
大蒜素调控黑腹果蝇产卵偏嗜性和适合度
2022-03-17 01:58周思艺陈利荣威1
昆虫学报 2022年2期
周思艺, 夏 静, 闫 琴, 芦 韬, 陈利荣, 刘 威1,
(1. 安徽农业大学植物保护学院, 合肥 230036; 2. 山西医科大学汾阳学院临床医学系, 山西汾阳 032200;3. 山西医科大学汾阳学院医学检验系, 山西汾阳 032200; 4. 中山大学肿瘤中心病理科, 广州510000)
大蒜为百合科葱属植物的地下鳞茎,具有强烈辛辣的蒜臭味,含有丰富的蛋白质、低聚糖和多糖类、矿物质等,因此成为膳食中一种常用的调料。大蒜不仅可作调味料,还含有多种生物活性化合物,如有机硫化物、多糖等,可入药,是著名的食药两用植物(Shangetal., 2019)。大蒜素(allicin)是从大蒜鳞茎中提取的一种有机硫化合物,主要成分为三硫代烯丙醚类,淡黄色粉末或油状液体。大蒜素现已被证实为大蒜主要的活性成分,具有广谱抗菌、降脂降血糖和增强机体抵抗疾病能力等作用(张庭廷等, 2007)。然而,大蒜素具有强烈的臭味,并且接触后会引起辛辣的痛觉(Macphersonetal., 2005)。有研究表明,欧洲椋鸟Sturnusvulgaris对添加大蒜油的食物表现出明显的厌恶行为(Hileetal., 2004)。因此我们推测,大蒜素可能会对昆虫产生一定的驱避作用。
黑腹果蝇Drosophilamelanogaster是大家熟知模式生物之一。为了物种的繁衍延续和满足后代良好的生存环境,雌果蝇要综合温度、湿度、营养条件、毒素、天敌等多种因素选择合适的产卵地点,从而有效躲避有毒物质的侵害,促进后代的生长发育、提高存活率,因此产卵决策是一个优秀的行为模型(Yangetal., 2008; Aietal., 2010; 王璐璐等, 2017)。本研究旨在探究大蒜素对黑腹果蝇决策行为的影响,以寻找适宜的环境并躲避大蒜素的危害。
果蝇会通过不同类型的受体神经元对外界刺激物做出强烈的反应,故常被选作为研究对象(Liuetal., 2017; 高露等, 2019)。各种感觉由其对应的感觉神经元所控制,这些不同的神经元接收到刺激后引起机体不适的感受,并最终作出相应的反应。Transient receptor potential ankyrin 1(TRPA1)是一种瞬时受体电位的非选择性阳离子通道,是一种多模态损伤传感器,可检测多种损伤信号,包括寒冷、化学和机械刺激(Bautistaetal., 2013; Zygmunt and Högestätt, 2014)。TRPA1在检测不同物种的有害化学物质方面高度保守,它可以被许多有害的活性化学激动剂直接激活,如异硫氰酸烯丙基(AITC)、肉桂醛(Bandelletal., 2004; Jordtetal., 2004)和大蒜素(Bautistaetal., 2005; Macphersonetal., 2005)等。这些化学激动剂主要包括亲电性和非亲电性激动剂两大类。亲电性激动剂如大蒜素,通过对TRPA1上的半胱氨酸残基共价修饰来开放TRPA1(Hinmanetal., 2006; Macphersonetal., 2007);而非亲电性激动剂如香芹酚,被认为是通过直接与通道结合来激活TRPA1(Xuetal., 2006; Zygmunt and Högestätt, 2014)。但是该受体是否介导对大蒜素产卵避性仍然未知。因此,本研究利用果蝇产卵决策模型,研究果蝇能否感受大蒜素而调整产卵行为,探究介导这一行为的感觉系统,并进一步探究大蒜素对果蝇后代的发育历期和存活率以及对成虫寿命和健康的影响。
1 材料与方法
1.1 果蝇品系与培养
野生型黑腹果蝇品系Oregon R(OR)和Canton S(CS)来自本实验室保种库,突变体OdorantReceptorCo-Receptor2(Orco2)和TrpA1由Bloomington果蝇品系库所提供。Orco2突变体对绝大多数气味丧失感知(Larssonetal., 2004),TrpA1突变体丧失对刺激性物质的感知(Bautistaetal., 2013; Zygmunt and Högestätt, 2014)。
