
荷叶铁线蕨孢子体解剖结构和组织化学特征研究
2022-03-17 01:05:46李林宝黄桂云张国禹吴笛吴锦华梁前艳杨兰芳陈会员汪婷杨朝东
广西植物 2022年1期
李林宝 黄桂云 张国禹 吴笛 吴锦华 梁前艳 杨兰芳 陈会员 汪婷 杨朝东
摘 要: 荷葉铁线蕨为岩生珍稀蕨类植物,分布在中国重庆市万州、涪陵等极少地区。为揭示荷叶铁线蕨生长特性,采集栽培在种质资源圃中的荷叶铁线蕨根样、根状茎、在阳光下生长和在阴暗环境下生长的叶片,固定于甲醛-酒精-乙酸溶液中,用双面刀片进行徒手切片,分别用三种0.1%苏丹红、0.1%硫酸氢黄连素-苯胺兰、0.05%甲苯胺蓝染色剂染色,之后在莱卡光学显微镜、荧光显微镜下观察,研究荷叶铁线蕨适应生长的环境解剖学和组织化学特征。结果表明:(1)荷叶铁线蕨孢子体具初生结构,不定根内皮层具凯氏带和栓质化,厚壁组织层,皮层和表皮。(2)茎具网状中柱,内皮层具凯氏带且栓质化,厚壁组织层,皮层和薄的角质层。(3)叶具内皮层包围维管束,周缘厚壁组织层,等面叶,木质化表皮和薄角质层。(4)该植物的网状中柱、内皮层、厚壁组织层和木质化表皮等结构表明荷叶铁线蕨适应旱生环境,而其薄的角质层和等面叶则表明该植物适应阴生环境。因此,荷叶铁线蕨的解剖特征表明其适应阴生、干旱的岩生环境。
关键词: 内皮层, 网状中柱, 厚壁组织层, 栓质层, 薄角质层
中图分类号: Q944.5
文献标识码: A
文章编号: 1000-3142(2022)01-0090-10
收稿日期: 2021-01-28
基金项目: 中国长江三峡集团有限公司生态环保基金(WWKY-2020-0257)[Supported by the Ecological Environmental Protection Fund of China Three Gorges Corporation (WWKY-2020-0257)]。
第一作者: 李林宝(1991-),硕士,助理工程师,主要从事珍稀植物保护和分子遗传育种研究,(E-mail)1239393556@qq.com。
*通信作者: 杨朝东,博士,硕士生导师,研究方向为湿地植物生理生态,(E-mail)546728708@qq.com。
Anatomical structure and histochemical features of Adiantum
reniforme var. sinense sporophyte (Pteridaceae)
LI Linbao1, HUANG Guiyun1, ZHANG Guoyu1,WU Di1, WU Jinhua1, LIANG Qianyan1,
YANG Lanfang1, CHEN Huiyuan1, WANG Ting2, YANG Chaodong2*
( 1. China Three Gorges Corporation Yangtze River Rare Plant Research Institue,Yichang 44300, Hubei, China;
2. Yangtze University College of Horticulture and Gardening, Jingzhou 434025, Hubei, China )
Abstract: The distribution of the rare perennial fern Adiantum reniforme var. sinense (Pteridaceae) which is endemic to shady cliff environments is limited to small areas of Wanzhou and Fuling, Chongqing, China. In order to reveal the growth characteristics of A. reniforme var. sinense in the germplasm resource nursery,we observed the tissue structure of A. reniforme var. sinense, which including in the roots, rhizomes, leaves grown in the sun and in shade. First of all, the roots, rhizomes, leaves were collected and fixed in formaldehyde-alcohol-acetic acid solution, and we cut the tissue structure of the plant with a double-sided blade to make freehand sections, then the tissue were stained with three kinds of 0.1% Sudan red, 0.1% berberine bisulfate-aniline blue, and 0.05% toluidine blue, at last plant tissue sections were observed under the Lycra optical microscope and the fluorescence microscope. In this study, we used brightfield and epifluorescence microscopy to investigate the anatomical structures and histochemical features that may allow this species to thrive in shady, dry cliff environments. The results were as follows: (1) The A. reniforme var. sinense sporophyte had a primary structure, the roots had an endodermis with Casparian walls, suberin lamellae, a thickened sclerenchyma layer around the endodermis, a cortex, and a rhizodermis. (2) The stems had a dictyostele, an endodermis with Casparian walls and suberin lamellae, a thickened sclerenchyma layer around the endodermis, a cortex, and a thin cuticle. (3) The leaves had an endodermis surrounded the vascular bundles, a peripheral sclerenchyma layer, an isolateral mesophyll, a lignified epidermis and a thin cuticle. (4) The dictyostele, endodermis, sclerenchyma layers, and lignified epidermal walls reflected the drought tolerance of A. reniforme var. sinense, while its thin cuticle and isolateral mesophyll suggested tolerance of shady environments. Thus, according to the anatomical characteristics of A. reniforme var. sinense, it is concluded that A. reniforme var. sinense are consistent with adaptations to shady, dry cliff environments.
