
甜荞pin型花与thrum型花雌雄蕊发育的比较研究
2022-03-17 01:16:41王旋陈艳刘志雄
广西植物 2022年1期
王旋 陈艳 刘志雄



摘 要: 為从形态上研究甜荞(Fagopyrum esculentum)二型花发育的差异性,该文运用石蜡切片技术对甜荞pin型花和thrum型花大、小孢子发生和雌、雄配子体的发育过程进行观察比较。结果表明:(1)甜荞2种花的雌蕊、雄蕊发育的细胞学特征有很高的相似性,具直生胚珠、双珠被、厚珠心;大孢子四分体直线型排列,合点端为功能大孢子,蓼型胚囊。花药4室,花粉囊壁由表皮、药室内壁、一层中层和一层绒毡层组成,其中绒毡层发育为腺质型,小孢子母细胞减数分裂过程中的胞质分裂为同时型,小孢子四分体常为四面体型,但thrum型花花粉囊中偶见十字交叉型小孢子四分体;成熟花粉粒多数为2细胞型,少数为3细胞型。(2)从2种花雌蕊、雄蕊的发育进度来看,与pin型花相比,thrum型花小孢子发育相对较慢,雄配子体发育较快,但开花时,两者雌蕊、雄蕊均发育成熟。(3)综合甜荞两种花生殖发育的过程来看,其有许多相对原始的胚胎学特征,但也出现一些如直生胚珠、3细胞型花粉等相对进化的性状。该研究完善了甜荞二型花生殖发育的形态细胞学资料的同时,也为理解石竹目物种间的亲缘关系提供了胚胎学证据。
关键词: 甜荞, pin型花, thrum型花, 大、小孢子发生, 雌、雄配子体发育
中图分类号: Q945.6,S517
文献标识码: A
文章编号: 1000-3142(2022)01-0133-10
收稿日期: 2020-12-29
基金项目: 国家自然科学基金(31771867,31571736) [Supported by the National Natural Science Foundation of China (31771867, 31571736)]。
第一作者: 王旋(1994-),硕士研究生,主要从事作物种质资源创新与利用研究,(E-mail)2807467116@qq.com。
*通信作者: 刘志雄,博士,教授,主要从事植物发育遗传与种质创新研究,(E-mail)zxliu77@yahoo.com。
Development of stamens and carpels on pin type
and thrum type flowers of common buckwheat
WANG Xuan, CHEN Yan, LIU Zhixiong*
( College of Horticulture and Gardening, Yangtze University, Jingzhou 434025, Hubei, China )
Abstract: In order to reveal the morphological differences between pin type and thrum type flowers development of common buckwheat (Fagopyrum esculentum), megasporogenesis, microsporogenesis and development of female and male gametophytes in pin type and thrum type flowers of F. esculentum were investigated by using the paraffin section method. The results were as follows: (1) The cytological characteristics of pistil and stamen development of pin type and thrum type flowers were highly similar. Both type flowers had orthotropous ovule, inner and outer integument, and thickly nucellus. Moreover, the megaspore mother cells underwent meiosis and divided into linear type tetrads, and the one in the chalazal end was functional, which suggested that their development of embryo sac belonged to Polygonum type. The anther had four chambers and the anther wall was composed of epidermis, endothecium, one layer of middle layer and glandular tapetum. In addition, meiosis of microspore mother cell was followed by simultaneous cytokinesis and results in common tetrahedral tetrads. Furthermore, most of tetrads were tetrahedral, but a small number of decussate tetrads were also observed in thrum flower. Most of mature pollen grains were 2-cell type, and 3-cell type pollen grains were also observed in pin type and thrum type flowers. (2) Comparing the pistil and stamen developmental progress between the pin type and thrum type flowers, the development of microspores in thrum type flower was more slowly than that of the pin type flower, but the male gametophytes in thrum type flower grew faster. However, mature embryo sac and pollen grains were observed when pin type and thrum type flowers bloomed. (3) According to the reproduction development process of two type flowers, many relatively primitive embryological characteristics, and some relatively evolutionary embryological characteristics, such as orthotropous ovule and 3-cell type pollen grains, coexisted in common buckwheat. This study not only accumulates the morphological data of the distylous flower reproductive development in common buckwheat, but also provides embryological evidence for understanding the genetic relationships among the species of Caryophyllales.