若无特别说明,所有果蝇均在温度25℃、相对湿度60%的恒温箱中培养,培养光周期为12L∶12D (Liuetal., 2012)。通常情况下,果蝇用玉米粉-酵母培养基饲养,为避免酵母对果蝇产卵行为的影响,行为分析使用的培养基以水解酪蛋白代替酵母。
1.2 试剂与仪器
1.2.1试剂:大蒜素(源叶生物,98%);蔗糖(光复,合格);酪蛋白;琼脂;玉米粉。
1.2.2仪器:25℃恒温箱(恒丰, WS-01Y);天平(Denver Instrument, XE-310);显微镜(Olympus SZX7; Nikon SMZ745T)。
1.3 黑腹果蝇产卵行为检测
实验选用中空透明圆柱体为产卵双选择装置,高度为80 mm,直径为60 mm,底部放入果蝇产卵的培养皿(Josephetal., 2009)。涂有酵母膏的葡萄汁板(700 mL水,30 g琼脂,300 mL葡萄汁,50 g蔗糖)育肥果蝇2~3 d,换为果蝇产卵所用培养基,产卵时间为12 h。含有0.01%, 0.015%和0.2%大蒜素的食物(实验组)与玉米-酪蛋白食物(1 000 mL水,70 g玉米粉,50 g蔗糖,15 g酪蛋白,15 g琼脂,对照组)各一半,拼接成果蝇产卵培养基,并将25头雌果蝇和少许雄果蝇放于装置中,观察其产卵行为。实验重复4次。产卵12 h后分别统计两边培养基上卵的数量,产卵指数(OI)=(实验组卵数-对照组卵数)/(实验组卵数+对照组卵数)。强迫产卵试验则是果蝇经育肥2~3 d,然后转移到加有大蒜素的食物上产卵12 h,统计卵数,计算果蝇单雌产卵量:单雌产卵量=总产卵量/雌性头数。
1.4 黑腹果蝇位置效应检测
将50头黑腹果蝇雄成虫、250头雌成虫放入高140 mm、直径100 mm的产卵双选择装置,待果蝇静置适应30 min后,每隔10 min分别计数两侧实验和对照组(同1.3节)培养基上的果蝇数量,连续记录2 h。 实验重复4次。 位置指数(position index, PI)=(实验组果蝇数目-对照组果蝇数目)/(实验组果蝇数目+对照组果蝇数目)(Josephetal., 2009)。
1.5 毛细管摄食法(capillary feeder, CAFE)检测黑腹果蝇的摄食行为
将20~40头黑腹果蝇成虫置于玻璃瓶(高90 mm、直径20 mm)中禁食16 h,玻璃瓶底部为ddH2O浸润的滤纸以满足饮水需求。然后将果蝇转移到一个新的玻璃瓶中进行摄食试验。毛细管插入塞子,内加入等量两种液体,一侧为不同浓度的大蒜素溶液(实验组)(同1.3节),一侧为水(对照组),插入毛细管的深度均为2 mm。允许果蝇在环境照明条件下进食30 min,毛细管弯液面下降长度作为食物消耗。实验重复4次。摄食指数(feeding index, FI)=(含化学试剂的溶液消费量-仅含水的消费量)/总消费量(Jaetal., 2007)。
1.6 黑腹果蝇前足手术
用解剖镊除去OR黑腹果蝇成虫部分前足,使其部分味觉感受器丧失,待24 h后检测其产卵决策行为。实验重复4次。
1.7 Y迷宫实验
将雌/雄黑腹果蝇成虫各20头置于小管中,饥饿处理16 h,在Y迷宫的两头放入含有大蒜素的玉米粉食物,在另一头将果蝇放入Y迷宫中,处理6 h,果蝇转移时在冰上将其麻醉,6 h后计数Y迷宫两边果蝇数目(Lietal., 2020)。实验重复3次。
1.8 黑腹果蝇触角手术
用解剖镊除去OR黑腹果蝇成虫触角,使其部分嗅觉感受器丧失,待24 h后检测其产卵决策行为。实验重复4次。
1.9 黑腹果蝇的发育历期及存活率统计