Key words: endodermis, dictyostele, sclerenchyma layer, suberin lamellae, thin cuticle
荷叶铁线蕨(Adiantum reniforme var. sinense)是一种自然分布在中国重庆市万州区、涪陵、石柱县的覆有薄土阴暗的岩石上及石缝中的多年生蕨类植物,作为中药材已有100多年历史(Lin, 1980;张泽宏和吴小霞,2013;Rothfels & Schuettpelz, 2014;PPG, 2016;Pryer et al., 2016)。荷叶铁线蕨与其近缘物种具有洲际间断分布特点,A. reniforme分布于亚速尔群岛,A. reniforme var. asarifolium分布于非洲中南部(Lin, 1980;张泽宏和吴小霞,2013;Wang et al., 2015)。为了更好地保护荷叶铁线蕨,自2012年中国三峡大坝完工以后,荷叶铁线蕨已被迁地保护在中国长江三峡集团有限公司和中国科学院武汉植物园的种质资源苗圃中(周赛霞等,2008;吴金清,2012)。由于荷叶铁线蕨濒临灭绝,分布范围狭窄,遗传多样性低,因此被列为国家一级保护蕨类植物(Lin, 1980;徐天全等,1987;Fu & Jin, 1992;Liu et al., 2007;Zhang et al., 2013;Wang et al., 2015)。
蕨类植物已进化出各种形态解剖学结构来适应陆生、旱生、附生、岩生和水生环境(Wylie, 1949;Zhang et al., 2013;Neira et al., 2017;Srbu et al., 2017;Wetzel et al., 2017)。蕨类植物根状茎常具有网状中柱;不定根具初生木质部和初生韧皮部,皮层具内皮层和外皮层或者皮下层,或者中间皮层木质化;叶具内皮层、厚壁组织层和角质层(Jeffreys, 1903;Wardlaw, 1944;Chapple & Peterson, 1987;Damus et al., 1997;Schneider, 1997;Ribeiro et al., 2007;De los ngeles Lagoria et al., 2018;Mahley et al., 2018;Palacios-Rios et al., 2019)。陽生叶和阴生叶的形态特征明显不同,阴生植物的叶片大、细胞大、叶肉组织少、气孔和叶脉密度少(Boardman, 1977;Ribeiro et al., 2007;张泽宏和吴小霞,2013;Neira et al., 2017;Dematteis et al., 2019;Shah et al., 2019;Baer et al., 2020)。
铁线蕨属植物(Adiantum)的系统发育和遗传亲缘关系是以解剖学特征为基础,特别是叶片中的网状中柱(Wylie, 1949;Imaichi, 1988;Huiet et al., 2018)。然而,近年来的分子遗传学分析认为,荷叶铁线蕨与A. nelumboides相似程度高而合并为同一个物种,经典形态分类学则认为是两个物种(Lin, 1980;Zhang et al., 2013;Wang et al., 2015)。因此,荷叶铁线蕨在铁线蕨属的分类学、进化学和系统发育多样性地位等方面还需要进一步探讨(Kato & Imaichi, 1997;Li et al., 2013;Vasco et al., 2013;Plackett et al., 2015;Harrison & Morris, 2017;Huiet et al., 2018)。目前对荷叶铁线蕨孢子体解剖结构特征研究很少(Wu et al., 2011)。
本文主要研究荷叶铁线蕨孢子体的解剖结构和组织化学特征,以确定哪些结构特征使其能适应于阴暗干旱的岩生环境。实验采集了荷叶铁线蕨孢子体的阳生叶和阴生叶,研究其适应阴生环境的形态学和解剖学特征,其叶片是否表现出与其他阴生植物相同的特征。这些适应性特征有助于解释荷叶铁线蕨对阴暗、干旱岩生环境的适应能力。本研究结果也为今后荷叶铁线蕨及其近缘种的迁地保护、分类学、进化和系统发育等方面的研究提供参考依据。