Key words: Fagopyrum esculentum, pin type flower, thrum type flower, megasporogenesis and microsporogenesis, female and male gametophytes
甜荞(Fagopyrum esculentum)属石竹目(Caryophyllales)蓼科(Polygonaceae)荞麦属(Fagopyrum Mill.)的3个栽培种之一(唐宇等,2019),是兼食、药、观赏和土壤修复等多用途于一体的经济植物。甜荞的籽粒蛋白质、纤维素及微量元素的含量普遍高于水稻、小麦和玉米,且富含芦丁、槲皮素、山奈酚等黄酮类化合物,具有很高的营养价值和保健功效,作为一种可供功能食品开发的作物,近年来备受关注和重视(Joshi et al., 2019;张娇等,2020;章洁琼等,2020;Huda et al., 2021)。然而,一方面,甜荞作为典型的异型花柱植物,自然群体中短花柱长雄蕊(thrum)型和长花柱短雄蕊(pin)型花植株按1∶1分离,仅异型花间等高的柱头和花药间互相授粉才能正常结实(图版I),产量低,杂交育种因难,极大地限制了这一重要作物的推广应用。另一方面,籽粒作为甜荞主要产品器官,其产量高低和品质与甜荞生殖发育密切相关。因此,弄清甜荞生殖发育的规律对指导甜荞人工去雄、杂交育种和提升甜荞品质等有重要的科学意义和实际应用价值。在甜荞生殖发育研究方面,前人主要从外源激素对花芽分化的调控(金晓蕾等,2019),果皮开裂类型及其对籽粒早期萌发性状的影响(李鹏等,2020),甜荞等花柱资源与栽培品种杂交结实规律(陈稳良等,2020),甜荞雌蕊、雄蕊及花被片发育调控的分子机制(Li et al., 2017;Liu et al., 2019;张良波等,2021)等方面进行了深入的研究。在甜荞二型花柱异形分化的研究方面,前人主要围绕异型自交不亲和的遗传、基因组学、育种应用和二型花柱发育调控的分子机制展开(Barrett, 2019;Matsui & Yasui, 2020),而有关甜荞二型花发育的形态细胞学研究的资料甚少。
本文通过观察比较甜荞pin型与thrum型花雌蕊和雄蕊发育的形态细胞学过程,在系统分析2种花大、小孢子发生和雌、雄配子体发育异同的基础上,完善目前甜荞二型花生殖发育的形态细胞学资料的同时,为理解荞麦属植物间的进化和花型演变,乃至石竹目植物间亲缘关系的演化提供胚胎学证明。
1 材料与方法
1.1 材料
2019年9月初,挑选颗粒饱满的甜荞品种‘北早生’籽粒播种于长江大学作物遗传育种研究基地的塑胶花盆(21 cm×14 cm×20 cm),常规肥水管理。待花序开始分化和发育时,取不同叶龄植株上发育分化最快的花序,立即使用FAA固定液 [V(甲醛)∶V(冰乙酸)∶V(70%乙醇)=1∶1∶18]固定,4 ℃冰箱保存备用。
