
长江源区水生态系统健康研究进展
2022-03-11 01:59:58熊芳园吴兴华陈宇顺王殿常
中国环境监测 2022年1期
熊芳园,陆 颖,刘 晗,2,程 琳,吴兴华,陈宇顺,2,王殿常
1.中国科学院水生生物研究所,淡水生态与生物技术国家重点实验室,湖北 武汉 430072
2.中国科学院大学,北京 100049
3.中国长江三峡集团有限公司长江生态环境工程研究中心,北京 100038
长江源区位于青藏高原,其生态系统对气候变化十分敏感[1]。 1960—2012 年,该地区升温速率是全球平均升温速率的2 倍[2]。 很少有文章系统总结长江源区水生态系统健康相关研究的现状以及未来的研究重点。 在此背景下,本文对长江源区气候变化、水资源和水生生物相关文献进行了综述,并强调了三者之间潜在的相互作用,以及未来气候变化对该区域水生态系统的影响研究的重要性,以期为长江源区的生态保护提供政策性建议。
1 研究区域
长江源区位于青海省南部,属于青藏高原腹地,地 理 位 置 为 30° 00′ ~ 36° 15′ N、 89°40′~97°50′E(图1)。 三 江 源 区(长 江、黄 河、澜 沧江)的总面积为29. 7 万km2,其中长江源区面积为12. 8 万km2,占43. 2%。 长江源由三大源头组成,主源头为沱沱河,南源头为当曲,北源头为楚玛尔河(图 1)。 主源头沱沱河全长358 km,发源于格拉丹东雪山的姜根迪如冰川。其中,沱沱河与当曲汇合处(囊极巴陇)至巴塘河汇入口的长江上游干流河段为通天河干流段,全长828 km[3]。 北源头楚玛尔河长515 km,流域面积2. 08 万km2,年径流量5. 7 亿m3。南源头当曲长352 km,流域面积3.1 万km2,年径流量11.4 亿m3,是长江三大源流中水流量最大的河流。
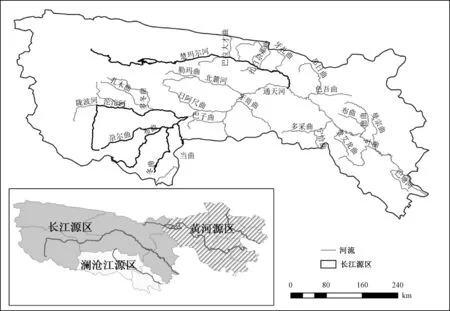
图1 长江源区位置示意图Fig.1 Location of the headwater region of the Yangtze River
长江源区为典型的高原大陆性季风气候,有明显的旱季和雨季。 该区域土地利用类型主要为草地、未利用土地以及水域用地,其中,高寒草甸和高寒草地是草地的主要类型[4]。 同时,三江源地区拥有丰富的河流、湖泊、高山积雪和冰川资源,是重要的产流区和水源涵养区,被称为世界上最大的高山湿地生态系统[4]。
2 研究方法
本综述采用文献计量学方法,于2021 年6 月通过ISI Web of Science 和中国知网(CNKI)数据库进行文献检索。 水生态系统是由水生生物群落与水环境共同构成的具有特定结构和功能的动态平衡系统,因此,本文利用气温、降水、径流、蒸发、湖泊、河流、冰川、浮游生物、硅藻、藻类、鱼类、渔业、底栖生物、大型无脊椎动物、三江源、长江源等关键词,检索数据库中关于水生态研究的有效数据。 根据构建本综述所需信息的相关性,选取截至2021 年6 月的所有长江源区水生态系统相关文献。 通过综合分析相关文献,本文试图回答以下问题:①目前针对长江源区主要开展了哪些水生态相关科学研究? ②长江源区气候变化状况如何? ③长江源区水资源状况如何? ④长江源区水质状况如何? ⑤长江源区水生生物状况如何?⑥长江源区水生态系统健康方面目前主要有哪些研究空白? 未来的研究重点是什么?