将适量黑腹果蝇成虫放入收集器中,在葡萄汁板上涂抹酵母膏,收集8 h内果蝇所产的卵,将其置于含有不同浓度(同1.3节)大蒜素的小管食物中。每一小管食物上放置50枚卵。每天计数每组小管中成蛹和成虫的数目(n=50)。实验重复4次。T=(T1×N1+T2×N2+…+Tm×Nm)/(N1+N2+…+Nm),其中T表示对应阶段发育历期,Tm表示从产卵后到对应阶段需要的天数,Nm表示第Tm天形成对应阶段果蝇的数目。统计计算每组成蛹时间(卵产出至成蛹时间)和成虫羽化时间(卵产出至成虫羽化时间)。存活率=成蛹或羽化的数目/50×100%(高露等, 2019)。
1.10 黑腹果蝇死亡率统计
将20头雌/雄黑腹果蝇成虫置于含有不同浓度(同1.3节)大蒜素的小管中,每天计数小管中果蝇数目(n=20)。实验重复4次。死亡率=雌/雄果蝇数目/20×100%。
1.11 黑腹果蝇的奔跑攀爬能力检测
将20头黑腹果蝇成虫置于含有5%蔗糖溶液和0.1%大蒜素溶液的小管中,处理48 h后,置于有刻度的玻璃量筒中,计算它们的攀爬和奔跑速度(Lietal., 2020)。实验重复4次。
1.12 ROS染色实验
挑取酵母玉米培养基中OR黑腹果蝇成虫(羽化后4 d)各20头,分别饲喂5%蔗糖溶液,0.02%大蒜素-5%蔗糖溶液,3 d后在磷酸盐缓冲溶液中解剖肠道,用5 μmol/L浓度的细胞内ROS敏感荧光染料二氢乙啶(dihydroethidium, DHE)室温黑暗孵育30 min。然后用PBS洗涤肠道3次,以及用4%多聚甲醛固定组织30 min,再用PBS洗涤肠道3次。随后将肠道转移到玻片上,封片剂封片,在荧光显微镜采集图像并用ImageJ软件对荧光强度进行定量分析(Lietal., 2020)。实验重复4次
1.13 数据分析
使用Excel软件整合分析数据,每组数据得平均值(mean)和标准误(SE),用SPSS软件对各处理间的差异进行单样本t检验分析(one-samplet-test)。使用Graphpad和Adobe Illustrator软件制作相关图。
2 结果
2.1 黑腹果蝇对大蒜素的产卵避性反应
为探究某些辛辣植物对黑腹果蝇产卵选择行为的影响,首先选用大蒜作为刺激物。在食物中加入一定量大蒜汁,与对照组食物构成双向选择器,使用果蝇产卵双向选择器进行定量研究(图1: A)。其结果显示,黑腹果蝇成虫对大蒜汁有明显的产卵避性,仅仅将7.92%的卵产在含有大蒜汁的一侧,产卵指数(OI)为-0.84(图1: B),说明了大蒜汁中的某种物质导致果蝇产卵避性。进一步寻找大蒜汁中引起此行为的特异化合物,采用0.01%, 0.015%和0.02%大蒜素进行实验。其结果显示,OR果蝇对3个不同浓度的大蒜素均有明显产卵避性,且呈现一定的浓度梯度效应,即随着大蒜素浓度加大,果蝇产卵避性更加显著,OI分别为-0.53,-0.77和-0.88,与对照组相比均差异极显著(P<0.01)(图1: C)。另一野生品系CS黑腹果蝇对大蒜素有同样的避性反应,OI分别为-0.41,-0.66和-0.67,与对照组相比均差异极显著(P<0.01)(图1: D)。上述结果说明,大蒜素可作为大蒜主要排斥果蝇的产卵刺激物。为了增加实验的特异性,后续实验均使用大蒜素作为刺激物。各浓度大蒜素处理组与对照组相比,黑腹果蝇成虫单雌产卵量没有显著差异(P>0.05)(图1: E),说明大蒜素对果蝇产卵能力的影响可以忽略不计。

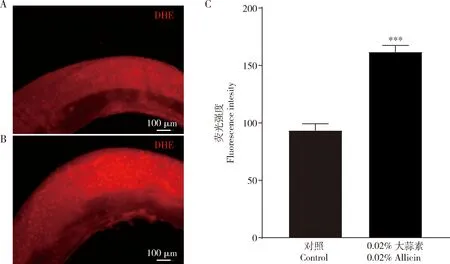
图1 黑腹果蝇雌成虫对大蒜素的产卵避性反应Fig. 1 Oviposition avoidance response of female adults of Drosophila melanogaster to allicinA: 产卵选择行为试验装置Oviposition selection behavior test device; B: 野生型黑腹果蝇OR在含大蒜的培养基上的产卵避性Oviposition avoidance of the wild-type OR strain of D. melanogaster on a medium containing garlic; C: 野生型黑腹果蝇OR对大蒜素的产卵避性 