1.2 方法
甜荞‘北早生’多数植株的主茎上第3片真叶开始发育时可观察到明显的花序分化,且多数植株主茎上第4片真叶叶腋花序上的花最先开放。因此,取材从植株主茎上出现第3片真叶时开始,随机选取120个植株挂牌标记,同时从中随机选取20株挂牌植株摘除顶端花序,使用FAA固定液固定,做好标记,以便植株开花后确定样本的花型。余下100株挂牌标记植株则从主茎第4片真叶发育开始,主茎上每增加1片真叶,随机选取20株挂牌植株摘取第4片真叶叶腋的花序并用FAA固定液固定,直至第4片真叶叶腋花序中的第1朵花开放。同时做好标记,待所有挂牌标记植株开花后确定样本的花型。分别筛选pin型花和thrum型花不同叶龄植株各发育时期典型的花序样本,每个时期随机选取3个花序,投入一个新的盛有新鲜FAA固定液的小瓶混样,脱水前再次抽气后用50%乙醇润洗三次,后依次经70%、85%、95%、100%乙醇梯度脱水,每级2 h;经1/2乙醇+1/2二甲苯、100%二甲苯透明,每级2 h;浸蜡时先将透明后的材料转入纯二甲苯石蜡饱和溶液,在38 ℃恒温箱中放置15 h,后转入纯的液体石蜡,68 ℃恒温箱中放置12 h,每隔4 h换1次纯蜡。之后经包埋成型,用Leica RM2235石蜡切片机连续切片,切片厚度8 μm(李正理,1996)。用1%番红-0.1%固绿对染。加拿大中性树胶封片后在Caikon RCK-40C显微镜下观察细胞结构并拍照。
2 结果与分析
2.1 甜荞pin型花的小孢子发生与雄配子体的发育
甜荞pin型花植株的花序随主茎上第3片真叶开始发育时分化,此时可见花序部分花芽中近矩形的花药,花药的角隅可见细胞核明显的小孢子孢原细胞,有的角隅小孢子孢原细胞经平周分裂产生紧贴表皮的周缘细胞和靠内的造孢细胞(图版Ⅱ:A)。当主茎上第4片真叶开始发育时,其叶腋花序上部分花芽药室内可观察到小孢子母细胞,此时花药4室,呈蝶形,周缘细胞经平周分裂形成药室内壁、中层和多核绒毡层(图版Ⅱ:B);部分花芽药室内的小孢子母细胞间出现间隙并开始减数分裂,多核绒毡层细胞间隙明显(图版Ⅱ:C);而发育较快的花芽药室中小孢子母细胞通过减数分裂形成多核后再形成正四面体型四分体(图版Ⅱ:D,E)。待主茎第5片真叶开始发育时,第4片真叶叶腋花序上的部分花芽药室中小孢子四分体开始解体为单核小孢子,不同药室的小孢子发育存在不同步现象,此时中层消失,多核绒毡层进一步解体,部分绒毡层细胞与药室内壁产生明显间隙(图版Ⅱ:F)。当主茎第6片真叶开始发育时,第4片真叶叶腋花序上的部分花芽药室中可观察到单核小孢子体积增大、细胞壁增厚、中央大液泡明显,核从中间移向一側,发育为单核靠边期小孢子,此时多核绒毡层于药室内壁旁溶解并可见一些残迹,表皮细胞颜色加深(图版Ⅱ:G)。当主茎上第7片真叶开始发育时,第4片真叶叶腋花序上的部分花芽花粉囊中形成2细胞型花粉粒(图版Ⅱ:H),同时也可见少量3细胞型花粉粒(图版Ⅱ:I)。