1 材料与方法
1.1 植物材料的收集与处理
因为三峡库区蓄水淹没了荷叶铁线蕨原有生态环境,所以现有它的种质资源都为迁地保护群落。本研究于2019年10月在湖北宜昌中国三峡集团公司种质资源苗圃,采集了该物种的孢子体材料。从5棵植物中收集了10个根样品(长度为70~130 mm),8个根状茎样品(长度为40~60 mm)。从10棵植株中分别采集5片在强光下生长的成年叶片(光照强度约为4 840 lx,湿度为51.6%)和5片在阴暗环境下生长的成年叶片(光照强度约为805.5 lx;湿度为49.5%)。将采集的新鲜样品立即固定于甲醛-酒精-乙酸(FAA)溶液中(Ruzin, 1999)。
1.2 解剖结构和组织化学特征
在立体解剖镜下(JNOEC JSZ6,中国),用双面刀片在距根尖5、10、20 mm部位进行徒手切片。根状茎切片位置:幼嫩部分(距茎尖约10 mm,表面呈白色)和成熟部分(距茎尖约30 mm,表面呈棕色)。叶柄切片位置:幼嫩部分(表面白色)和成熟部分(表面黑色)。在健康叶片中心位置进行切片以确定组织厚度及其各结构。将切片分为三组,每组包含3~6个切片(从不同的样本中获得),分别代表距离根尖不同位置特征,幼嫩和成熟的根状茎、叶柄不同位置特征,叶片的中心和边缘不同位置特征。
每组切片分别用以下三种染色剂染色:0.1%苏丹红7B (SR7B)用于检测栓质化细胞壁(Brundrett et al., 1991),0.1%(m/V)硫酸氢黄连素-苯胺兰(BAB)对染切片检验凯氏带和木质化细胞壁(Brundrett et al., 1988;Seago et al., 1999),0.05%甲苯胺兰(TBO)用于更清晰地观察组织器官的结构(Feder & O’Brien, 1968;Peterson et al., 2008)。所有样本染色后均用无菌水清洗2~3次,并滴加无菌水盖片,在莱卡光学显微镜(Leica DME,德国)下观察,用数码相机和测微尺(Nikon E5400,日本)拍照记录。硫酸氢黄连素-苯胺兰染色的切片在荧光显微镜(Olympus IX71)紫外光下用激发滤光片G 365 nm、吸收滤光片U-WB(蓝光)、双色镜DM 500、补偿激发滤光片BP 450-480、补偿吸收滤光片BA 515观察,并用数码相机和测微尺(RZ200C-21,中国)拍照记录(Yang et al., 2011)。
2 结果与分析
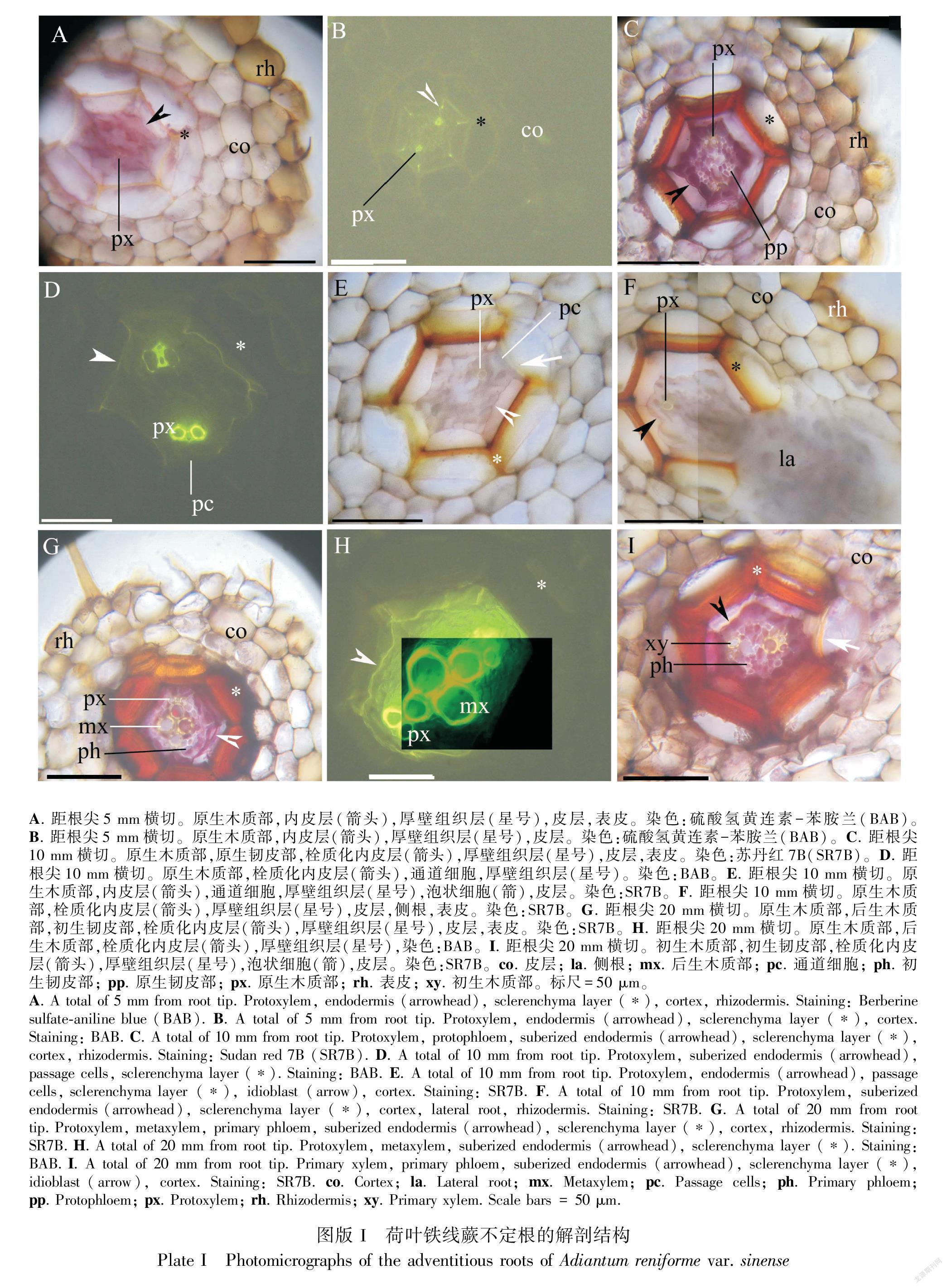
2.1 不定根的解剖结构
不定根中柱为二原对称型并具原生木质部(图版I:A-I)。在距根尖5 mm处,不定根内皮层中有微弱的凯氏带,其周围有一层细胞的厚壁组织层,根表面有表皮(图版I:A,B)。在距根尖10 mm处,侧根从中柱鞘伸出,此时中柱具有显著的原生木质部和原生韧皮部(图版I:C-F)。此外,内皮层具完整的栓质层细胞壁和少数通道细胞,与通道细胞和原生木质部不同的是,厚壁组织层具较厚的细胞壁(图版I:C-F)。在距根基部20 mm处,中柱具完整初生木质部和初生韧皮部组织,内皮层具较厚的栓质化细胞壁,厚壁组织层进一步增厚(图版I:G-Ⅰ)。
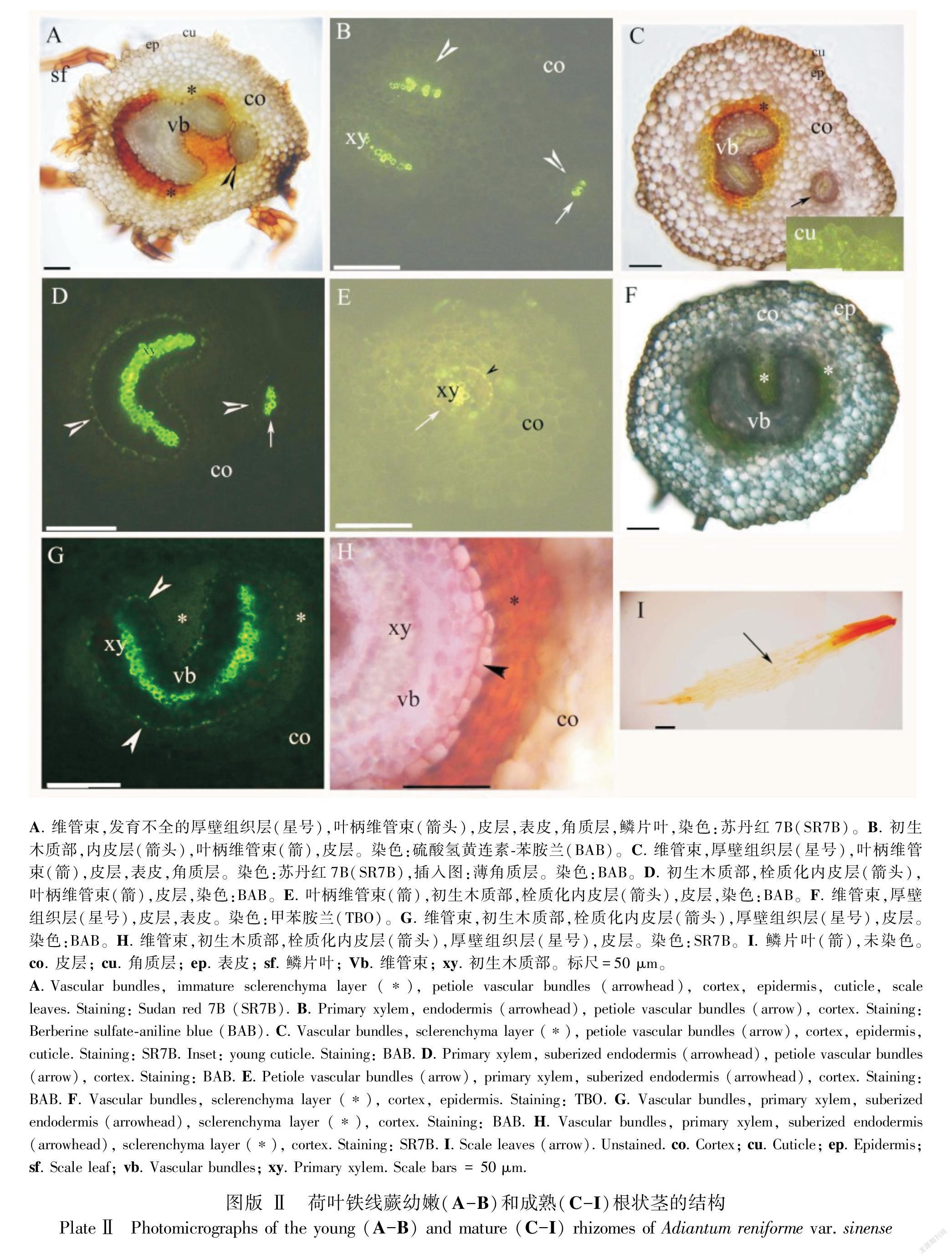
2.2 根状茎的解剖结构
幼嫩和成熟的根状茎中都有被厚壁组织层包围着的网状中柱。网状中柱包含叶柄维管束,其中部有显著的原生木质部(图版Ⅱ:A-H)。每个维管束中都有内皮层并具凯氏带(图版Ⅱ:B,D,E,G),并在成熟时栓质化(图版Ⅱ:D,E,G,H)。根状茎皮层为薄壁组织,其表面具薄的角质层和褐色鳞叶(图版Ⅱ:A,C,F,I)。叶柄维管束来源于根状茎(图版Ⅱ:A-E)。
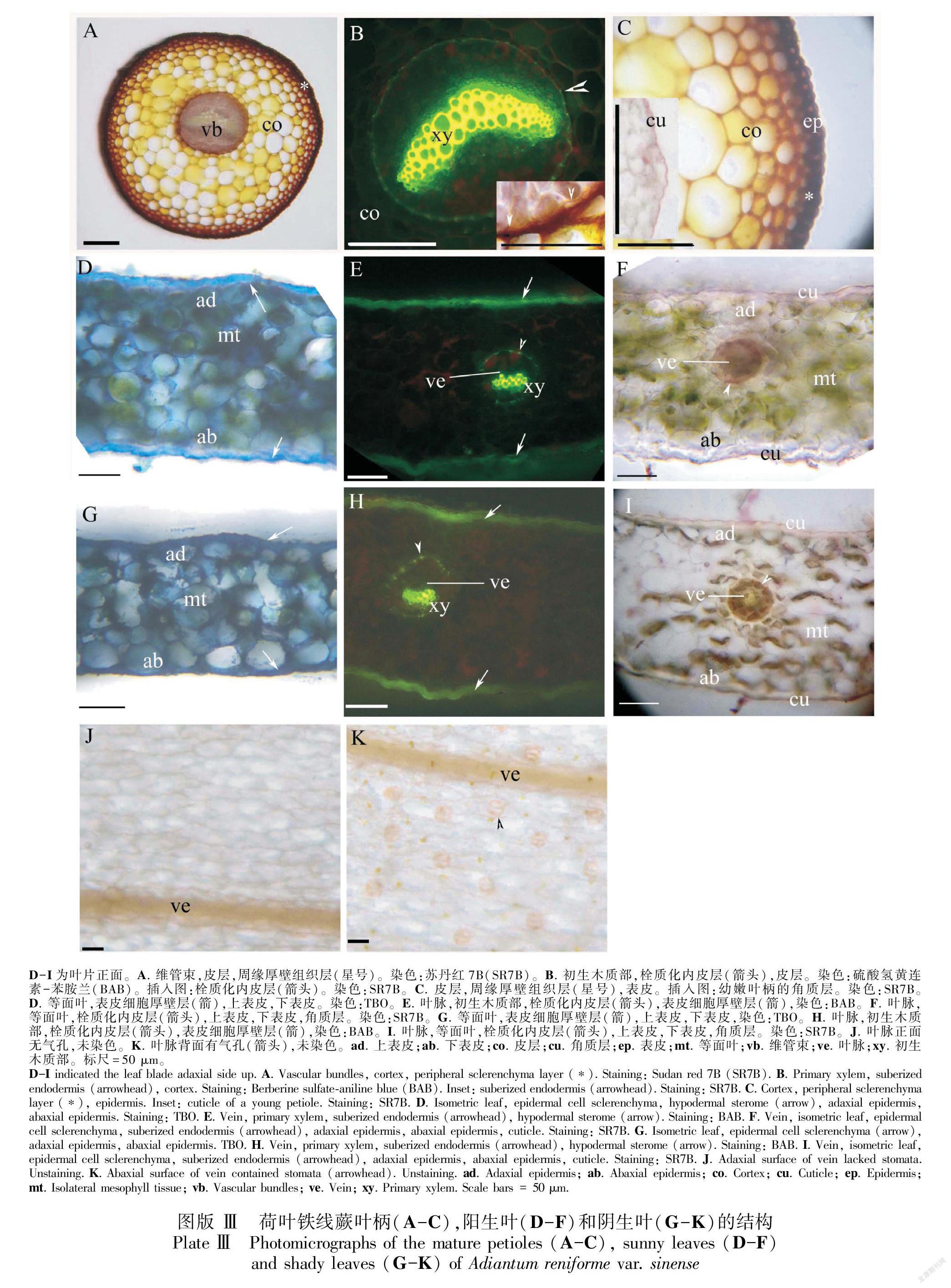
2.3 不同光强下叶的解剖结构
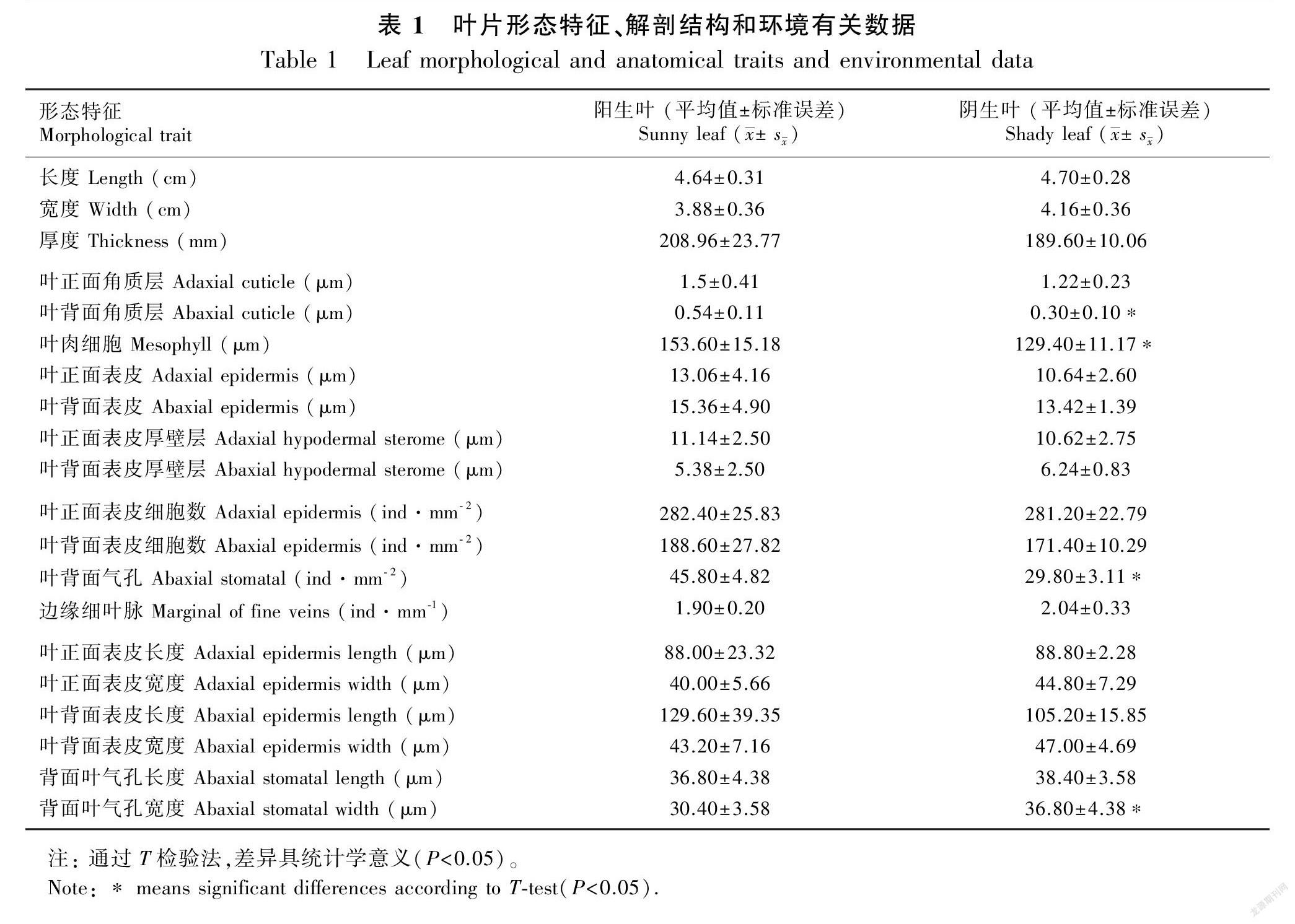