3 研究结果
3.1 研究热点
近几十年来,关于长江源区的出版物的数量急剧增加(图2)。 其中,水质方面的研究始于20世纪80 年代[5],气候方面的科学论文自2001 年以来陆续发表[6]。 以长江源区或三江源区为关键词,CNKI 上的文章数量约为2 405 篇,ISI Web of Science 上的文章数量约为932 篇。 其中,近10年发表的占50% 以上,近5 年发表的占25%以上。
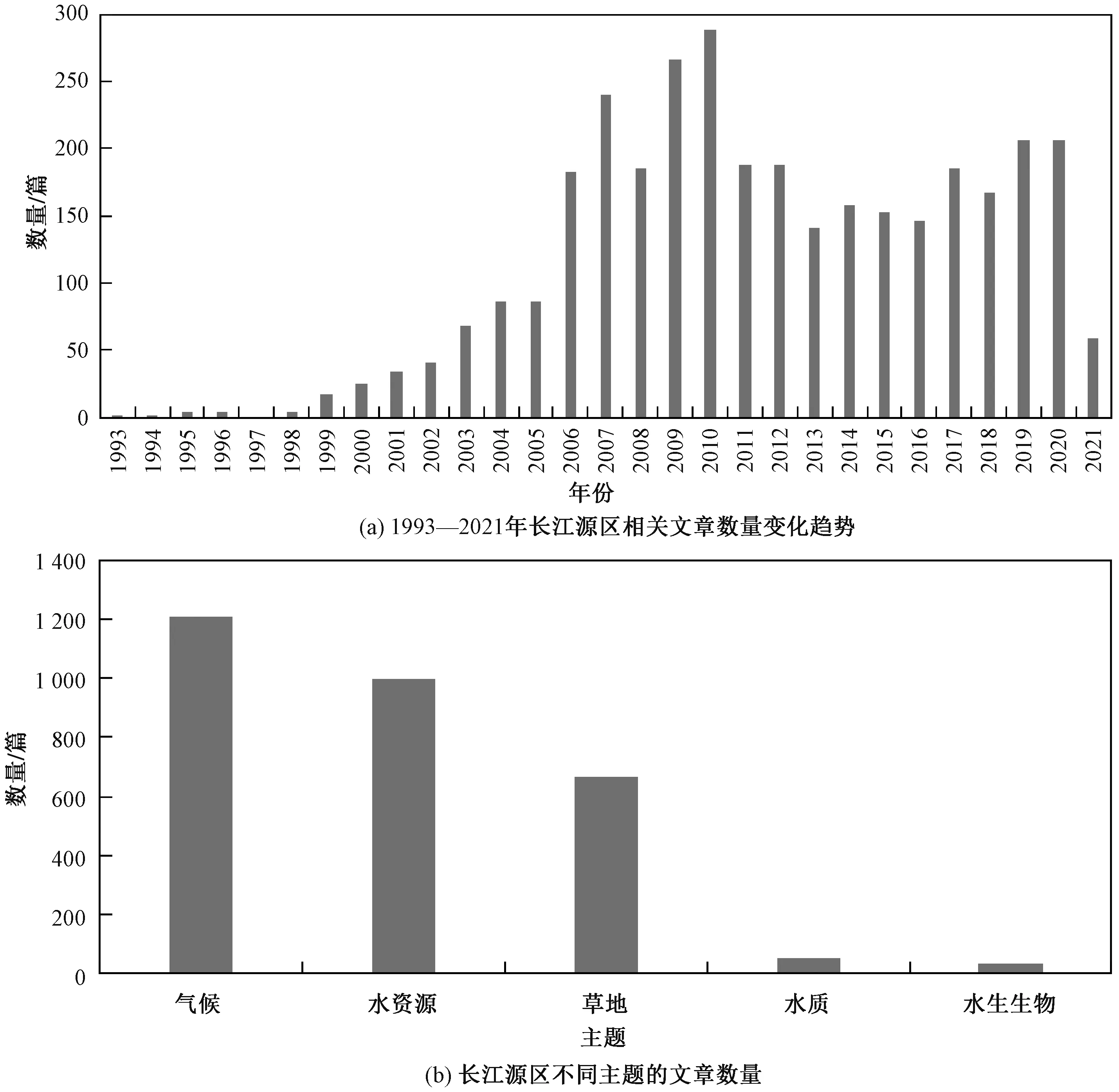
图2 关于长江源区的文章数量Fig.2 Number of papers about the headwater region of the Yangtze River
在这些研究中,研究主题以气候变化为主,其次是水资源变化及草地退化。 近5 年来,越来越多的文章聚焦于冰川融化和水资源变化。 然而,对水质和水生生物(浮游生物、大型无脊椎动物和鱼类等)的研究相对较少。
3.2 气候变化
3.2.1 气温和降水
我国关于三江源区气候变化对水文循环和水资源的影响的研究始于21 世纪初[7-23]。 由于使用了不同时段或不同气象站的观测资料,不同研究对长江源区气温和降水的平均值及变化趋势的研究结果略有差异,但大多数研究都得出了相似的结论[7-24]:1948—2019 年,长江源区年平均气温为-5.2~-0.5 ℃,呈上升趋势,上升速率为0.2~0.5 ℃/10 a[7-24]。 其中,春季平均值为-1.2 ~-0.7 ℃,夏季平均值为3.5~9.7 ℃,秋季平均值为-3.3~-0.6 ℃,冬季平均值为-11.8~-9.5 ℃。升温速率方面,春季为0.1~0.7 ℃/10 a,夏季为0.0~0.8 ℃/10 a,秋季为0.2~0.6 ℃/10 a,冬季为0.0~1.0 ℃/10 a[7-24]。
1948—2019 年,长江源区年平均降水量为310.1~ 2 633.5 mm,夏季降水量最高(223.6~1 268.0 mm),其次为秋季(68.1~657.9 mm)、春季 ( 40.4 ~ 517.5mm ) 和 冬 季 ( 6.1~190.1 mm)[7-24]。 其中,春季和冬季的降水量有增加的趋势,增加速率分别为1.1~26.6 mm/10 a和0.1~9.1 mm/10 a[7-24]。 2007 年之前,夏季和秋季降水量呈降低趋势,降低速率分别为3.1~25.5 mm/10 a 和0.7 ~ 9.6 mm/10 a[7-24]。 2007年之后,夏季和秋季降水量呈增加趋势,增加速率分 别 为 1.5 ~ 26.1 mm/10 a 和 0.4 ~ 43.8 mm/10 a[7-24]。
3.2.2 径流量和蒸发量
以长江源区径流量和蒸发量为研究主题的科学论文的发表始于2001 年[17-29]。 研究显示,长江源区全年平均径流量的增加速率为11.8 ~79.6 m3/(s·10 a),其 中,夏 季 最 大,为25.5 ~161.9 m3/(s · 10 a)。 全 年 平 均 蒸 发 量 为476.0~ 1 350.1 mm,以夏季为最高(474.0 ~493.0 mm),其次为春季(427.0~515.0 mm)、秋季( 270.0 ~ 285.0 mm) 和冬季( 170.0 ~184.0 mm)[17-29]。 大部分研究表明,1956—2014年,长江源区和三江源区的蒸发量呈增加趋势,增速 分 别 为 7.6 ~ 71.6 mm/10 a 和 0.0 ~8.5 mm/10 a。 长江源区蒸发量的增加速率在夏季最高(3.3~24.0 mm/10 a),其次为秋季(1.2~16.7 mm/10 a)、冬季(0.3~14 mm/10 a)和春季(1.4~10.7 mm/10 a)[17-29]。