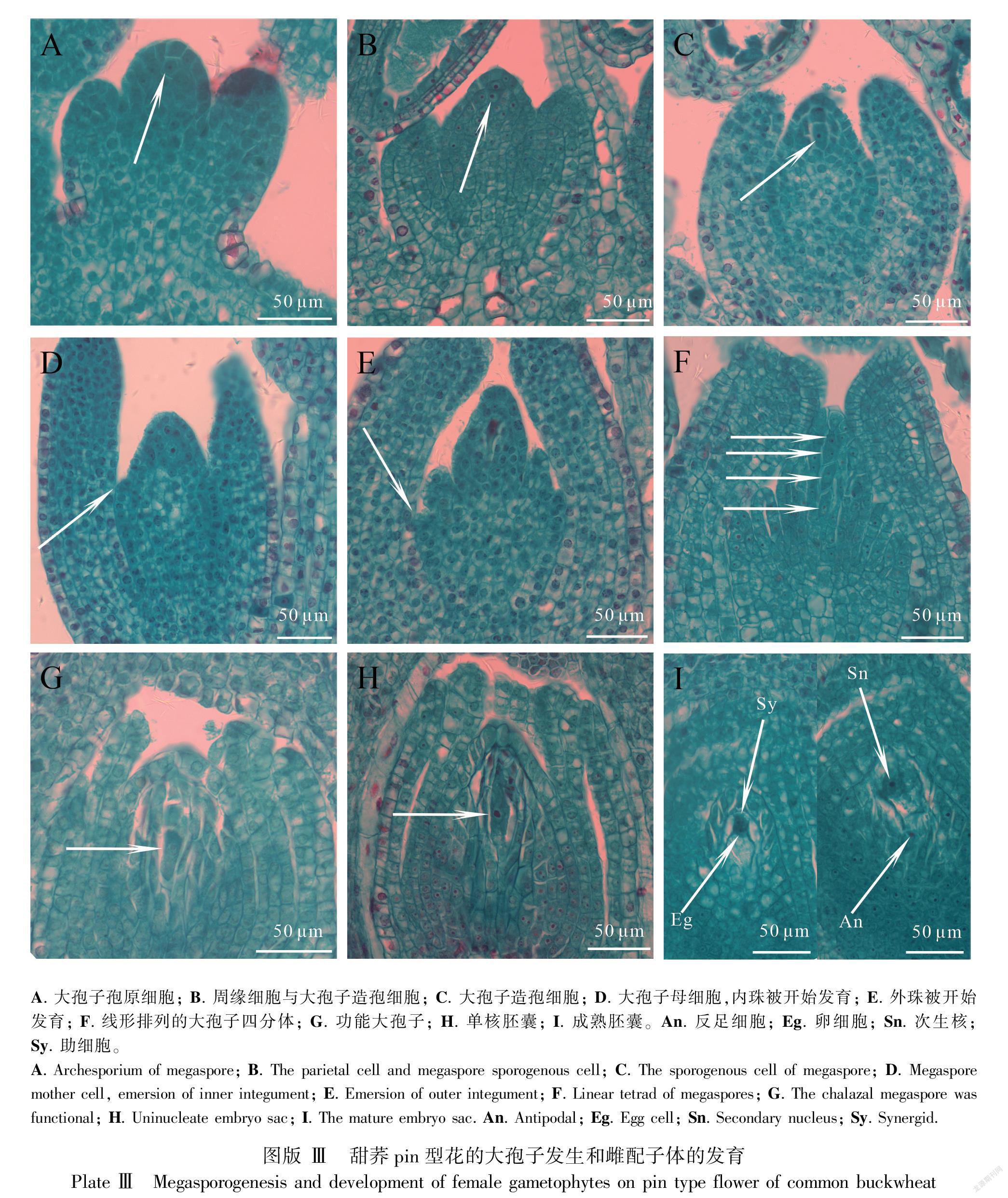
2.2 甜荞pin型花的大孢子发生与雌配子体的发育
当甜荞pin型花植株主茎上第4片真叶开始发育时,可见其叶腋花序上部分花芽珠心表皮下分化出一个细胞核明显、细胞体积较大的大孢子孢原细胞(图版Ⅲ:A);部分花芽珠心中的大孢子孢原细胞进行平周分裂产生一个周缘细胞和一个造孢细胞(图版Ⅲ:B);而发育较快的花芽大孢子造孢细胞逐渐变大,发育为大孢子母细胞(图版Ⅲ:C,D)。待主茎第5片真叶开始发育时,第4片真叶叶腋花序上的部分花芽中大孢子母细胞开始减数分裂,此时外珠被开始发育(图版Ⅲ:E)。当主茎第6片真叶开始发育时,第4片真叶叶腋花序上的部分花芽中可观察到大孢子母细胞已完成减数分裂形成直线排列的四分体(图版Ⅲ:F);发育较快的花芽中的大孢子四分体远离合点端的3个大孢子开始逐渐退化,合点端大孢子发育为功能大孢子,此时外珠被延伸至直生胚珠的顶端(图版Ⅲ:G)。当主茎上第7片真叶开始发育时,第4片真叶叶腋花序上的部分花芽可见体积较大、细胞质浓厚的单核胚囊,内珠被开始在直生胚珠的顶端靠拢形成珠孔(图版Ⅲ:H)。而发育较快的花芽中可见成熟的蓼型胚囊,珠孔端的3个细胞呈品字形,靠近珠孔端的2个助细胞和靠近合点端的1个卵细胞共同构成卵器,此时胚囊中的2个极核融合为次生核(图版Ⅲ:I)。
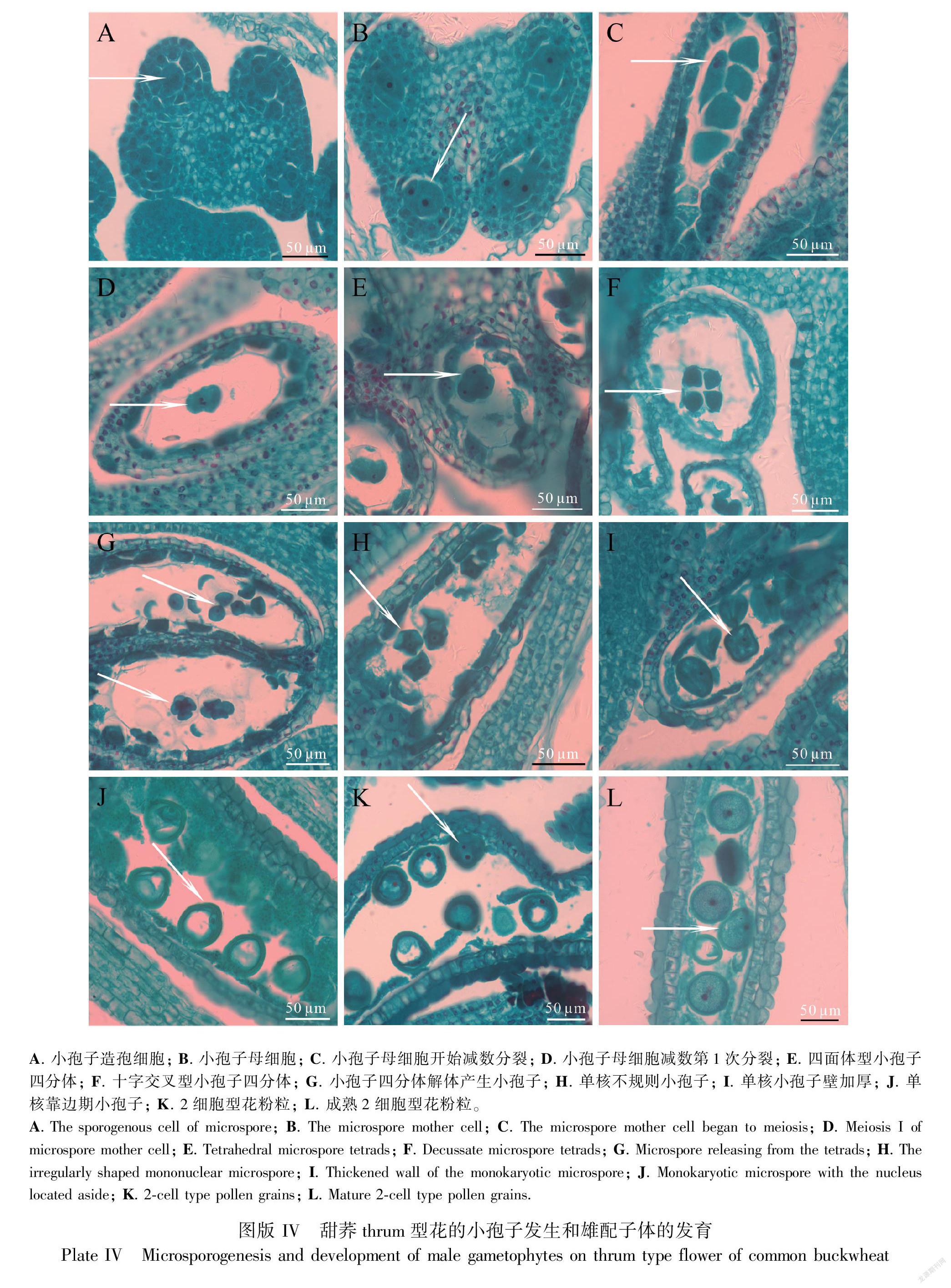
2.3 甜荞thrum型花的小孢子发生与雄配子体的发育
甜荞thrum型花植株的花序也随主茎上第3片真叶开始发育时分化,此时花序部分花芽中出现蝶形的花药,花药角隅可见体积较大的小孢子造孢细胞,造孢细胞周围的周缘细胞平周分裂进度不一致(图版IV:A);发育较快的花芽花粉囊中可观察到明显的小孢子母细胞(图版IV:B)。当主茎第4片真叶开始发育时,可见其叶腋花序上部分花芽药室内的小孢子母细胞开始减数分裂形成四面体型四分体,稀十字交叉型小孢子四分体(图版IV:C-F);发育较快的花芽药室中部分小孢子四分体开始解体(图版IV:G)。当主茎上第5片真叶开始发育时,第4片真叶叶腋花序上的部分花芽花粉囊中可观察到许多形状不规则的单核小孢子(图版IV:H);随后单核小孢子体积增大,形状变圆并产生显著的壁(图版IV:I)。当主茎第6片真叶开始发育时,第4片真叶叶腋花序上的部分花芽小孢子处在单核靠边期(图版IV:J);发育较快的花芽药室中可观察到2细胞型花粉粒(图版IV:K)。当主茎第7片真叶发育时,发育较快的花芽中可见以2细胞型花粉粒为主的成熟花粉(图版IV:L)。
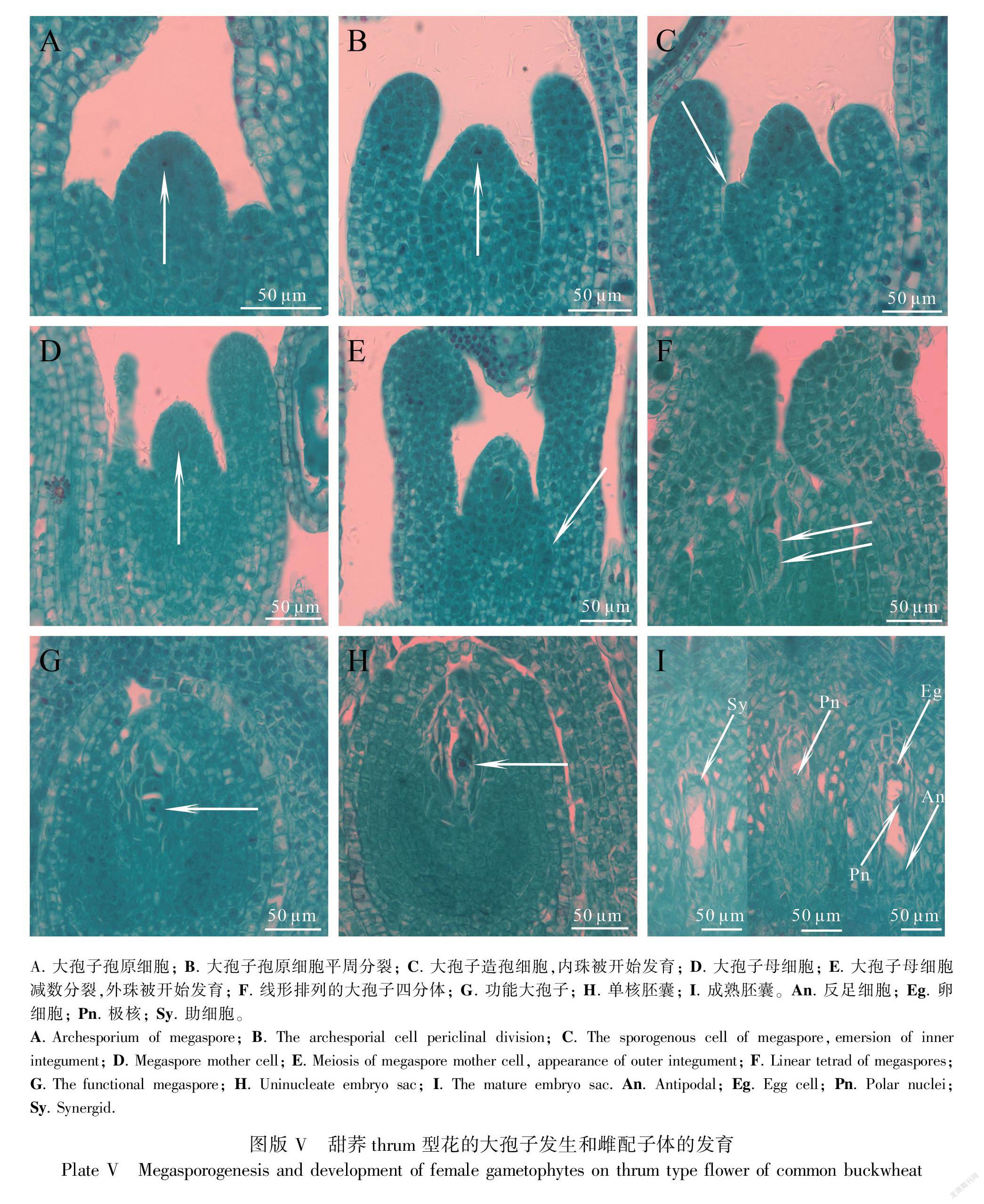
2.4 甜荞thrum型花的大孢子发生与雌配子体的发育