幼嫩和成熟的叶柄中有单个维管束,中央具内皮层、皮层、周缘厚壁组织层和表皮(图版Ⅲ:A-C)。成熟时,内皮层具凯氏带且栓质化(图版Ⅲ:B)。叶柄表面具薄角质层(图版Ⅲ:C)。荷叶铁线蕨的叶片面积、组织厚度、组织密度和表皮等主要特征见表1。阳生和阴生环境中的叶片均具表皮,等面叶和二叉叶脉具厚壁组织层(图版Ⅲ:D-I,表1);二叉叶脉的内皮层具凯氏带且栓质化(图版Ⅲ:E, F, H, I)。气孔仅存在于下表皮(图版Ⅲ:J, K);与阳生叶相比,阴生叶的气孔更少且更大(表1)。与阴生叶相比,阳生叶的正面和背面角质层相对较厚(图版Ⅲ:D-I,表1)。叶片表皮细胞具厚壁层(图版Ⅲ:D-I)。与阴生叶相比,阳生叶片的中部叶肉更厚(图版Ⅲ:D-I,表1)。
3 討论与结论
本文研究了荷叶铁线蕨孢子体不同部位的形态特征和解剖结构,这些特征有利于其在干旱、阴暗的环境中成功繁衍。荷叶铁线蕨的不定根、根状茎和叶中都具有内皮层,并具凯氏带且栓质化,周围有厚壁组织层。这些特征与其他维管植物的组织和器官存在较多的共同之处(Ogura, 1972;Fahn, 1990;Lersten, 1997;Enstone et al., 2003;Evert, 2006;Crang et al., 2018)。陆生植物Asplenium sp.和附生植物Pleopeltis macrocarpa与荷叶铁线蕨相似,在整个植物的维管束周围分布着内皮层和厚壁组织层(Wetzel et al., 2017;De los ngeles Lagoria et al., 2018)。蕨类植物内皮层和厚壁组织层支撑并保护维管植物,这些结构表明了荷叶铁线蕨能在干旱的岩生环境中生存(Chapple & Peterson, 1987;Fahn, 1990;Lersten, 1997;Enstone et al., 2003;Evert, 2006;Neira et al., 2017;Wetzel et al., 2017;Crang et al., 2018;De los ngeles Lagoria et al., 2018)。
许多蕨类植物的不定根中具有内皮层和厚壁组织层,其中包括陆生植物鹿角蕨(Platycerium bifurcatum)(Chapple & Peterson, 1987)、Asplenium sp. (Schneider, 1997;Leroux et al., 2011;Wetzel et al., 2017)、玉柏石松(Lycopodium obscururn)、Selaginella sp.(Damus et al., 1997)、蜈蚣草(Pteris vittata)(Bondada et al., 2006;Sridhar et al., 2011)、Pleopeltis sp. (Hernández et al., 2013;De los ngeles Lagoria et al., 2018)和Doryopteris triphylla (Neira et al., 2017)。这些植物的根与荷叶铁线蕨的根也有其他相似之处,如鹿角蕨的内皮层栓质化(Chapple & Peterson, 1987);在鹿角蕨和Pleopeltis macrocarpa的厚壁组织层中存在具薄细胞壁的泡状细胞(Chapple & Peterson, 1987;De los ngeles Lagoria et al., 2018);Selaginella sp.的根具内皮层和木质化皮层细胞壁(Damus et al., 1997);鹿角蕨、Pleopeltis sp.和Doryopteris triphylla的根中具2层或多层细胞的厚壁层(Chapple & Peterson, 1987;Hernández et al., 2013;Neira et al.,2017;De los ngeles Lagoria et al., 2018);Asplenium sp.的根表皮细胞壁具螺旋状增厚(Wetzel et al., 2017)。与这些旱生植物相比,荷叶铁线蕨的根对干旱环境的适应性较弱(Chapple & Peterson, 1987;Damus et al., 1997;Hernández et al., 2013;Neira et al., 2017;Wetzel et al., 2017;De los ngeles Lagoria et al., 2018)。
