016年11期)2016-03-20 13:16:06
中外医疗(2015年18期)2016-01-04 06:52:02
磁共振成像(2015年1期)2015-12-23 08:52:21
中国当代医药(2015年31期)2015-03-01 02:08:30
中国中医药现代远程教育(2014年21期)2014-03-01 04:32:11
