
植物凋落物分解对土壤有机碳稳定性影响的研究进展
2022-03-15 05:20:34苏卓侠苏冰倩上官周平
水土保持研究 2022年2期
苏卓侠, 苏冰倩, 上官周平
(西北农林科技大学 黄土高原土壤侵蚀与旱地农业国家重点实验室, 陕西 杨凌 712100)
土壤作为陆地生态系统最大的碳汇载体(1 500 Gt),其碳储量超过了植被和大气碳储量的总和[1],在表层30 cm土壤中,有机碳储量每年增加4‰,就会抵消掉每年人为排放CO2增量[2]。关于土壤中有机碳储存的一个关键问题是其在土壤中保留的时间长短即有机碳稳定性问题[3]。增加有机碳的存储和稳定是“双赢”的策略:除了缓解气候变化外,更高的有机碳还有助于改善土壤肥力、土壤结构稳定性和生产力。因此,土壤固碳效应及其稳定机制也成为当前土壤学、植物营养学和生态学研究的热点。
在生态系统内,植物产生凋落物并将其归还给土壤,凋落物是分解者物质和能量的来源,是连接土壤碳库和植物碳库的重要组分,对陆地生态系统碳循环具有重大影响[4]。凋落物作为植物向土壤输入有机碳的主要途径,其分解过程中一部分碳以CO2的形式进入到大气中,另一部分以有机碳的形式输入到土壤中[5],源于凋落物的碳可能会导致原土壤有机碳(SOC)的分解(激发效应)或增加。目前对源于凋落物碳是如何固存在土壤中并稳定存在的机制仍然缺乏清晰的认识。因此,本文通过综述凋落物输入土壤后对土壤有机碳及其组分、土壤呼吸以及微生物特性的影响,以期厘清植物凋落物-土壤-微生物在稳定有机碳形成过程中的作用,从而全面理解土壤碳循环过程。
1 凋落物分解研究方法
凋落物是指在生态系统中由植物组分产生并归还到土壤表面,为分解者提供能量和物质,从而维持生态系统功能的有机物质[6]。凋落物分解研究方法起步较早,常用的有尼龙网袋法、小容器法、室内分解培养法等[7]。尼龙网袋法操作简单,能最大程度模拟凋落物自然分解状态,目前常用于测定凋落物分解速率,但它耗时较长,且网袋孔径大小会限制土壤动物及微生物的活动,使得凋落物分解减慢。小容器法使内部微环境与外部环境完全隔绝,不能完全模拟自然环境下的分解。室内分解培养的方法即在室内模拟枯落物的自然分解状态,与野外试验相比,室内培养试验温度、水分等生境条件可人为控制,枯落物分解速率较快,可在短时间内使土壤总有机碳发生变化,但所得数据在非自然状态下产生,只具有相对意义。在森林生态系统中,经常用凋落物添加和去除试验(DIRT,Detritus input and removal treatments)来研究凋落物分解,又称凋落物添加和去除转移试验(DIRT),它是研究植物凋落物输入来源和速率如何影响森林土壤有机质(SOM)和养分的积累和动态的重要手段[8]。
随着科技的发展,近红外光谱分析技术(near infrared spectroscopy,NIRS)、核磁共振技术(nuclear magnetic resonance, NMR)、气相色谱-质谱(gas chromatography-mass spectrometry, GC/MS)等技术已经应用于凋落物分解及土壤固碳方面的研究。近红外光谱分析技术(NIRS)是利用化学物质在近红外光谱区的光学吸收特性,快速测定某种样品中的一种或多种化学成分含量和特性的技术[9]。核磁共振技术(NMR)具有无损和非侵入等特点,可了解凋落物不同有机碳组分的分解特征,从而得知不同凋落物组分在土壤中的相对稳定性,及其对SOC形成和稳定的贡献程度[10]。同位素法可以让有机物处在自然环境中,不会改变它们的生境要素。Almeida等[11]提取了同位素标记(13C)的桉树(Eucalyptusgrandis×E.urophylla)植物的叶子、树枝、树皮和根,然后采用GC/MS系统观测了各组分的分子组成,将植物凋落物的生物化学组分、分解特性和土壤有机质的形成联系了起来,其耦联分析方法为深入探究凋落物分解在SOC形成、稳定中的作用提供了重要的技术手段,对于定量理解植物凋落物的转化和稳定SOC的形成等生物地球化学过程具有重要意义。