3.3 水资源
3.3.1 河流
长江源头河流主要发源于冰川雪山。 长江源区河网发达,拥有较多广泛发育的辫状河道,主要河流水系基本成平行的扇形格局,如沱沱河、当曲下游、楚玛尔河、布曲、尕尔曲、通天河上游等,共同组成了庞大的辫状河群[30]。 长江源区共有一级支流340 条,其中集水面积在500 km2以上的河流有85 条[31]。 1960—2016 年,长江源区地表水资源总量呈增长趋势。 1994 年之前,地表水资源总量变化相对较平稳,年际波动较小;1994—2016 年则呈现出较明显的上升趋势。 地表水资源的空间分布较为不均,分布格局为南部和东部较为丰沛,西北部则相对较少[31]。
三江源地区因其重要的淡水供应功能而被称为中国的“水塔”,其中长江源区提供了约20%的长江水量[32]。 根据青海省生态环境状况公报,2020 年三江源区水资源总量为607.6 亿m3,比1998—2019 年的多年平均值增加了 41.3%。2015 年,长江源区地表水资源量为155.2 亿m3,占三江源区地表水资源总量的42.8%;三江源区河流水资源面积约为400 km2,大概占三江源区水资源总面积的2.5%。
3.3.2 湖泊
全球变暖和区域气候变化对湖泊面积的扩大起着重要作用。 此外,湖泊扩张还取决于其他因素,如地质条件、地理位置、地下水补给过程、湖盆形态和人为活动等[33]。 2015 年7—8 月,三江源区湖泊面积约为5 396.5 km2,长江源区湖泊面积约为3 854.7 km2,长江源区湖泊面积占三江源区湖泊面积的70%以上[34]。 1975—2015 年,三江源区和长江源区湖泊面积均有所扩张,扩张面积为2.7 ~ 831.6 km2, 扩 张 速 度 为 0.3 ~ 96.2 km2/a[33-52]。 长江源区湖泊面积扩张的主要原因包括[33-52]:①气候变暖导致冰川加速消融,冰川融水扩大了湖泊面积,如乌兰乌拉湖。 ②降水增加导致湖泊扩张,如卓乃湖。 ③部分湖泊因水量增加而引发决口,其湖水流入邻近湖泊,导致邻近湖泊面积扩张,如库赛湖和海定诺尔湖。 也有部分湖泊因蒸发量大于当地降水量和径流量而发生面积萎缩,如藏南的羊卓雍错等湖泊以大气降水和地表径流为主要补给源,该区降水量虽有所增加,但升温引起的湖泊蒸发效应超过降水量的增加,使该区湖泊面积有所下降。
3.3.3 冰川
长江源区共有冰川627 条,总面积为1 168.2 km2,分别占长江流域冰川总量和总面积的47.1%和61.6%。 长江源区的冰量为983.0 亿km3,相当于840.0 亿km3的水资源[3]。 唐古拉山是长江源区最大的山脉,拥有最丰富的冰川。其中,唐古拉山北部地区共有冰川520 条,总面积为1 021.0 km2,分别占唐古拉山冰川总数和总面积的82.9%和87.4%[3]。