当甜荞thrum型花植株主茎第3片真叶开始发育时,可见花序部分花芽胚囊珠心中出现一个细胞核明显、相对体积较大的孢原细胞(图版V:A)。当主茎第4片真叶开始发育时,可见其叶腋花序上部分花芽大孢子孢原细胞开始平周分裂(图版V:B);部分花芽大孢子孢原细胞平周分裂产生一个周缘细胞和一个造孢细胞,随着造孢细胞体积增大,内珠被开始发育(图版V:C);部分花芽大孢子造孢细胞发育为大孢子母细胞(图版V:D)。当主茎上第5片真叶开始发育时,第4片真叶叶腋花序上的部分花芽大孢子母细胞仍在减数分裂,外珠被开始发育(图版V:E)。当主茎第6片真叶开始发育时,第4片真叶叶腋花序上的部分花芽大孢子母细胞完成减数分裂形成线形四分体(图版V:F);部分花芽近珠孔端3个大孢子先后退化,合点端的大孢子发育为功能性大孢子(图版V:G);发育较快的花芽近珠孔端的3个大孢子退化消失,合点端的功能性大孢子体积增大,细胞核变大,发育为单核胚囊(图版V:H)。当主茎第7片真叶发育时,发育较快的花芽中单核胚囊进行3次有丝分裂形成8核7细胞的成熟蓼型胚囊(图版V:I)。
2.5 甜荞pin型花与thrum型花雌蕊、雄蕊发育的比较
甜荞两种类型花的雌蕊、雄蕊发育的基本胚胎学特征进程有一定的差异。与pin型花相比,thrum型花小孢子发育相对较慢,雄配子体发育较快,但2种花开花时,其雌蕊、雄蕊均发育成熟。具体相关性状见表1。
3 结论与讨论
与pin型花相比,thrum型花小孢子发育相对较慢,雄配子体发育较快,其原因可能是thrum型花具有更长的花丝,小孢子发育时雄蕊花丝的伸长需要消耗更多的营养,从而导致thrum型花的小孢子发育较慢。但是pin型和thrum型花雄蕊花药在小孢子四分体时期,2种花的雌蕊胚珠中均出现了大孢子母细胞,此时雌蕊的花柱开始发育;而与thrum型花相比,pin型花具有更长的花柱,同样需要消耗更多的营养,可能导致pin型花的雄配子体发育相对较慢。与pin型花相比,thrum型花的花粉粒稍大,与张玉霞和陈庆富(2002)的研究结果一致。张大爱等(2015)通过研究甜荞结实性及授精过程,发现pin型花较thrum型花更容易授粉结实,孙淑范等(2021)发现二型花柱植物连翘(Forsythia suspensa)也表現为长花柱的更易授粉结实。我们通过形态细胞学分析发现,pin型花在开花前,成熟胚囊中的极核已融合为次生核,能更好地为双受精做准备,为pin型花较thrum型花更容易授粉的结果提供了更直观的形态细胞学证据。
二型花柱异花传粉的甜荞与同型花柱自花传粉的苦荞(Fagopyrum tataricum)主要胚胎学特征有较高的相似性,与苦荞相比,甜荞thrum型花小孢子四分体多为四面体型,偶见十字交叉型;pin