荷叶铁线蕨根状茎具网状中柱、内皮层、厚壁组织层和薄角质层,这些结构特征与其他陆生蕨类植物相似,例如球子蕨(Onoclea sensibilis)(Wardlaw, 1944)、Polypodium sp. (Srivastava & Chandra, 2009)、蜈蚣草(Sridhar et al., 2011)、Ceradenia sp. (Deroin & Rakotondrainibe, 2015)、Blotiella lindeniana (Becari-Viana & Schwartsburd, 2017)和Doryopteris triphylla (Neira et al., 2017)。附生植物和旱生植物如Asplenium sp、Pleopeltis macrocarpa和贴生石韦(Niphobolus adnascens),这些植物根状茎的表皮下均有周缘机械厚壁组织层。因此,这些植物比阴生的荷叶铁线蕨更适应旱生环境(Pande, 1935;Wetzel et al., 2017;De los ngeles Lagoria et al., 2018)。
荷葉铁线蕨的叶柄表皮下有内皮层和周缘厚壁层,这与其他蕨类植物相似(Hernandez-Hernandez et al., 2012),如Anemia (Ribeiro et al., 2007)、Pteris (Bondada et al., 2006;Sridhar et al., 2011;Martínez & Vilte, 2012;Palacios-Rios et al., 2019)、Davallia (Ummu-Hani et al., 2013)、Blechnum (Noraini et al., 2014)、Asplenium (Wetzel et al., 2017)、Doryopteris triphylla (Neira et al., 2017)和Pleopeltis macrocarpa (De los ngeles Lagoria et al., 2018)。在Asplenium植物中,叶柄内皮层有一层厚壁组织层包围(Wetzel et al., 2017),而在Doryopteris triphylla中,叶柄表皮层具厚角质层(Neira et al., 2017)。
荷叶铁线蕨的叶具栓质化内皮层,叶脉周围具凯氏带和表皮细胞具厚壁层(Mahley et al., 2018)。这些结构与Anemia sp.相似(Ribeiro et al., 2007;Mahley et al., 2018),表明荷叶铁线蕨适应于陆生环境。荷叶铁线蕨具等面叶,这在黄连(Coptis chinensis)、Doryopteris pentagona、海金沙(Lygodium japonicum)、井栏边草(Pteris multifida)、肾蕨(Nephrolepis cordifolia)、Adiantum capillus-veneris和白羽凤尾蕨(Pteris ensiformis cv. victoriae)中也能观察到(袁王俊等,2007;张泽宏和吴小霞,2013;Dematteis et al., 2019)。荷叶铁线蕨叶片的角质层薄,这类似于其他阴生蕨类植物,如海金沙、井栏边草、肾蕨、Adiantum capillus-veneris、白羽凤尾蕨和Doryopteris pentagona (张泽宏和吴小霞,2013;Dematteis et al., 2019)。与具厚角质层的阳生或旱生蕨类植物Cheilanthes glauca 和Doryopteris triphylla相比,具薄角质层的荷叶铁线蕨适应于阴生环境(张泽宏和吴小霞,2013;Neira et al., 2017;Dematteis et al., 2019;Shah et al., 2019)。在荷叶铁线蕨中,与阳生叶片相比,阴生叶具更薄的等面叶和较低的气孔密度,表明这种植物在不同环境条件下可塑性和适应不同环境条件的能力(Ribeiro et al., 2007;Santos-Silva et al., 2011;Neira et al., 2017;Dematteis et al., 2019;Shah et al., 2019;Baer et al., 2020)。