研究显示[39-52]:1969—2005 年,青藏高原冰川面积减少了4 152. 3 km2(103. 8 km2/a);1969—2002 年,长江源区冰川面积减少了68. 1 km2(2. 0 km2/a);1969—2015 年,格拉丹东冰川面积减少了14.9~79.0 km2(0.5~10.0 km2/a)。青藏高原冰川退缩的原因主要包括[39-52]:①气温上升。 气温上升是冰川消退的最关键原因,特别是夏季气温的加速上升。 ②降水与气温的相互作用。 在强降水和低气温条件下,积雪可以持续存在很长时间,反之则会很快消融。 ③高风速。 大风不仅会加速空气对流和水分蒸发,而且会增加冰川的沙尘覆盖,增大冰川对太阳辐射的吸收,加快冰川消融。 ④其他变量也会影响冰川消退,如下垫面条件和阳光强度。
3.4 水质
长江源区大部分河段水质为Ⅱ类及以上[53-61]。 根据目前的研究,1986—2015 年,该区域水质没有明显变化,总氮浓度范围为0.4~0.9 mg/L,总磷为0.01~0.4 mg/L,硝态氮为0.5~2.1 mg/L,氨态氮为0.2 ~0.4 mg/L,化学需氧量为0.6~16.0 mg/L,叶绿素a 低于检测下限,溶解氧为4.8~ 8.6 mg/L,水体pH 呈弱碱性(7.5~8.6)(图3)[53-61]。 与其他水体相比,长江源区水体该8 项指标的含量/数值相对较低。 各类营养盐含量、化学需氧量以及叶绿素a 含量较低,可能是由于长江源区水体尚未受到人类污染,目前处于清洁状态;水体呈弱碱性,可能与该地区碱性的土壤环境有关;溶解氧浓度相对偏低,则与高海拔引起的低气压环境有关[53-61]。
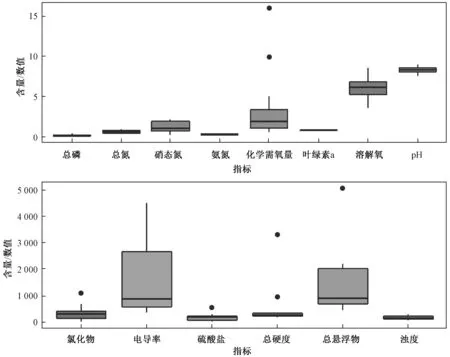
图3 长江源区的水质状况Fig.3 Water quality condition in the source region of the Yangtze River
此外,氯化物的浓度范围为14.3~1 102.5 mg/L,电导率为362.4~4 523.1 μS/cm,硫酸盐为9.8~558.7 mg/L,总硬度为181.0~ 3 307.0 mg/L,总悬浮物为457.0~5 063.0 mg/L,浊度为57.7~286.2 NTU(图3)[53-61]。 总硬度的变化范围较大,反映了该地区复杂的地质组成[53-61]。 与其他水体相比,长江源区水体中该6 项指标的含量/数值相对较高[53-61]。 氯化物含量较高,可能与长江源区部分河水的水化学类型为Cl-Na 型有关;电导率、硫酸盐含量、总硬度较高,可能与长江源区的地质条件有关;总悬浮物浓度、浊度较高,可能是由于长江源区水体泥沙含量较大,整体较为浑浊[53-61]。
金属元素中, 铁的浓度范围为 0.04 ~5.1 mg/L,钾为2.1 ~ 17.6 mg/L,钠为22.6 ~1 017.1 mg/L,钙 为 51.8 ~ 104.6 mg/L, 镁 为7.6~64.7 mg/L,镉为0.004 ~ 0.8 μg/L,铅为0.4~ 23.1 μg/L,砷 为0.8 ~ 66.5 μg/L, 锰 为0.2~240.0 μg/L[53-61]。 长江源区水体中的金属类元素主要有以下几种来源:①水和岩石的相互作用,如硫酸钠盐(Na2SO4)和火山岩[61]。 ②降水。 对于一些总溶解固体含量较低(<40 mg/L)的小支流(流域面积<200 km2),降水可以提供高达24%的水溶性阳离子[61]。
3.5 水生生物
3.5.1 浮游生物