型花和thrum型花成熟花粉粒多为2细胞型,也有少量3细胞型成熟花粉,与苦荞小孢子四分体仅为四面体型,成熟花粉粒仅为2细胞型有明显区别(吴凯等,2017)。在石竹目其他植物中,目前仅发现苋科(Amaranthaceae)植物甜菜(Beta vulgaris)、白花丹科(Plumbaginaceae)补血草属(Limonium Mill.)中的几种植物成熟花粉为3细胞型(申家恒等,1986;李伟等,2006;周玲玲等,2007;孔冬瑞等,2008);而花药中同时存在四面体型小孢子四分体和十字交叉型小孢子四分体的现象在核心石竹目苋科植物华北驼绒藜(Ceratoides arborescens)中也有发现(卢立娜等,2008)。这说明甜荞与苋科、白花丹科植物有较近的亲缘关系。
甜荞花药4室、双珠被、厚珠心、合点端的大孢子为功能大孢子和胚囊为蓼型等是相对原始的性状(王伏雄,1980)。但是与石竹目植物常见的倒生胚珠不同,甜荞胚珠的着生方式为相对进化的直生型,而白花丹科植物为拳卷型(胡适宜,2005;李伟等,2006;周玲玲等,2007;孔冬瑞等,2008),说明蓼科与白花丹科同为非核心石竹目,我们的研究结果和分子系统学的分类结果有很好的印证(Angiosperm Phylogeny Group et al., 2016)。
在目前报道的石竹目其他科植物中,也有少数胚胎学特征相对原始,出现一些衍生性状的情况。如核心石竹目商陆科(Phytolaccaceae)植物美洲商陆(Phytolacca americana)和商陆(P. acinosa)都具有花药4室、倒生胚珠、双珠被、厚珠心、合点端的大孢子為功能大孢子和蓼型胚囊等典型的原始胚胎学特征,但其大孢子减数分裂只形成3个大孢子,是典型的三分体(辛洪婵等,2000;李祖任等,2015);另一非核心石竹目植物柽柳 (Tamarix chinensis)具双珠被、厚珠心、倒生胚珠等原始特征的同时,其还具胚珠多数、贝母型胚囊等衍生性状(王仲礼等,2005)。
从甜荞pin型花与thrum型花大、小孢子发生和雌、雄配子体发育过程来看,两者有较大的相似性,但也存在一定的差别。从两者雌蕊、雄蕊的发育进度来看,与pin型花相比,thrum型花小孢子发育相对较慢,雄配子体发育较快;thrum型花花粉囊中偶见十字交叉型小孢子四分体,但pin 型花中全为四面体型小孢子四分体。综合分析甜荞和其他石竹目植物胚胎学特征发现,石竹目植物的胚胎学特征多表现相对原始的特征,但也存在3细胞型花粉、直生和拳卷型胚珠、大孢子三分体和贝母型胚囊等衍生性状,体现了石竹目植物在演化上的复杂性。
参考文献:
ANGIOSPERM PHYLOGENY GROUP, CHASE MW, CHRISTENHUSZ MJM, et al.,2016. An update of the angiosperm phylogeny group classification for the orders and families of flowering plants: APG IV [J]. Bot J Linn Soc, 181(1): 1-20.