2 土壤有机碳组分及土壤有机碳稳定性
土壤有机碳(SOC)在全球碳循环中扮演了重要角色,通过储碳和充当温室气体排放的碳库来缓解潜在的气候变化[12]。土壤中有机碳相对组成的差异往往会导致土壤有机质的稳定性不同[1]。Parton等[13]将SOC分为活性碳库、慢性碳库及惰性碳库等。其中,活性碳库也被称为易分解碳库,是容易被土壤微生物分解矿化的碳库,对植物养分供应起着直接作用,活性碳库的周转率为几周、几月或者几年;慢性碳库活性介于活性和惰性碳库之间,也被称为难分解有机碳,其周转率长达几十年;惰性碳库是指存在于土壤中的惰性碳和极难分解的被物理保护的部分有机碳,其物理化学性质非常稳定,转换时间为数百至数千年。活跃的和稳定的有机碳在土壤碳库周转和养分循环中起不同的作用。
不同组分碳代表着不同功能碳库,具有不同周转期,通常活性有机碳对生境因子变化的反应比总有机碳更为敏感[14]。根据土壤有机碳提取方法的不同,可以从物理-化学-生物化学等不同的视角对有机碳进行分组(表1)。物理组分是基于有机碳化合物与矿物质结合状态进行分组,化学分组是基于不同的浸提剂与土壤有机碳化合物的相互作用,生物分组是区分活体细胞与死亡细胞。
近年来对生态系统SOC稳定性机制的研究,一直是生态学与土壤学关注的热点领域,但仍未达成一致意见[18]。土壤科学经典的观点认为,稳定的有机质由腐殖质等化合物组成,由于其复杂的高度芳香的结构而抵抗分解。Marschner等[19]假设认为稳定的SOC库由难分解的有机分子组成,如木质素等,由于其复杂的化学结构,在植物组织腐烂过程中被选择性地保存下来。Lorenz等[20]认为有机碳长期稳定经历两个重要的过程:(1) 物理保护过程,即分解者或其水溶性降解酶在空间上无法接近有机碳;(2) 有机矿物配合物和有机-金属相互作用,即有机碳与矿物、金属离子和其他有机物的相互作用。物理保护可能会使有机碳分解延缓几十年到几百年,而有机矿物配合物或有机金属相互作用可能是几个世纪到几千年大多数有机碳稳定的原因[21]。随着研究手段与技术的迅速发展,更多研究关注微生物对稳定土壤有机碳的调控,并提出新的SOC形成和稳定机制——“土壤微生物碳泵(microbial carbon pump, MCP)概念体系”[22],即微生物通过同化作用将有机碳合成为自身生物量,再通过残留物形式不断输入到稳定土壤有机碳库中。这一概念体系强调了土壤微生物同化合成产物对于土壤稳定有机碳库形成的重要作用,对深入认识陆地碳汇功能和应对气候变化具有重要意义。

表1 土壤有机碳组分
3 凋落物分解对土壤有机碳库稳定性的作用机理
凋落物是生态系统的重要组成部分,凋落物分解作为生态系统物质循环的核心过程,参与生态系统中养分的周转与循环,对生态系统健康可持续发展起着重要作用[23],凋落物的分解也影响着生态系统碳平衡。凋落物在土壤碳动态转化中发挥着重要作用(图1)。
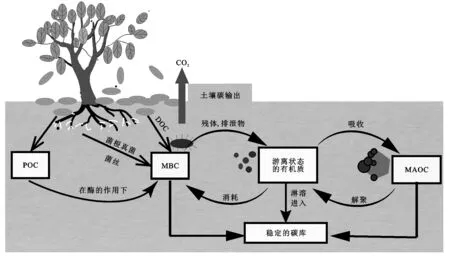
图1 凋落物碳在土壤碳库的动态转化
3.1 凋落物对土壤有机碳及活性碳组分的影响
凋落物是影响SOC周转的重要驱动力,是SOC形成的重要来源(图1)。Palviainen等[24]研究表明全球森林生态系统每年通过凋落叶分解归还到土壤有机碳含量约为50 Gt。凋落物分解对有机质形成的贡献主要通过两个路径:(1) 高质量凋落物(通常是水溶性碳)快速降解,在稳定在矿质结合态有机质之前,被微生物或其他土壤有机体同化。(2) 植物结构性物质被机械拉开,直接通过物理方式掺入下层矿物土壤的颗粒有机质中[25]。较高的生物量或凋落物产量并不意味着较高的土壤有机碳储量。凋落物分解过程中产物去向决定了土壤有机碳的赋存状态,高质量的凋落物其分解产物向土壤转移的比例更高[26]。