长江源区水体中共有浮游植物6 门134种[62-66]。 其中:硅藻门的种类数最多,有100 种,常见的种类有颗粒直链藻(Melosiragranulata)、尖针杆藻(Synedraacusvar) 和普通等片藻(Diatomavulgare)等;其次是绿藻门,有18 种,常见 种 类 有 弓 形 藻(Schroederiasp.)、 小 球 藻(Chlorellasp.)、并联藻(Quadrigulachodatii)等;再次是蓝藻门, 有 13 种, 常见种类有席藻(Phormidiumsp.)、平裂藻(Merismopediasp.)和小颤藻(Oscillatoriatenuis)等[62-66]。 浮游植物丰度很低,仅为9.1×104~68.0×104ind. /L[62-66]。
长江源区水体中共有浮游动物4 门36种[65-68]。 其中:轮虫的丰富度最高,为16 种;其次是原生动物和桡足类, 分别为10 种和9种[65-68]。 常见浮游动物种类有砂壳虫(Difflugiasp.)、 表 壳 虫(Arcellasp.)、 螺 形 龟 甲 轮 虫(Karutellacochlearis) 和窄腹剑水蚤(Limnoithra sinensis)等。 浮游动物平均密度为489 ind. /L,生物量为0.2 mg/L[65]。
3.5.2 底栖硅藻
长江源区水体中共有底栖硅藻9 属20种[65]。 其中,曲壳藻属(Achnanthessp.)6 种,等片藻属(Diatomasp.)4 种,双眉藻属(Amphorasp.)、桥弯藻属(Cymbellasp.) 和异极藻属(Gomphonemasp.)各2 种,双壁藻属(Diploneissp.)、 桅 杆 藻 属(Fragilariasp.)、 舟 形 藻 属(Naviculasp.) 和菱形藻属(Nitzschiasp.) 各1种。 常见的种类有极小曲壳藻(Achnanthes minutissima)、微细异极藻极细变种(Gomphonema parvulumvar.exilissimum)、 念 珠 状 等 片 藻(Diatomamoniliformis)、隐头舟形藻(NavicuIa cryptocephala)和谷皮菱形藻(Nitzschiapalea)等。底栖硅藻平均细胞密度为0.8×1010ind. /m2。
3.5.3 大型无脊椎动物
长江源区水体中共有大型无脊椎动物32 种,隶属于4 门9 目[65,68]。 常见的种类有霍甫水丝蚓 (Limnodrilushoffmeisteri)、 深 栖 水 丝 蚓(Limnodrilushoffmeisteri) 和 钩 虾(Gammarideasp.)等。 大型无脊椎动物的密度和生物量(干重)分别为59 ind. /m2、0.03 g/m2[65,68]。
3.5.4 鱼类
长江源区共有鱼类21 种,隶属于3 目8属[69-72]。 其中:川陕哲罗鲑(Huchobleekeri)被列为濒危物种和国家一级保护鱼类[69];长丝裂腹鱼(Schizothorax
dolichonema)、 齐 口 裂 腹 鱼(Schizothoraxprenanti)和黄石爬鱼兆(Euchiloglanis kishinouyei)为省级保护物种;川陕哲罗鲑、长丝裂腹 鱼、 黄 石 爬 鱼兆、 裸 腹 叶 须 鱼(Ptychobarbus kaznakovi)和中华鱼兆(Pareuchiloglanissinensis)被列入《中国物种红色名录》[69]。 长江源区鱼类组成简单, 以适应高原环境的裂腹鱼亚科(Schizothoracinae)和条鳅亚科(Noemacheilinae)为优势种,且特有性较高[71]。
4 未来研究方向