Oviposition avoidance of the wild-type OR strain of D. melanogaster to allicin; D: 野生型黑腹果蝇CS对大蒜素的产卵避性 Oviposition avoidance of the wild-type CS strain of D. melanogaster to allicin; E: 大蒜素对黑腹果蝇产卵的影响 Effects of allicin on the oviposition of D. melanogaster. 产卵指数(OI)=(实验组卵数-对照组卵数)/(实验组卵数+对照组卵数) Oviposition index (OI)=(number of eggs in the experimental group-number of eggs in the control group)/(number of eggs in the experimental group+number of eggs in the control group). 图中数据为平均值±标准误。柱上符号表示与对照组(H2O)间的差异显著性(nsP>0.05; *0.01 图2 大蒜素对黑腹果蝇成虫摄食(A)和位置效应(B)的影响Fig. 2 Effects of allicin on feeding (A) and location effect (B) of Drosophila melanogaster adults摄食指数(FI)=(含化学试剂的溶液消费量-仅含水的消费量)/总消费量Feeding index (FI)=(solution consumption containing chemical reagents-only water consumption)/total consumption; 位置指数(PI)=(实验组果蝇数目-对照组果蝇数目)/(实验组果蝇数目+对照组果蝇数目) Position index (PI)=(number of flies in the experimental group-number of flies in the control group)/(number of flies in the experimental group+number of flies in the control group). 使用不添加大蒜素的食物作对照 Foods without allicin were used as the control. 为了研究黑腹果蝇对大蒜素的摄食偏好性,利用毛细管摄食法进行检测(图2: A)。结果显示,黑腹果蝇对5%蔗糖有显著的摄食偏好,摄食指数(FI)为0.91,与已有报道(Prasad and Hens, 2018)一致; 而黑腹果蝇对0.01%和0.015%大蒜素的摄食呈现明显避性反应,FI分别为-0.76和-0.77,这与果蝇对大蒜素的产卵避性趋势一致。在一定时间内,产卵选择装置也能检测果蝇对大蒜素的位置偏嗜性(图2: B)。其结果显示,果蝇对0.01%和0.015%的大蒜素呈现极显著的位置避性(P<0.001),其位置指数(PI)分别为-0.09和-0.22,表明果蝇对大蒜素产生位置排斥反应。上述结果提示,摄食和位置可能会影响果蝇产卵选择性,但是产卵选择性是果蝇天生行为,具有其独立性,有别于摄食与位置行为。 目前大蒜素是通过何种感觉系统影响果蝇的产卵避性反应尚未可知,我们尝试从以下几个方面进行探究。利用暗盒检测视觉系统是否介导果蝇对大蒜素的产卵避性反应。结果发现,黑暗条件下,果蝇对大蒜素仍具有明显的产卵避性,0.01%, 0.015%和0.02%大蒜素处理组果蝇的产卵指数分别为-0.59,-0.64和-0.68,与对照组无显著差异(P>0.05)(图3: A),说明视觉不介导果蝇对大蒜素的产卵避性反应。 图3 嗅觉系统介导黑腹果蝇雌成虫对大蒜素产卵避性Fig. 