荷叶铁线蕨的不定根、叶柄和根状茎具有适应干旱环境的结构,其中内皮层、厚壁机械组织层和表皮细胞厚壁层是适应干旱环境的重要结构特征(Chapple & Peterson, 1987;Neira et al., 2017;Wetzel et al., 2017;De los ngeles Lagoria et al., 2018)。然而,像许多适应阴生环境的蕨类植物一样(Evert, 2006;袁王俊等,2007;张泽宏和吴小霞,2013;Crang et al., 2018),荷叶铁线蕨也具有薄的角质层和等面叶,这表明了对阴生环境的适应性。相反,附生和旱生蕨类植物更适应于干旱环境,它们的根具外皮层,有多层细胞的厚壁组织层,且表皮细胞壁具螺旋增厚(Chapple & Peterson, 1987;Damus et al., 1997;Neira et al., 2017;Wetzel et al., 2017;De los ngeles Lagoria et al., 2018);根状茎具周缘厚壁层(Wetzel et al., 2017;De los ngeles Lagoria et al., 2018);叶具厚角质层(Neira et al., 2017)。因此,荷叶铁线蕨的解剖结构和组织化学特征表明该植物适应于干旱、阴暗的岩生环境。
參考文献:
BAER A, WHEELER JK, PITTERMANN J, 2020. Limited hydraulic adjustments drive the acclimation response of Pteridium aquilinum (L.) Kuhn to variable light [J]. Ann Bot, 125(4): 691-700.
BECARI-VIANA I, SCHWARTSBURD PB, 2017. Morpho-anatomical studies and evolutionary interpretations of the rhizomes of extant dennstaedtiaceae [J]. Am Fern J, 107(3): 105-123.
BOARDMAN NK, 1977. Comparative photosynthesis of sun and shade plant [J]. Ann Rev Plant Phys, 28(1): 355-357.
BONDADA B, TU C, MA L, 2006. Surface structure and anatomical aspects of Chinese brake fern (Pteris vittata; Pteridaceae) [J]. Brittonia, 58(3): 217-228.
BRUNDRETT MC, ENSTONE DE, PETERSON CA, 1988. A berberine-aniline blue fluorescent staining procedure for suberin, lignin and callose in plant tissue [J]. Protoplasma, 146(2-3): 133-142.
BRUNDRETT MC, KENDRICK B, PETERSON CA, 1991. Efficient lipid staining in plant material with Sudan red 7B or Fluorol yellow 088 in polyethlene glycol-glycerol [J]. Biot Hist, 66(3): 111-116.
CHAPPLE CCS, PETERSON RL, 1987. Root structure in the fern Platycerium bifurcatum (Cav. ) C. Chr. (Polypodiaceae) [J]. Bot Gaz, 148(2): 180-187.