总体而言,长江源区水生态系统健康状况良好。 近年来其气象因子以及水资源状况有所改变,未来气候变化可能会进一步影响长江源区水生态系统健康状况。 因此,今后亟须加强对长江源区的本底调查,完善基础数据,关注气候变化对水资源和水质的影响,并探索气候变化对水生生物的影响。
4.1 加强本底调查,完善基础数据
目前已有的长江源区水生态系统研究大多集中在河流,而忽视了对湖泊和水生生物的研究。此外,大多数的研究报道仅基于一次监测数据,且大部分调查的采样力度(采样点、采样断面、采样周期等)不够,使得整个流域的水生生物数据不够全面,甚至不够准确。 目前关于长江源区水生态系统水生生物状况的研究非常少。 同时,很多文献只列出了物种名录,而未报道各类生物的丰度、生物量等重要指标。 这种粗糙的调查和报道方式大大降低了基础数据的使用价值,也给今后的现状与历史数据分析带来困难。
为此,提出了以下几点建议:①开展持续性、全区域、多指标的本底调查。 加强对长江源区干流、支流、湖泊和湿地等不同区域和生境的水生态系统调查,探索流域内水生生物多样性保护对策,阐明水生生物赖以生存的关键栖息地的保护与修复策略。 ②构建标准化的科学监测体系。 整合并构建长江源区水生态监测标准方法体系及监测网络体系,为长江源区水生态安全评估与保障提供基础理论和数据支撑。 同时,通过搭建大数据平台、树立伙伴型部门间关系、创建部门间信任合作机制等举措,建立基于大数据技术的政府信息资源跨部门共享机制。 ③加大宣传力度。 地方政府应大力宣传环境保护和生态文明理念,同时支持研究人员的监测工作,特别是对鱼类的采集。
4.2 关注气候变化对水资源和水质的影响
由于长江源区受人类活动干扰少,气候变化成为其水生态系统面临的主要挑战。 气候变化导致的水文变化,特别是积雪融化量的增加,提升了长江源区源头河流及相关湖泊的水位。 浅水湖泊具有较高的面积体积比,更容易受到风力的影响。风力增强会加大沉积物与水的相互作用,改变浊度、养分循环和溶解氧浓度[73]。 此外,温度升高和降水增多则会导致径流增大,使得一些内陆水域的水体盐度和硬度降低[74]。 水体中的磷酸盐、硝酸盐、总磷浓度也与气候变化(如降水、气温)之间具有相关关系[75]。
虽然目前对长江源区气候变化与水质、水资源量之间的相互关系的研究还处于起步阶段,但随着研究的进一步深入,定量研究两者之间的相互关系成为未来发展的必然趋势。 确定并量化两者之间的相互关系,有助于加强对气候变化趋势和水质、水资源量变化趋势的预测。 因此,未来需进一步加强长期监测,同时结合定量分析、水文/水质模型及气候情景分析,从而更好地预测气候变化下的水质、水文、水资源及酸性沉积变化。
4.3 探索气候变化对水生生物的影响
浮游植物可以作为“哨兵”来指示当前气候变化对生态系统的影响。 前期的相关研究报道了长江源区部分点位的浮游生物种类、丰度和生物量情况,但未涉及浮游生物与气候变化之间的相互关系。 相比之下,在其他研究区域,大量研究论文对全球变暖下的浮游生物动力学特征进行了分析。 一些研究表明,湖泊水温的升高将导致浮游植物生产力下降[76]。 例如,由于气候变暖,卡里巴湖(Lake Kariba)的初级生产力下降了50%[77]。然而,另外一些湖泊的初级生产力则会随气候变暖而升高。 例如,由于气候变暖,在20 世纪60—90 年代,维多利亚湖(Lake Victoria)的总氮含量增加了两倍,初级生产力也增加了两倍[77]。 随着气候变暖导致浮游植物生物量增加,能够固氮且不受氮限制的蓝藻将取代硅藻成为优势种[78]。