添加凋落物对土壤有机碳含量的影响表现为增加或影响不显著[27]。Mitchell等[28]使用13C同位素标记作物残体,追踪新鲜残余物进入到土壤组分的命运,结果表明植物残体输入使土壤中源于凋落物有机碳增加了4~5倍,同时也抑制了原土壤有机碳的激发效应。源于凋落物的溶解性有机碳(DOC)被微生物群落所固定,并固存在森林土壤中[29]。Zhong等[4]研究表明凋落物添加显著增加了各恢复阶段土壤水溶性碳氮的含量,说明这些来自凋落物分解的养分输入增强了其在土壤中的浓度。此外,凋落物分解过程中所释放的挥发性有机化合物(Volatile organic compound,VOC)能够直接扩散到土壤基质中,具有促进稳定矿质结合态有机质形成的潜力,VOC可能是植物来源的碳进入土壤并促进有机质形成的重要机制[25]。
3.2 凋落物分解对土壤呼吸及激发效应的影响
凋落物是土壤呼吸的重要碳源,凋落物分解是养分循环的核心过程(图2)。它通过改变土壤微环境条件[30]、增加土壤碳的有效性[31],影响根系生长和土壤微生物结构和功能等[32],进而影响到土壤呼吸过程。

图2 凋落物分解与土壤呼吸间的关系
关于凋落物分解对土壤呼吸的影响已有许多工作,不少研究表明添加凋落物能促进土壤呼吸,如Zhong等[33]对子午岭完整的次生林演替序列进行研究,发现未添加凋落物时,由于演替后期土壤养分有效性提高,演替后期的土壤呼吸更高;但添加凋落物后,演替前期土壤呼吸最高,表明在演替早期微生物分解能力较强,这一发现表明新碳源(凋落物)的添加可能在形成微生物功能方面发挥重要作用。Yan等[34]研究同样表明微生物呼吸作用随演替而增加,并且添加凋落物显著促进了微生物呼吸(16.5%~72.9%),特别是在演替初期(草地和灌丛)。凋落物输入和清除处理(DIRT)试验为研究地上凋落输入对土壤呼吸的影响及植物碳输入对土壤的贡献提供了一个很好的机会。Wang等[35]对杉木林进行了4 a的碳输入模拟,发现减少碳输入会显著减少土壤呼吸。土壤异养呼吸(由微生物分解的土壤有机质)占土壤总呼吸的52.1%,地上新近凋落物分解和地下自养呼吸(活根和相关微生物)分别占土壤总呼吸的23.7%,24.2%。众多研究均表明凋落物添加能促进微生物呼吸,但不同类型凋落物分解对促进土壤微生物呼吸的程度以及对固碳的影响仍有待进一步探究。
激发效应(priming effect, PE)是连接土壤中碳输入和输出的机制之一,实际上PE被定义为在加入一定量的新鲜有机质后土壤有机质分解速率的变化[36]。外源基质输入的改变会通过激发效应改变土壤有机碳储量。添加基质引起有机物分解的增加为正激发效应,反之为负激发效应[37]。PE的大小和方向取决于添加的底物的量和化学计量及底物的化学结构,不稳定底物比顽固性底物具有更大的激发效应[38]。关于激发效应的本质可用两种替代机制来解释,即“化学计量分解”理论(‘stoichiometric decomposition’ theory)和“微生物氮挖掘”理论(‘microbial nitrogen mining’ theory)。“化学计量分解”理论[39]指当底物化学计量与微生物需求相匹配时,快速生长的微生物(R-策略)可能会由于微生物活性的增加而刺激有机质的分解。而“微生物氮挖掘”理论(也被定义为优先底物利用)[40]强调当外源碳充足时,PE的方向取决于氮的可用性。具体来说,基质中高比例的碳氮比可能导致正的PE,因为微生物必须分解更多的土壤腐殖质来获得足够的氮来满足它们的需求。相比之下,基质中较低的碳氮比通常会导致负的PE,从而限制了有机碳的分解。最初认为微生物对土壤有机质分解的贡献受微生物群落的生长策略和氮有效性的控制,但随着分解的进行,这两种机制也可能随着时间的推移而变化,这可能与不同演替阶段微生物群落和功能有关。