3 The olfactory system mediates the oviposition avoidance of female adults of Drosophila melanogaster to allicinA: 黑暗条件不影响果蝇对大蒜素产卵避性Darkness has no effect on the oviposition avoidance in Drosophila to allicin; B: 摘除前足的果蝇对大蒜素的产卵避性仍存在The oviposition avoidance in Drosophila with the foreleg removed to allicin is still present; C: 果蝇对大蒜素有趋避性Drosophila is repellent to allicin; D: 摘除触角的果蝇对大蒜素产卵避性反应明显减弱 The oviposition avoidance response of Drosophila with antennae removed to allicin is obviously weakened; E: 突变体Orco2果蝇不影响其对大蒜素产卵避性The mutant Orco2of Drosophila does not affect its oviposition avoidance to allicin; F: 突变体TrpA1果蝇对大蒜素产卵避性显著减弱 The mutant TrpA1 of Drosophila has significantly impaired oviposition avoidance to allicin. 反应指数= (实验组果蝇数-对照组果蝇数)/(实验组果蝇数+对照组果蝇数) Response index= (number of flies in the experimental group-number of flies in the control group)/(number of flies in the experimental group + number of flies in the control group). 产卵指数(OI)=(实验组卵数-对照组卵数)/(实验组卵数+对照组卵数)Oviposition index (OI)=(number of eggs in the experimental group-number of eggs in the control group)/(number of eggs in the experimental group+number of eggs in the control group). 图中数据为平均值±标准误。柱上符号表示两组间的差异显著性(nsP>0.05; *0.01 果蝇的味觉主要存在于初级味觉结构上:唇瓣、前足、翼缘和产卵器等(Lingetal., 2014)。为了检测味觉系统是否在果蝇对大蒜素的产卵避性反应中起决定性作用,通常经外科手术切除初级味觉器官(第一对胸足跗节),使果蝇丧失部分味觉感知。其结果显示,除了波动大的低浓度大蒜素组,味觉系统的部分丧失并没有改变果蝇对大蒜素的产卵避性反应(P>0.05)(图3: B),表明味觉系统并没有在果蝇对大蒜素的产卵避性反应中起决定性的作用。 大蒜素还引起强烈的嗅觉刺激(Borlinghausetal., 2014)。为了验证此观点,使用Y迷宫实验检测大蒜素是否能引起果蝇的嗅觉刺激。其结果显示,果蝇对大蒜素具有明显的位置避性,反应指数为-0.67(图3: C)。果蝇的嗅觉系统主要位于触角,经外科手术切除嗅觉器官,可致果蝇丧失嗅觉感知。其结果显示,0.01%, 0.015%和0.02%大蒜素处理组果蝇的产卵指数分别为-0.01, -0.04和-0.11(图3: D),说明嗅觉系统的丧失导致了果蝇对大蒜素的产卵避性反应显著降低,由此可知,正是嗅觉系统介导了果蝇对大蒜素的产卵避性反应。