在大型无脊椎动物群落方面,只有一篇文章对长江源区的物种名单、丰度和生物量进行了分析。 然而,目前的研究表明,气候变暖会影响到大型无脊椎动物的群落结构,极端水文事件(如干旱、洪水)可能会显著改变大型无脊椎动物的群落组成,影响其物种丰度[79]。 气候变暖对大型无脊椎动物群落的影响主要包括:①改变大型无脊椎动物的群落结构。 例如,由于富营养化和气候变暖引起缺氧加剧,摇蚊(Chironomussp.)和幽蚊(Chaoborussp.)将成为优势蚊种[80]。 ②改变大型无脊椎动物的生物量。 例如,由于气候变暖,维多利亚湖、汉宁顿湾(Hannington Bay)、菲尔丁湾(Fielding Bay)和默奇森湾(Murchison Bay)等地的大型无脊椎动物的生物量显著减少[80]。
目前仍然缺乏关于气候变化对长江源区鱼类生活史及种群结构的影响的研究。 气候变化,尤其是温度、风速和降水量的变化,会引起鱼类群落组成、鱼类生产力、入侵性水生植物扩散方式以及寄生虫-媒介-宿主相互作用方式的改变[81]。 近几十年来,温度和降水变化显著增加了水体的净初级生产力,而初级生产力的增加可能会加快以其为食的高级消费者(鱼类)的增长速率[82]。 此外,水温可以决定鱼类的分布范围。 例如,随着水温的升高,大多数鲤科鱼类的栖息地可能会向北扩张[83]。 在雅鲁藏布江,鱼类的生长与年平均气温呈显著负相关关系,如杂食性鱼类拉萨裸裂尻鱼(Schizopygopsisyounghusbandi)和肉食性鱼类拉萨河尖裸鲤(Oxygymnocyprisstewartii)[1]。 同时,温度和降水的增加会提高鱼类肌肉组织中的汞的富集速率[84]。 由于气候变暖,一些冷水性鱼类甚至可能会在未来发生灭绝。
总之,气候变化不仅能够直接影响某一营养层级水生生物的群落结构,而且能够通过上行效应和下行效应间接影响其他营养层级水生生物的物种组成及资源量大小等。 理清水生生物对气候变化的响应过程,探究气候变化下的水生生物群落构建和维持机制及关键性驱动因子,成为在长江源区开展水生生物资源保护的关键性前提,也是淡水生态学研究的关注重点。 因此,需要进一步加强气候变化对具有不同生境适宜性以及处于不同生活史阶段的特定水生生物物种的影响研究。 此外,通过性状分析,还可以为厘清不同生物类群、不同地理区域和不同生态系统类型之间的性状-脆弱性关系提供足够的信息。 对于具有不同气候变化适应能力的鱼类来说,单一的管理策略可能并不理想,生态环境管理者还需要确定针对具有不同适应能力的多种鱼类的多重管理机制,以便提高所有鱼类在气候变化压力下的适应和恢复能力[81]。
猜你喜欢
包装工程(2023年16期)2023-08-25 11:32:34
北京大学学报(自然科学版)(2022年2期)2022-04-08 12:30:28
现代艺术(2022年1期)2022-02-07 11:06:24
小天使·四年级语数英综合(2020年2期)2020-02-04 16:12:41
创新作文(1-2年级)(2019年2期)2019-08-06 04:19:18
金色少年(奇趣科普)(2017年12期)2017-12-26 06:20:00
环境科技(2016年4期)2016-11-08 12:18:58
长江科学院院报(2016年3期)2016-03-23 12:04:13
长江蔬菜(2014年1期)2014-03-11 15:09:48
椰城(2013年4期)2013-11-16 06:31:11