BARRETT SCH, 2019. ‘A most complex marriage arrangement’: recent advances on heterostyly and unresolved questions [J]. New Phytol, 224(3): 1051-1067.
CHEN WL, LI XL, SHI XH, et al., 2020. A preliminary study on the hybridization of common buckwheat isostyle resources and cultivated buckwheat [J]. J Plant Genet Resour, 21(4): 1030-1035. [陈稳良, 李秀莲, 史兴海, 等, 2020. 甜荞等花柱资源与栽培甜荞杂交初步研究 [J]. 植物遗传资源学报, 21(4): 1030-1035.]
HU SY, 2005. Reproductive biology of angiosperms [M]. Beijing: Higher Education Press: 94. [胡适宜, 2005. 被子植物生殖生物学 [M]. 北京: 高等教育出版社: 94.]
HUDA MN, LU S, JAHAN T, et al., 2021. Treasure from garden: Bioactive compounds of buckwheat [J]. Food Chem, DOI: https: //doi.org/10.1016/j.foodchem.2020.127653.
JIN XL, LIU JH, LUO ZW, et al., 2019. Effects of PP333 and 6-BA on the bud differentiation and endogenous hormones of common buckwheat [J]. Acta Bot Boreal-Occident Sin, 39(11): 1988-1997. [金晓蕾, 刘景辉, 罗中旺, 等, 2019. 叶面喷施PP333和6-BA对甜荞花芽分化及内源激素的影响 [J]. 西北植物学报, 39(11): 1988-1997.]
JOSHI DC, CHAUDHARI GV, SOOD S, et al., 2019. Revisiting the versatile buckwheat: reinvigorating genetic gains through integrated breeding and genomics approach [J]. Planta, 250(3): 783-801.
KONG DR, WANG ZL, LIU LD, et al., 2008. Megasporogenesis, microsporogenesis and development of the female and male gemetophyte of Limonium frantii [J]. J Chin Electron Microsc Soc, 27(3): 223-228. [孔冬瑞, 王仲礼, 刘林德, 等, 2008. 紫花补血草的大小孢子发生及雌雄配子体发育 [J]. 电子显微学报, 27(3): 223-228.]
LI LY, FANG ZW, LI XF, et al., 2017. Isolation and characterization of the C-class MADS-box gene from the distylous pseudo-cereal Fagopyrum esculentum [J]. J Plant Biol, 60(2): 189-198.
LI P, CHEN QJ, SHI TX, et al., 2020. Cracking types of common buckwheat pericarp and its effect on early germination traits [J]. Guihaia, 40(7): 954-962. [李鹏, 陈其皎, 石桃雄, 等, 2020. 甜荞果皮开裂类型及其对籽粒早期萌发性状的影响 [J]. 广西植物, 40(7): 954-962.]
LI W, ZHOU LL, LIU P, 2006.Megasporogenesis, microsporogenesis and developments of male and female gametophytes in Limonium otolepis [J]. Acta Bot Boreal-Occident Sin, 26(7): 1355-1362. [李伟, 周玲玲, 刘萍, 2006. 耳叶补血草大、小孢子发生与雌、雄配子体的发育 [J]. 西北植物学报, 26(7): 1355-1362.]
LI ZL, 1996. Plant tissue sectioning technology [M]. Beijing: Peking University Press: 129-137. [李正理, 1996. 植物组织制片学 [M]. 北京: 北京大学出版社: 129-137.]
LI ZR, YAN SL, LIAO HM, et al., 2015. The megasporogenesis and microsporogensis of invasive weeds Phytolacca americana and formation of its male and female gametophyte [J]. Acta Agric Boreal-Sin, 30(S1): 81-86. [李祖任, 晏升禄, 廖海民, 等, 2015. 入侵杂草美洲商陆大小孢子的形成和雌雄配子体的发育 [J]. 华北农学报, 30(S1): 81-86.]
LIU ZX, FEI Y, ZHANG KB, et al., 2019. Ectopic expression of a Fagopyrum esculentum APETALA1 ortholog only rescues sepal development in arabidopsis ap1 mutant [J]. Int J Mol Sci, 20(8): 2021.
LU LN, HE X, YI J, et al., 2008. Megasporogenesis, microsporogenesis and development of male and female gametophytes in Ceratoides arborescens [J]. Acta Bot Boreal-Occident Sin, 28(7): 1319-1325. [盧立娜, 贺晓, 易津, 等, 2008. 华北驼绒藜大小孢子的发生及雌雄配子体发育过程的解剖学研究 [J]. 西北植物学报, 28(7): 1319-1325.]