为了进一步验证嗅觉系统介导的这种行为,使用嗅觉系统突变体Orco2进行实验。然而结果却出乎意料,突变体Orco2仍然对大蒜素具有显著的产卵避性反应(图3: E),且与野生型果蝇OR无显著差异(P>0.05)。可能原因是尽管突变体Orco2对大多数气味丧失感知的,但是仍然可以感受到大蒜素气味。 TRPA1是一种瞬时受体电位的非选择性阳离子通道受体,可作为环境和有害刺激的生物传感器。TrpA1突变体果蝇可丧失对刺激性物质的感知,因此选择TrpA1突变体果蝇进行产卵双选择实验。与OR果蝇的产卵选择行为相反,TrpA1突变体果蝇对0.01%, 0.015%和0.02%大蒜素避性减弱,OI分别为-0.09, -0.06和0.13(图3: F)。以上结果表明,大蒜素对机体具有刺激性和毒害性。 为探究在大蒜素影响下果蝇后代的发育和生长状况,将果蝇的卵转移到含不同浓度大蒜素的食物中,记录果蝇成蛹和成虫羽化时间。数据显示,随着大蒜素浓度增大,成蛹和成虫羽化时间逐渐延长,其中在0.02%浓度时,成蛹时间比对照组延长了1.85 d(图4: A),成虫羽化时间比对照组延长了1.88 d(图4: B)。其结果说明, 大蒜素会引起果蝇的发育生长迟缓。为进一步验证大蒜素对果蝇发育的毒害作用,分别计算了蛹和成虫的存活率。与发育历期结果一致,蛹和成虫的存活率也随着大蒜素浓度升高而下降,其中在0.02%浓度时,蛹存活率下降了26.01%,成虫存活率下降了31.28%,与对照组差异极显著(P<0.001)(图4: C, D)。这表明大蒜素会降低后代适合度,同时也解释了亲代避开大蒜素产卵行为的原因。 图4 大蒜素条件下黑腹果蝇后代的发育历期和存活率Fig. 4 Developmental duration and survival rate of offspring of Drosophila melanogaster exposed to allicinA, B: 大蒜素延缓果蝇的成蛹和羽化 Allicin retards the puparium formation and adult eclosion of Drosophila; C, D: 大蒜素降低后代的存活率 Allicin reduces the survival rate of offspring. 成蛹和成虫羽化时间均自初产卵开始计时。Time to puparium formation and time to adult eclosion were calculated from the beginning of spawning. 为检测大蒜素是否对成虫有危害作用,我们检测了在不同浓度大蒜素条件下雌雄黑腹果蝇成虫的寿命。在此实验中,将果蝇暴露于含有大蒜素溶液的小管中,每天记录存活的果蝇数。其结果显示,随着大蒜素浓度增大,果蝇雌雄成虫寿命均显著地缩短(P<0.05)(图5: A, B)。通过奔跑与攀爬检测果蝇的运动能力。结果发现,对照组(暴露于5%蔗糖溶液)果蝇奔跑速度为1.36 cm/s,经0.1%大蒜素处理的果蝇奔跑速度为1.01 cm/s(图5: C),稍低于对照组速度。同样地,经0.1%大蒜素处理的果蝇攀爬速度要显著慢于对照组果蝇(P<0.01)(图5: D)。总的来说,大蒜素能缩短亲代黑腹果蝇雌雄成虫寿命,并对其运动能力造成一定的损伤,可降低亲代的适合度。 图5 大蒜素对黑腹果蝇成虫寿命和健康的影响Fig. 5 Effects of allicin on longevity and health of Drosophila melanogaster adultsA, B: 大蒜素缩短雌雄果蝇成虫寿命Allicin shortens the life span of female and male adults of Drosophila; C, D: 大蒜素损害果蝇健康Allicin damages Drosophila health. 