CRANG R, LYONS-SOBASKI S, WISE R, 2018. Plantanatomy [M]. Gewerbestrasse, Switzerland: Springer International Publishing.
DAMUS M, PETERSON RL, ENSTONE DE, et al., 1997. Modifications of cortical cell walls in roots of seedless vascular plants [J]. Bot Acta, 110(2): 190-195.
DE LOS NGELES LAGORIA M, AVILA G, NEIRA DA, et al., 2018. Morphoanatomical and histochemical characteristics of the epiphytic fern Pleopeltis macrocarpa (Polypodiaceae) [J]. Braz J Bot, 41(3): 739-750.
DEMATTEIS B, SOLS SM, YESILYURT JC, et al., 2019. Comparative anatomy in four Cheilanthoid ferns [J]. Bol de la Soc Arg Bot, 54(2): 203-214.
DEROIN T, RAKOTONDRAINIBE F, 2015. Comparative rhizome anatomy of some species of Ceradenia LE Bishop and Zygophlebia LE Bishop (Polypodiaceae, formerly Grammitidaceae) from Madagascar [J]. Mod Phyt, 7: 5-12.
ENSTONE DE, PETERSON CA, MA F, 2003. Root endodermis and exodermis: structure, function, and responses to the environment [J]. J Plant Growth Reg, 21(4): 335-351.
EVERT RF, 2006. Esau’s plant anatomy: meristems, cells, and tissues of the plant body [M]. Hoboken, New Jersey, USA: Wiley Interscience, John Wiley & Sons.
FAHN A, 1990. Plant Anatomy [M]. 4th ed. Oxford, UK: Pergamon Press.
FEDER N, O’BRIEN TP, 1968. Plant microtechnique: some principles and new methods [J]. Am J Bot, 55(1): 123-142.
FU LK, JIN JM, 1992. China plant red data book-rare and endangered plant [M]. Beijing: Science Press.
HARRISON CJ, MORRIS JL, 2017. The origin and early evolution of vascular plant shoots and leaves [J]. Phil Trans R Soc B: Biol Sci, 373(1739): 20160496.
HERN NDEZ MA, TERAN L, MATA M, et al., 2013. Helical cell wall thickenings in root cortical cells of Polypodiaceae species from Northwestern Argentina [J]. Am Fern J, 103(4): 225-240.
HERNANDEZ-HERNANDEZ V, TERRAZAS T, MEHLTRETER K, et al., 2012. Studies of petiolar anatomy in ferns: structural diversity and systematic significance of the circumendodermal band [J]. Bot J Linn Soc, 169(4): 596-610.
HUIET L, LI FW, KAO TT, et al., 2018. A worldwide phylogeny of Adiantum (Pteridaceae) reveals remarkable convergent evolution in leaf blade architecture [J]. Taxon, 67(3): 488-502.
IMAICHI R, 1988. Developmental anatomy of the shoot apex of leptosporangiate ferns. II. Leaf ontogeny of Adiantum capillus-veneris (Adiantaceae) [J]. CAN J Bot, 66(9): 1729-1733.
JEFFREYS EC, 1903. The structure and development of stem in pteridophyta and gymnosperms [J]. Phil Trans Royal Soc B Biol Sci, 195(60): 119-146.
KATO M, IMAICHI R, 1997. Morphological diversity and evolution of vegetative organs in pteridophytes [M]//Evolution and diversification of land plants. Tokyo: Springer, 1997: 27-43.
LEROUX O, BAGNIEWSKA-ZADWORNA A, RAMBE SK, et al., 2011. Non-lignified helical cell wall thickenings in root cortical cells of Aspleniaceae (Polypodiales): histology and taxonomical significance [J]. Ann Bot, 107(2): 195-207.
LERSTEN NR, 1997. Occurrence of endodermis with a casparian strip in stem and leaf [J]. Bot Rev, 63(3): 265-272.
LI X, FANG YH, YANG J, et al., 2013. Overview of the morphology, anatomy, and ontogeny of Adiantum capillus-veneris: an experimental system to study the development to ferns [J]. J Syst Evol, 51(5): 499-510.
LIN YX, 1980. New taxa of Adiantum L. in China [J]. Acta Phyt Sin, 18: 102.
LIU XQ, GITURU RW, CHEN LQ, 2007. Genetic variation in the endangered fern Adiantum reinforme var. sinense (Adiantaceae) in China [J]. Ann Bot Fenn, 44(1): 25-32.
MAHLEY JN, PITTERMANN J, ROWE N, et al., 2018. Geometry, allometry and biomechanics of fern leaf petioles: their significance for the evolution of functional and ecological diversity within the Pteridaceae [J]. Front Plant Sci, 9: 197.
MARTNEZ OG, VILTE I, 2012. The structure of petioles in Pteris (Pteridaceae) [J]. Am Fern J, 102(1): 1-11.
NEIRA DA, ANDRADA AR, P EZ VDL , et al., 2017. Anatomical, histochemical and cytogenetic features of Doryopteris triphylla (Pteridaceae) [J]. Am J Plant Sci, 8: 907-920.
NORAINI T, AMIRUL-AIMAN AJ, JAMAN R, et al., 2014. Systematic significance of stipe anatomy in peninsular Malaysian Blechnum L. (blechnaceae) species [J]. Mal Appl Biol J, 43(2): 119-125.
OGURA Y, 1972. Comparative anatomy of vegetative organs of the Pteridophytes [M]. Berlin, Germany: Gebruder Borntraeger, pp. 1-39.
PALACIOS-RIOS M, GALAN JMGY, PRADA C, et al., 2019. Structure of the petioles and costae of Mexican and Central American species of Pteris (Polypodiopsida, Pteridaceae) [J]. Phytotaxa, 401(2): 101-116.
PANDE SK, 1935. Notes on the anatomy of a xerophytic fern Niphobolus adnascens from the Malay Peninsula [J]. Proc Ind Acad Sci-Sect A Part 3 Mathemat Sci, 1(9): 556-564.
PETERSON RL, PETERSON CA, MELVILLE LH, 2008. Teaching plant anatomy through creative laboratory exercise [M]. Ottawa Ontartio: N.P C. Press.
PLACKETT ARG, DI STILIO VS, LANGDALE JA, 2015. Ferns: the missing link in shoot evolution and development [J]. Front Plant Sci, 6: 972.
PPG I, 2016. A community-derived classification for extant lycophytes and ferns [J]. J Syst Evol, 54: 563-603.