此外,增加底物输入引起的SOM分解温度敏感性(Q10)的变化也决定了陆地碳平衡对全球变暖的响应[41]。目前,凋落物添加对土壤微生物呼吸和温度敏感性的影响尚无一致结论。较多的研究发现难分解有机碳的Q10要大于易分解有机碳的Q10,如Chen等[42]研究表明秸秆添加降低了土壤微生物呼吸的温度敏感性,但与秸秆类型无关。Wang等[41]探究在不同林型中增加凋落物对土壤有机质分解的温度敏感性的影响,结果表明针叶林土的Q10由不添加凋落物处理的2.41降低到增加凋落物处理的2.05,阔叶林土壤Q10由2.14降低到1.82,表明增加凋落物降低了Q10。这一现象可以用碳-质量-温度(CQT)假说来进行解释,即复杂底物的分解需要更高的总活化能,因此比简单的碳底物的分解对温度的升高更为敏感[43]。但也有研究发现难分解有机碳的Q10并不比易分解有机碳的Q10值高,如两种凋落物处理的土壤碳分解均比单纯土壤处理对温度更敏感(Q10更高)[44]。所以,关于凋落物分解对土壤有机质分解的温度敏感性仍有待进一步研究。改善全球变暖条件下土壤有机碳动态的预测对于理解温度和凋落物输入对SOC分解的共同作用具有重要意义。
3.3 凋落物分解对土壤微生物群落结构及酶活性的影响
凋落物分解的主要参与者是土壤微生物,在凋落物分解过程中微生物把大分子有机物分解为能够被植物吸收利用的小分子物质,对植物生长和土壤改良等起着重要作用。
首先,凋落物自身特性会影响到土壤微生物群落。凋落物的数量与质量[45]、组成[46]、多样性等[47]都会影响着土壤微生物的群落结构。陈法霖等[48]研究表明,添加桉树凋落物的土壤中细菌、真菌、放线菌以及磷脂脂肪酸的总丰度显著高于不添加凋落物的土壤。源于凋落物的挥发性有机组分(VOC)也能影响细菌和真菌群落的多样性和组成[25]。真菌和细菌的比值常用来反映分解过程中微生物群落结构的变化及评价生态系统的稳定性,真菌/细菌的比值越高,表明真菌的生物量及菌丝体增加,能够固定更多的养分,使得土壤生态系统更加稳定。其次,凋落物分解通过改变土壤环境影响到土壤微生物群落结构。凋落物初始养分含量及分解过程中养分的释放对土壤微生物的生长与分布具有一定的选择作用,凋落物碳氮比越低,越易分解,会向土壤输送大量有机碳,且分解过程中温湿度增加会进一步促进微生物的生长[49]。树木通过凋落物和根部形成周围环境,从而产生小规模的异质性,并可能控制土壤生物的群落模式。
土壤酶是土壤微生物作用于土壤环境的媒介,它是由微生物活动、植物根系分解以及动植物残体腐解的过程中释放到土壤中具有催化作用的生物活性物质。它是最活跃的土壤有机组分之一,反映了土壤中生物化学过程的方向及强度,对土壤生态系统有机碳的固定具有重要作用[50]。Hu等[51]研究发现与单一叶凋落物相比,杉木[Chinesefir-Cunninghamialamcealata(Lamb.) Hook]、枫香(LiquidambaformosanaHance)及桤木(AlnuscremastogyneBurk)混合凋落物处理更利于土壤酶活性(脲酶、转化酶、脱氢酶活性)升高;凋落物分解过程中释放的DOC促进了胞外酶的活性[52]。Ge等[53]发现马尾松(Pinusmassoniana)人工林凋落物剩余质量与酶活性之间呈显著的线性关系,突出了酶活性在影响凋落物分解过程中的重要作用,这将进一步影响森林生态系统中的养分循环。总的来说,凋落物添加之后酶的活性会上调,但上调的程度取决于凋落物类型[54]。但部分树种凋落物中含多酚类或单宁等次生代谢物质,可能会使土壤酶的结构在一定程度上遭到破坏,抑制土壤微生物的活动,进而抑制土壤酶的产生,严重时还会导致部分土壤酶失活。
3.4 植物-土壤-微生物相互作用过程对有机碳稳定性的影响
研究表明,多种因素可以影响碳封存和有机碳稳定性,包括植物(植被覆盖、凋落物、根系分泌物、细根)、土壤(土壤类型、土层深度以及矿物学)以及微生物(群落组成、功能基因)等[12,15],但植物-土壤-微生物相互作用过程对有机碳稳定性的影响还有待进一步研究。