存活率=当天存活果蝇数目/当天果蝇总数Survival rate= number of flies alive that day/total number of flies that day; 奔跑、攀爬速度=路程/时间Running and climbing speed=distance/time. 以置于5%蔗糖溶液中的果蝇为对照。不同浓度(0.02%, 0.05%和0.1%)大蒜素处理后48 h测定奔跑速度和攀爬速度。Flies exposed to 5% sucrose solution were used as the control. The running speed and climbing speed were tested at 48 h after treatment with allicin of different concentrations (0.02%, 0.05% and 0.1%). 机体在遭受有害刺激时,会产生大量的活性氧(ROS),从而引起应激应答(原慧萍和杨泽, 2015)。果蝇在摄取刺激性大蒜素后,可能激活一系列的应激反应。为验证此推论,使用ROS荧光探针二氢乙啶(DHE)检测黑腹果蝇成虫肠道ROS含量,以评定对果蝇肠道的应激水平。结果显示,具有代表性的ROS荧光强度在0.02%大蒜素处理组比对照组(5%蔗糖溶液)更亮(图6: A, B)。统计结果显示,0.02%大蒜素处理组的ROS荧光强度为162,明显高于未处理果蝇的荧光强度93(图6: C),ROS水平增加。上述结果表明,大蒜素刺激黑腹果蝇肠道上皮细胞,激活氧化应激反应。 图6 大蒜素调节黑腹果蝇成虫肠道内ROS水平Fig. 6 Allicin regulates ROS level in the intestine of Drosophila melanogaster adultsA: 对照组,5%蔗糖溶液喂养3 d的果蝇成虫肠道Control group, intestine of Drosophila adults fed with 5% sucrose solution for 3 d; B: 含0.02%大蒜素的5%蔗糖溶液喂养3 d的果蝇成虫肠道Intestine of Drosophila adults fed with 5% sucrose solution containing 0.02% allicin for 3 d; C: 果蝇肠道细胞ROS荧光强度定量分析Quantitative analysis of ROS fluorescence intensity in Drosophila intestinal cells. 对不同溶液饲养3 d的果蝇成虫进行解剖,对肠道进行染色,用二氢乙啶测定活性氧水平。Drosophila adults fed with different solutions for 3 d were dissected, the intestine was stained, and the ROS level was measured with dihydroethidium. 对于植食性昆虫来说,果蝇以腐烂水果为食,会与寄主植物进行长久的斗争(Richmondetal., 1979)。因此,许多植物利用化学防御系统产生许多次级代谢产物。有研究结果表明,果蝇对多种有害物质均产生强烈排斥反应(Ebenetal., 2020)。众所周知,大蒜素现已被证实为大蒜主要的活性成分(张庭廷等, 2007)。大蒜素具有强烈的臭味,并且接触后会引起辛辣的痛觉(Macphersonetal., 2005)。果蝇产卵选择行为可以用来检测幼虫和成虫对刺激物的厌恶(Aietal., 2010; 王璐璐等, 2017)。本研究发现,当果蝇在添加大蒜汁的食物和正常的食物之间进行选择时,果蝇对大蒜汁有明显的产卵避性(图1: B),并且大蒜素浓度越高,其产卵避性越明显,且呈现一定的浓度梯度效应(图1: C, D)。以上结果说明果蝇对大蒜素的产卵避性不是由遗传因素差异引起的。