MATSUI K, YASUI Y, 2020. Buckwheat heteromorphic self-incompatibility: genetics, genomics and application to breeding [J]. Breed Sci, 70(1): 32-38.
SHEN JH, LI HR, YIN H, 1986. Megasporogenesis and microsporogenesis and the development of their female and male gametophyte in sugar beet [J]. Acta Agron Sin, 12(3): 205-211. [申家恒, 李慧蓉, 殷华, 1986. 甜菜大、小孢子发生与雌、雄配子体发育 [J]. 作物学报, 12(3): 205-211.]
SUN SF, LI JX, SUN XL, et al., 2021. Study on characteristics of breeding system in distylous Forsythia suspensa [J]. Acta Bot Boreal-Occident Sin, 41(2): 317-322. [孙淑范, 李佳欣, 孙霞玲, 等, 2021. 连翘花柱二型的繁育系统特征研究 [J]. 西北植物学报, 41(2): 317-322.]
TANG Y, SHAO JR, ZHOU ML, 2019. A taxonomic revision of Fagopyrum Mill. from China [J]. J Plant Genet Resour, 20(3): 646-653. [唐宇, 邵继荣, 周美亮, 2019. 中国荞麦属植物分类学的修订 [J]. 植物遗传资源学报, 20(3): 646-653.]
WANG FX, 1980. New progress on plant embryology [J]. Plants, 7(2): 30-31. [王伏雄, 1980. 植物胚胎学研究的新进展(续) [J]. 植物杂志, 7(2): 30-31.]
WANG ZL, LIU LD, FANG YM, 2005. Observations on megasporogenesis, microsporogenesis and development of female and male gametophytes of Tamarix chinensis (Tamaricaceace) [J]. Bull Bot Res, 25(2): 144-150. [王仲礼, 刘林德, 方炎明, 2005. 柽柳大、小孢子发生和雌、雄配子体发育的观察 [J]. 植物研究, 25(2): 144-150.]
WU K, ZUO ZJ, XIA SY, et al., 2017. Observation of mega-and microsporogenesis and development of male and female gametophytes in Fagopyrum tataricum [J]. J Yangtze Univ(Nat Sci Ed), 14(10): 34-37. [吴凯, 左振君, 夏胜应, 等, 2017. 九江苦荞大小孢子发生与雌雄配子体发育观察 [J]. 长江大学学报(自然科学版), 14(10): 34-37.]
XIN HC, TAO SR, CAO YF, et al., 2000. The megasporogenesis and microsporogenesis of Phytolacca acinosa Roxb. and formation of its male and female gametophytes [J]. J Huazhong Agric Univ, 19(2): 179-181. [辛洪婵, 陶世蓉, 曹玉芳, 等, 2000. 商陆大小孢子的发生和雌雄配子体的形成 [J]. 华中农业大学学报, 19(2): 179-181.]
ZHANG DA, DU Y, QIAN YP, et al., 2015. Fecundity of common buckwheat and dynamic growth of pollen tube in pollination and fertilization [J]. J Northwest A & F Univ(Nat Sci Ed), 43(8): 103-108. [张大爱, 杜莹, 钱一萍, 等, 2015. 甜荞结实性及授粉受精过程中花粉管的生长动态 [J]. 西北农林科技大学学报(自然科学版), 43(8): 103-108.]
ZHANG J, WANG X, ZHANG LB, et al., 2020. Expression analysis of the FaesAP2B in mutant buckwheat with long pistil and long stamen [J]. Bull Bot Res, 40(2): 266-273. [張娇, 王旋, 张良波, 等, 2020. FaesAP2B基因在甜荞长雌蕊长雄蕊突变体lpls的表达分析 [J]. 植物研究, 40(2): 266-273.]
ZHANG JQ, ZOU J, LU Y, et al., 2020. Analysis and evaluation of main functional components of different buckwheat varieties [J]. Seed, 39(2): 107-112. [章洁琼, 邹军, 卢扬, 等, 2020. 不同荞麦品种主要功能成分分析及评价 [J]. 种子, 39(2): 107-112.]
ZHANG LB, WANG X, QIAN CX, et al., 2021. Cloning and expression analysis of FeFUL2 gene from buckwheat (Fagopyrum esculentum) [J]. Guihaia, 41(4): 591-597. [张良波, 王旋, 钱成旭, 等, 2021. 甜荞FeFUL2基因的克隆与表达分析 [J]. 广西植物, 41(4): 591-597.]
ZHANG YX, CHEN QF, 2002. Study on pollen morphology of six kinds of buckwheat flowers by means of electric microscope [J]. Guihaia, 22(3): 232-236. [张玉霞, 陈庆富, 2002. 六个不同类型荞麦花花粉粒形态的电镜观察比较研究 [J]. 广西植物, 22(3): 232-236.]
ZHOU LL, LI W, LIU P, 2007.Megasporogenesis, microsporogenesis and the development of the female and male gametophytes of Limonium gmelinii [J]. Bull Bot Res, 27(4): 402-407. [周玲玲, 李伟, 刘萍, 2007. 大叶补血草的大、小孢子发生与雌、雄配子体的发育 [J]. 植物研究, 27(4): 402-407.]
(责任编辑 何永艳)
3190501186543