不同植被恢复年限和不同土层深度有机碳的稳定性存在一定的差异。在早期恢复阶段,植被组成和多样性相对简单,群落结构不稳定,地上植被进入到土壤中的SOC相对较少,土壤有机碳分解和转化活动较弱,土壤有机碳相对稳定。随着恢复年限的增加,植被组成结构变的多样化和稳定,凋落物量、细根和根系分泌物显著增加,更多的有机物质进入到土壤中,土壤微生物活动增加,加速了土壤活性有机碳的分解和转化,降低了土壤有机碳的稳定性[55]。此外,植物碳的输入主要决定了土壤碳在表层土壤中的长期持久性,而矿物保护则在底层土壤中占主导地位[56]。深层土壤有机碳主要来源于根部残留物和分泌物,随着土层深度的增加,各恢复类型土壤有机碳和活性有机碳含量逐渐降低,土壤渗透性差、微生物分解活性和根系吸收减少,有机质的输入受到限制,且分解者难以接近深层土壤有机碳,因此,有机碳稳定性增加[57]。在演替的过程中,演替早期微生物群落的生长主要受土壤养分的限制,一般来说,演替早期微生物碳分解基因丰度较高,与氮分解相关的基因丰度增加,导致凋落物的快速分解和土壤养分的增加。随后,主要的资源限制从养分限制转换为另一种环境因子的限制。因此,在演替后期,充足的土壤养分有效性可能导致微生物碳分解基因丰度的下降,导致演替后期凋落物的分解速率的降低[33]。微生物在介导凋落物分解过程中发挥着重要作用,凋落物-微生物-土壤环境三者之间是互相调控的一个动态过程,通过这一系列的反馈与调控,实现生态系统的平衡和稳定(图3)。

图3 凋落物特性-微生物-土壤环境互相调控的动态过程
4 研究展望
凋落物分解对土壤固碳及其稳定性方面的研究已有不少,但土壤作为“黑箱”,土壤有机碳的转移和固定仍存在着很大困难,未来需要生态学、植物学和土壤学工作者的关注。因此,未来亟待强化如下3个方向的研究工作:
加强地上-地下凋落物分解对土壤碳动态影响的协同作用机制研究。目前凋落物分解对土壤碳动态的研究多关注凋落叶的分解,而忽略了地下凋落物对土壤有机碳库的贡献。一般来说,植物根以根凋落物和根沉积碳的形式向土壤提供有机碳,来控制和影响土壤有机碳的动态。根通常与菌根菌丝结合形成土壤团聚体来促进土壤有机碳的稳定。此外,根是一个分级系统,不同直径级别的根的形态、根沉积物的数量和质量、化学计量特征以及菌根真菌性状等方面存在差异,会对分解过程产生影响。植物地上/地下生物量是一个相互联系的有机整体,在植物生长过程中存在着不同的碳分配策略,未来应加强研究地上/地下凋落物分解对土壤碳动态的协同效应。
加强植物凋落物向土壤有机质转化的微生物学机制的研究。在考虑影响凋落物分解的食物网中,不仅要考虑到土壤动物(如蚯蚓,线虫)对凋落物的破碎作用,也要关注土壤微生物对凋落物的腐解作用。土壤微生物具有分解与合成有机质的双重功能,因此,将凋落物分解的土壤微生物学过程与土壤有机质(碳)的稳定性联系在一起,是未来研究的重要课题。此外,目前凋落物分解试验的周期较短,短期的室内分解无法反映真实的分解状态,应加强室内模拟与长期野外分解的连结性。
加强新一代技术手段的开发与应用。凋落物通过影响土壤微生物活性和土壤酶的能力来调控土壤碳循环,但是关于凋落物分解对碳代谢相关功能基因的调控仍然缺乏清晰的认识,未来有望结合代谢组学等技术在该方面进行深入研究。此外,目前缺乏对凋落物-土壤有机质-微生物系统不同来源碳的输入、转化和稳定的定量研究,未来可进一步结合稳定同位素示踪技术,明确凋落物输入后外源碳在土壤中周转和稳定过程之间的关系,认识土壤固碳的本质。
猜你喜欢
煤气与热力(2021年12期)2022-01-19 05:19:30
成都大学学报(自然科学版)(2021年1期)2021-05-22 01:31:24
绿色科技(2020年20期)2020-11-20 01:56:34
中成药(2018年8期)2018-08-29 01:28:26
中成药(2018年2期)2018-05-09 07:20:09
江苏农业科学(2017年6期)2017-05-11 14:35:37
林业勘查设计(2017年1期)2017-03-11 16:49:57
浙江农业学报(2016年7期)2016-06-15 20:30:08
中国农业文摘-农业工程(2016年5期)2016-04-12 05:38:16
现代农业(2016年4期)2016-02-28 18:42:07