值得注意的是,大蒜素还可以引起摄食和位置回避,且摄食和位置回避行为与果蝇的产卵避性反应存在一定的一致性,果蝇通过回避有浓度的大蒜素食物来降低对自身的危害性(图2)。但事实上,产卵偏好并不总是与位置偏好相对应,因为果蝇会对环境进行平衡预估,以确保其后代的生存。此外,在有或没有大蒜素的情况下,雌性的产卵量相当(图1: E)。这表明大蒜素对成年雌果蝇的产卵能力影响可以忽略。 动物具有趋利避害的本能,其中嗅觉发挥着重要的作用,可介导动物趋避行为(Macphersonetal., 2005)。果蝇的触角有嗅觉感受器(Heimbecketal., 1999)。经外科手术切除嗅觉器官,可致果蝇丧失嗅觉感知。本研究结果表明,嗅觉系统的丧失导致了黑腹果蝇对大蒜素的产卵避性反应明显减弱,由此可知,果蝇对大蒜素的产卵避性反应是由嗅觉系统介导的(图3: C)。TRPA1是一种瞬时受体电位的非选择性阳离子通道受体,可作为环境和有害刺激的生物传感器,因此选择TrpA1突变体果蝇进行产卵双选择实验。研究表明,其产卵行为与OR果蝇选择行为不同,并未出现明显的产卵偏嗜性 (图3: F)。TRPA1离子通道参与痛觉,同时也会被刺激性气味物质所激发(Latorreetal., 2007; Lietal., 2020)。研究表明人类TRPA1并不局限于对疼痛的感知,而是延伸到对气味的感知,TRPA1在小鼠嗅觉上皮和嗅球中的表达已被证实(Schützetal., 2014)。由此推测,TRPA1作为一种有害刺激的生物传感器,也可能会通过影响嗅觉进而影响果蝇的生物学行为。另外,避蚊胺(N,N-二乙基间甲酰胺)是世界上使用最广泛的外用驱蚊剂,避蚊胺抑制食物气味对果蝇的行为吸引,而这种抑制需要高度保守的嗅觉共受体OR83b(Ditzenetal., 2008)。由此可见,嗅觉系统在果蝇的趋避行为上起重要作用。 有研究表明大蒜精油对根蛆的存活率和繁殖率有很大的影响,特别是在其幼虫期(Zhuetal., 2017)。据果蝇生态行为和资源需求可知,幼虫移动能力有限,极易受到刺激物的攻击,为了增加幼虫的生存率,雌性成虫必须选择合适的产卵地点。研究发现,大蒜素会降低黑腹果蝇后代适合度,即大蒜素引起了果蝇后代发育与生长的迟缓,进一步验证了大蒜素对果蝇生长发育的毒性(图5: A, B)。这重要地解释了亲代避开大蒜素产卵行为 (图1: C) 的原因。果蝇摄取的大蒜素通过消化系统与肠道上皮细胞发生相应的反应,对消化系统造成一定程度的损害,导致肠道屏障功能的紊乱(图6)。肠道功能屏障是一个很好的预测指标,动物们为了适应环境,它们通常会调节其先天免疫来达到肠道内稳态(Biteauetal., 2008)。本研究发现大蒜素会影响中肠产生和清除ROS(图6),这可能对果蝇的健康及寿命造成不良后果。 综上所述,本研究首次发现大蒜素会引起果蝇显著的产卵避性反应并降低其后代的存活率。大蒜素可作为一种绿色驱避剂,达到生物防治和环境保护的目的。然而,大蒜素是否对其他昆虫同样也具有毒害作用仍未可知,有待进一步研究。在机制上,大蒜素是否影响果蝇肠道菌群平衡而影响后代的生长发育,或许能够进一步揭示果蝇分子病理学与生态行为之间的关系。
2.2 大蒜素对黑腹果蝇摄食和位置效应的影响
2.3 嗅觉系统介导黑腹果蝇雌成虫对大蒜素的产卵避性

2.4 TRPA1介导黑腹果蝇雌成虫对大蒜素的产卵避性
2.5 大蒜素降低后代适合度

2.6 大蒜素对亲代黑腹果蝇寿命和健康的影响

2.7 大蒜素激活黑腹果蝇氧化应激反应
3 讨论
猜你喜欢
学苑创造·A版(2022年3期)2022-03-29世界中医药(2020年8期)2020-10-21保健与生活(2019年22期)2019-11-25福建基础教育研究(2019年2期)2019-09-10学苑创造·A版(2019年6期)2019-07-11故事作文·高年级(2019年6期)2019-06-20福建基础教育研究(2019年2期)2019-05-28Coco薇(2016年8期)2016-10-09女士(2006年5期)2006-07-17