PRYER KM, HUIET L, LI FWEI, et al., 2016. Maidenhair ferns, Adiantum, are indeed monophyletic and sister to shoestring ferns, Vittarioids (Pteridaceae) [J]. Syst Bot, 41(1): 17-23.
RIBEIRO MLRC, SANTOS MG, MORAES MG, 2007. Leaf anatomy of two Anemia Sw. species(Schizaeaceae-Pteridophyte) from a rocky outcrop in Niterói, Rio de Janeiro, Brazil [J]. Braz J Bot, 30(4): 695-702.
ROTHFELS CJ, SCHUETTPELZ E, 2014. Accelerated rate of molecular evolution for vittarioid ferns is strong and not driven by selection [J]. Syst Biol, 63: 31-54.
RUZIN SE, 1999. Plant microtechnique and microscopy [M]. New York: Oxford University Press.
SANTOS-SILVA F, MASTROBERTI AA, DE ARAUJO MARIATH JE, 2011. Development of the epidermal cells of the pinnules of Adiantum raddianum C. Presl (Pteridaceae): environmental adaptive plastidial characteristics [J]. Am Fern J, 101(3): 172-181.
SRBU A, SMARANDACHE D, MARINESCU AT, et al., 2017. Anatomical-histological observations conducted on aquatic ferns In the Danube Delta [J]. J Plant Dev, 24: 3-21.
SCHNEIDER H, 1997. Root anatomy of Aspleniaceae and the implications for systematics of the fern family [J]. Fern Gaz, 15: 160-168.
SEAGO JR JL, PETERSON CA, ENSTONE DE, et al., 1999. Development of the endodermis and hypodermis of Typha glauca Godr. and T. angust ifolia L. roots [J]. CAN J Bot, 77(1): 122-134.
SHAH SN, AHMAD M, ZAFAR M, et al., 2019. Leaf micromorphological adaptations of resurrection ferns in Northern Pakistan [J]. Flora, 255(6): 1-10.
SRIDHAR BBM, HAN FX, DIEHL SV, et al., 2011. Effect of phytoaccumulation of arsenic and chromium on structural and ultrastructural changes of brake fern (Pteris vittata) [J]. Braz J Plant Phys, 23(4): 285-293.
SRIVASTAVA A, CHANDRA S, 2009. Structure and organization of the rhizome vascular system of four Polypodium species [J]. Am Fern J, 99(3): 182-193.
UMMU-HANI B, NORAINI T, MAIDEEN H, et al., 2013. Taxonomic value of the stipe anatomy in Davallia (Davalliaceae) in Peninsular Malaysia [J]. Mal Nat J, 65(2-3): 130-144.
VASCO A, MORAN RC, AMBROSE BA, 2013. The evolution, morphology, and development of fern leaves [J]. Front Plant Sci, 4: 345.
WANG AH, SUN Y, SCHNEIDER H, et al., 2015. Identification of the relationship between Chinese Adiantum reniforme var. sinense and Canary Adiantum reniforme [J]. BMC Plant Biol, 15(1): 36.
WARDLAW CW, 1944. Experimental and analytical studies of pteridophytes: IV. Stelar morphology: Experimental observations on the relation between leaf development and stelar morphology in species of Dryopteris and Onoclea [J]. Ann Bot, 8(32): 387-399.
WETZEL MLR, SYLVESTRE LDS, BARROS CF, et al., 2017. Vegetative anatomy of Aspleniaceae newman from Brazilian Atlantic rainforest and its application in taxonomy [J]. Flora, 233: 118-126.
WU JQ, 2012. Key rare and endangered species in Three Gorges Reservoir Area [J]. Chin Three Gorg Constr, 5: 26-34. [吳金清, 2012. 三峡库区重点珍稀濒危植物物种 [J]. 中国三峡, 5: 26-34.]
WU H, CHEN PT, YUAN L, et al., 2011. Study on the technique of making paraffin section of fern [J]. Hubei Agric Sci, 50(18): 3767-3769. [吴华, 陈聘婷, 袁玲, 等, 2011. 蕨类植物石蜡切片制作技术探讨 [J]. 湖北农业科学, 50(18): 3767-3769.]
WYLIE RB, 1949. Variations in leaf structure among Adiantum pedatum plants growing in a rock cavern [J]. Am J Bot, 36(3): 282-287.
XU TQ, ZHENG Z, JIN YX, 1987. On the distribution characteristic of the variety Adiantun reinforme var. sinense [J]. J Wuhan Bot Res, 3: 247-252. [徐天全, 郑重, 金义兴, 1987. 论荷叶铁线蕨的分布特点 [J]. 武汉植物学研究, 5: 247-252.]
YANG C, ZHANG X, ZHOU C, et al., 2011. Root and stem anatomy and histochemistry of four grasses from the Jianghan Floodplain along the Yangtze River, China [J]. Flora, 206(7): 653-661.
YUAN WJ, ZHANG WR, SHANG FD, 2007. Study on the anatomical structure of vegetative organs of Coptis chinensis and its pertinence to sciophyte conditions [J]. J Henan Agric Univ (Nat Sci Ed), 37(2): 184-186. [袁王俊, 张维瑞, 尚富德, 2007. 黄连营养器官解剖结构与其阴生环境相关性研究 [J]. 河南农业大学学报(自然科学版), 37(2): 184-186.]
ZHANG GM, LIAO WB, DING MY, et al., 2013. Pteridaceae [M]//WU ZY, RAVEN PH, HONG DY (eds.). Flora of China. Beijing: Science Press;St. Louis: Missouri Botanical Garden Press: 169-256.
ZHANG ZH, WU XX, 2013. Study on the anatomy structure of five species of fern leaves and its adaptability to sciophyte environment [J]. J Huazhong Norm Univ (Nat Sci Ed), 47(6): 840-843. [张泽宏, 吴小霞, 2013. 5种蕨类植物叶片解剖結构及其对阴生环境的适应性研究 [J]. 华中师范大学学报(自然科学版), 47(6): 840-843.]
ZHOU SX, JIANG MX, HUANG HD, 2008. Spatial pattern of Adiantum reniform var. sinense, endemic to the Three Gorge Reservoir Region [J]. J Wuhan Bot Res, 26(1): 59-63. [周赛霞, 江明喜, 黄汉东, 2008. 三峡库区特有植物荷叶铁线蕨种群分布格局研究 [J]. 武汉植物学研究, 26(1): 59-63.]
(责任编辑 何永艳)
2299501186303
